
甲虫表皮碳氢化合物研究进展
2022-03-24 10:01薛怀君
环境昆虫学报 2022年1期
王 苑,薛怀君
(1. 安徽大学物质科学与信息技术研究院,合肥 230601;2. 中国科学院动物研究所,北京 100101;3.南开大学生命科学学院,天津 300071)
昆虫表皮碳氢化合物(cuticular hydrocarbons, CHCs)是正烷烃、不饱和烃以及甲基支链烃的混合物,碳链长度一般在21到50个碳原子之间,有时甚至可达70个(Blomquist and Bagnères, 2010)。除了能够在昆虫表皮形成疏水层防止水分丧失和抵抗有害物质及病菌入侵之外(Gibbs, 1998; Herzner and Strohm, 2007),昆虫表皮碳氢化合物还具有多种生物功能。研究表明,表皮碳氢化合物被昆虫作为多种化学识别信号,其中包括种内性别识别、种间生殖隔离和种间信息素通讯等。社会性昆虫的表皮碳氢化合物则承载了更多的功能,包括族群识别、巢间识别、品级分化等(Howard and Blomquist, 2005; Blomquist and Bagnères, 2010; Chung and Carroll, 2015)。
鞘翅目Coleoptera昆虫俗称甲虫,是世界上多样性最丰富的昆虫类群,已记述的种类超过38万种(Zhangetal., 2018)。目前对甲虫表皮碳氢化合物研究开展颇多,本文主要对表皮碳氢化合物在甲虫化学通讯中的作用进行了归纳与总结,并对甲虫表皮碳氢化合物组成的影响因素、生物合成的分子机制及在化学分类中的应用等几个方面的研究进展进行了综述,以期为相关研究的进一步开展提供参考。
1 甲虫表皮碳氢化合物在化学通讯中的作用
1.1 性信息素
1.1.1利用表皮碳氢化合物作为性信息素的甲虫类群
性信息素在昆虫化学通讯中提供了重要的信号,一般来说,挥发性性信息素吸引远处的异性,而接触性性信息素则有助于交配识别和激发交配行为(Wyatt, 2003)。甲虫的表皮碳氢化合物主要为非挥发性的,据统计,表皮碳氢化合物至少在甲虫的9个科中的一些种类中起到性信息素的作用(表1)。
隐翅虫Aleocharacurtula表面用薄层树脂包裹或索氏抽提后的雌虫对雄虫不再具有性吸引作用,而雌虫身体的多个部位均能激发雄虫的交配行为,并且正己烷的雌虫体表提取物可以诱发雄虫的交配行为,这是在隐翅虫中第一次发现接触性性信息素的存在(Peschke, 1978),在后续的研究中确认了其成分为多种碳氢化合物(Peschke and Metzler, 1987; Peschke, 1987a)。
Ginzel等将脊虎天牛Xylotrechuscolonus雌性的表皮碳氢化合物用有机溶剂漂洗,发现其失去了对雄性的吸引力,将表皮碳氢化合物重置于漂洗过的雌性个体后则恢复了对雄虫的吸引,证明了雌虫表皮碳氢化合物在交配行为中的作用(Ginzeletal., 2003a)。对光肩星天牛Anoplophoraglabripennis的表皮碳氢化合物进行研究,以离心管作为假试体,发现用表皮碳氢化合物中的5种成分按照一定比例混合后处理,可以激发雄虫的交配行为,证实了雌性表皮碳氢化合物起到性信息素的作用(Zhangetal., 2003)。与之类似,Ibeas等对墨天牛Monochamusgalloprovincialis的交配行为进行了研究,当玻璃棒用雌性正己烷提取物处理后,雄性天牛会表现出与玻璃棒交配的行为,从而证实雌性表皮碳氢化合物起到性信息素的作用(Ibeasetal., 2009)。
另外,研究表明,表皮碳氢化合物在吉丁科(Lelitoetal., 2009; Silketal., 2009)、萤科(Ming and Lewis, 2010)、埋葬甲科(Steigeretal., 2007, 2009)、金龟科(Ortiz-Domínguezetal., 2006)、瓢甲科(Hemptinneetal., 1998)、叶甲科(Petersonetal., 2007; Kawazuetal., 2011; Zhangetal., 2014; Xueetal., 2016a; Shimomuraetal., 2017)和象甲科(Mutisetal., 2009; Chenetal., 2019; Isaetal., 2019)的一些种类起到性信息素的作用(表1)。
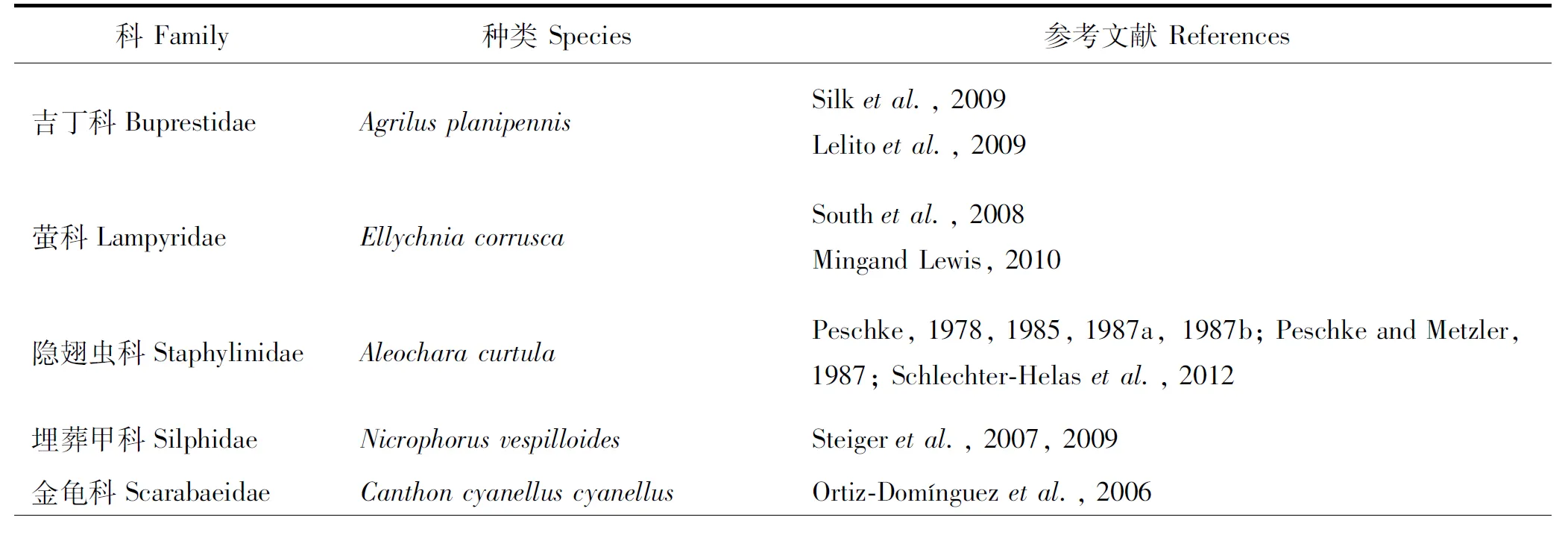
表1 利用表皮碳氢化合物作为性信息素的甲虫类群
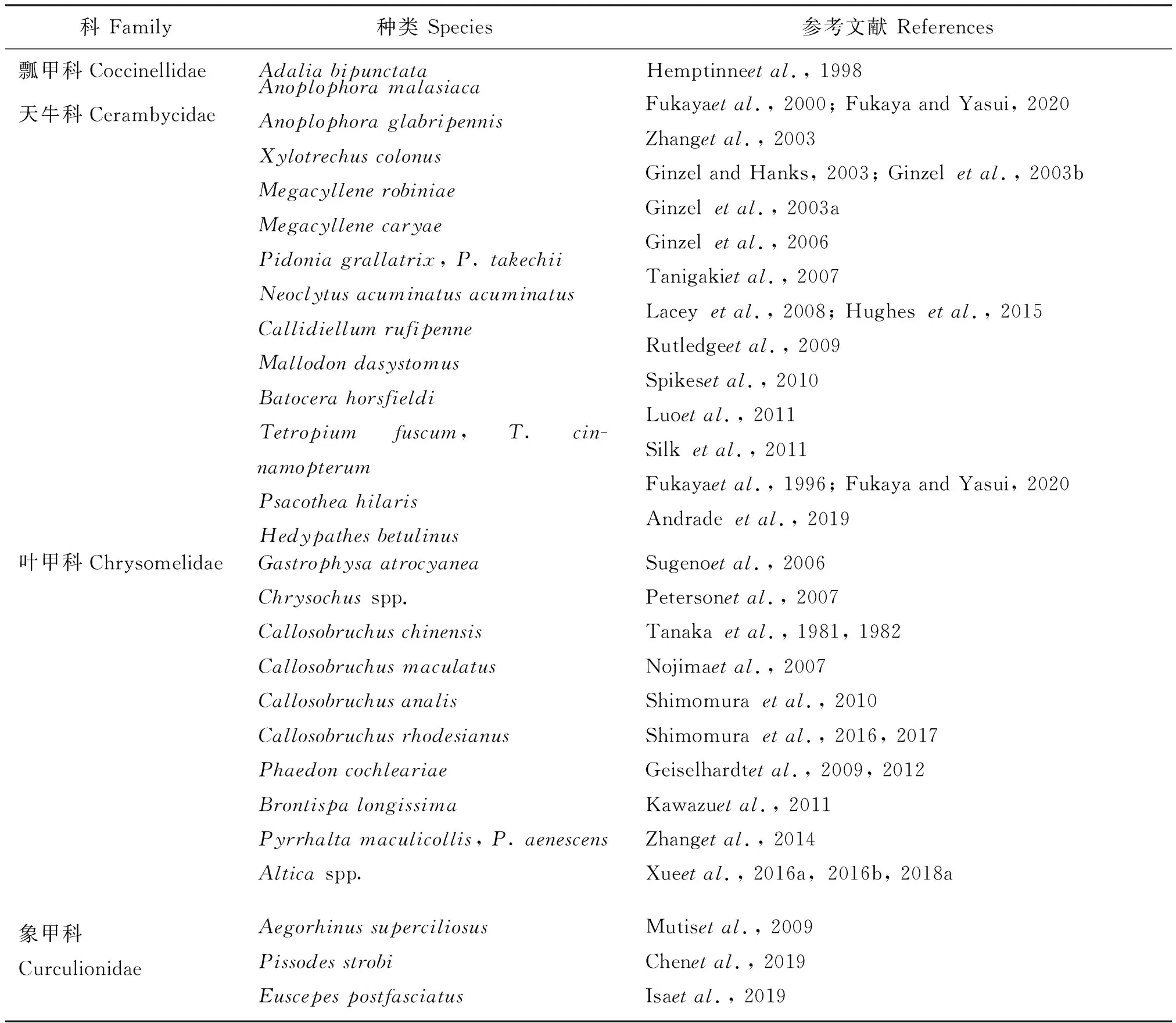
续表1 Continued table 1
1.1.2种内个体的识别
种内个体的识别在昆虫中非常重要,两性个体可以借此获益,比如,获得更有竞争力的后代和避免无谓的投资(Andersson and Simmons, 2006; Scharfetal., 2013)。
隐翅虫种群中存在偏雄现象,雄性之间竞争激烈。性未成熟、饥饿和已经多次交配的弱势雄虫存在种内的化学拟态,为了避免遭到同性攻击会产生类似于雌虫的性信息素,即两种烯烃((Z)-7-Heneicosene和(Z)-7-tricosene)(Peschke, 1985, 1987a,1987b)。在这种情况下,雌虫可以通过表皮碳氢化合物来识别雄性个体,拒绝与具有雌性化学表型的雄虫进行交配(Peschke, 1987a,1987b)。A.curtula雄虫可以通过表皮碳氢化合物的信息识别雌虫的交配状态,从而可以减少雄虫无谓的时间和能量浪费(Schlechter-Helasetal., 2012)。
埋葬甲Nicrophorusvespilloides对动物尸体有领地意识,雌虫对雄性个体有较高的容忍度,当被测试的雌雄个体表皮碳氢化合物互换之后,雌性的攻击行为发生了逆转,说明雌性埋葬甲通过表皮碳氢化合物来识别同种个体的雌雄;N.vespilloides雌虫也可通过表皮碳氢化合物信号识别亲本个体,表现出明显较弱的攻击性(Steigeretal., 2009)。
对3种跳甲(蛇莓跳甲Alticafragaria、老鹳草跳甲A.viridicyanea和蓟跳甲A.cirsicola)的研究表明,跳甲的雄性个体在交配时均可以识别被选择对象的性别,当互换被选择对象的表皮碳氢化合物时,交配选择行为发生了明显的改变;雄性个体也可以很好地识别初羽化和性成熟的雌性个体,当互换被选择对象的表皮碳氢化合物后,雄虫的交配选择行为也会发生改变。据此推测,表皮碳氢化合物是跳甲种内个体识别的重要信号(Xueetal., 2016b)。
但是,并不是所有甲虫的种类都具有种内个体识别能力,个体间表皮碳氢化合物指纹图谱的差异也并不意味着一定会被雄虫当作交配识别的信号。比如,在猿叶甲Phaedoncochleariae中,虽然表皮碳氢化合物起到性信息素的作用,但雌雄之间仅存在相对含量的差别,雄虫并不能依靠这种差异来识别种内个体的性别差异(Geiselhardtetal., 2009);异色瓢虫Harmoniaaxyridis雌虫交配后和交配前表皮碳氢化合物一些成分的相对含量发生变化,但是异色瓢虫雄虫在选择条件下并不能识别不同交配状态的雌虫(Legrandetal., 2019)。
1.1.3在生殖隔离中的作用
种间特异性的表皮碳氢化合物可以起到性信息素的作用,在同域物种的生殖隔离中常常发挥重要作用。对两种同域分布的访花天牛(Pidoniagrallatrix和P.takechii)的研究表明,雌虫的表皮碳氢化合物会引发同种雄性的交配行为,两个种的表皮碳氢化合物存在种特异性,通过替换雌性表皮碳氢化合物的方法证实了雄性可以通过这种接触性信息素识别同种雌性和异种雌性(Tanigakietal., 2007)。通过替换体表碳氢化合物的方法对两种萤叶甲(榆黄毛萤叶甲Pyrrhaltamaculicollis和榆绿毛萤叶甲P.aenescens)(Zhangetal., 2014)和两种跳甲(蛇莓跳甲和老鹳草跳甲)(Xueetal., 2016a)的研究也证实了表皮碳氢化合物在近缘物种间生殖隔离的作用。
Otte等对两种同域分布的猿叶甲(Phaedoncochleariae和P.armoraciae)进行了交配行为研究和表皮碳氢化合物表型比较,发现两个种在取食相同的寄主植物情况不存在行为隔离(性隔离),而在取食不同植物时雄虫则可以表现出明显的交配选择倾向,寄主植物诱导表皮碳氢化合物表型的可塑性导致了近缘种的行为隔离(Otteetal., 2016)。
表皮碳氢化合物在杂交体系中仍然会一定程度上起到性信息素的作用。蛇莓跳甲和老鹳草跳甲的各杂交后代(杂交一代、二代和回交代)均具有各不相同,也不同于亲本的表皮碳氢化合物指纹图谱(profile)。其中,蛇莓跳甲和杂交F1的雄虫可以利用表皮碳氢化合物的差异识别自身和老鹳草跳甲的雌虫,但杂交F1并不能有效的识别老鹳草跳甲和杂交F1雌虫,这种识别不对称的现象也出现在其他一些组合。总的来看,表皮碳氢化合物的这种差异并不能有效地阻止杂交个体和亲本个体之间的基因交流(Xueetal., 2018a)。
1.1.4性信息素活性成分的鉴定
在鞘翅目昆虫中,多数昆虫的表皮碳氢化合物成分非常复杂,因此活性成分的确认和鉴定存在很大难度,因此,目前只有为数不多的甲虫种类的表皮碳氢化合物活性成分得到了鉴定(表2)。

表2 部分甲虫的性信息素活性成分
隐翅虫Aleocharacurtula雌虫表皮碳氢化合物成分复杂,包括了直链烷烃、直链烯烃、单甲基烷烃和双甲基烷烃等,碳链长度在21到29,其中两种烯烃((Z)7-Heneicosene和(Z)7-tricosene)是接触性性信息素的主要活性成分(Peschke and Metzler, 1987)。
性信息素活性成分的鉴定工作主要集中于经济重要的种类,比如天牛科昆虫。目前已有超过100种天牛的挥发性性信息素被鉴定(Fukaya and Yasui, 2020),但由于接触性性信息素在害虫防治中的潜在利用价值相对较低(Nojimaetal., 2007),仅在少数种类中得到了研究。不同种类的天牛接触性性信息素的复杂程度有很大差别,比如,7-甲基二十七烷(7Me-C27)是黑腹尼虎天牛Neoclytusacuminatusacuminatus接触性信息素的主要成分,7Me-C25和9Me-C27可起到增效作用(Laceyetal., 2008);与之类似,黄星天牛Psacotheahilaris的接触性性信息素组成也比较简单,只包含一种碳氢化合物Z8-21Me-C35(Fukaya and Yasui, 2020);光肩星天牛Anoplophoraglabripennis的接触性性信息素则相对复杂,由5种烯烃((Z)-9-tricosene, (Z)-9-pentacosene, (Z)-7-pentacosene, (Z)-9-heptacosene, (Z)-7-heptacosene)共同组成(Zhangetal., 2003);而Anoplophoramalasiaca的接触性性信息素则包含了8种碳氢化合物、5种脂肪酮和3种内酯,这些物质共同作用才能诱发交配行为(Fujiwara-Tsujiietal., 2019; Fukaya and Yasui, 2020),这也是迄今为止所发现的最复杂的昆虫接触性性信息素组成(Fukaya and Yasui, 2020)。
叶甲科个别种类的性信息素成分也有一些研究。豆象属Callosobruchus昆虫为重要的仓储害虫,其中绿豆象C.chinensis的接触性性信息素由几种碳氢化合物和一种二羧酸((E)-3,7-dimethyl-2-octene-1,8-dioic acid)组成(Tanakaetal., 1981, 1982);四纹豆象C.maculatus具有复杂的接触性信息素,其活性成分为2,6-二甲基-1,8-二酸(2,6-dimethyloctane-1,8-dioic acid)、壬二酸(nonanedioic acid)以及C27-C35的直链烷烃和甲基支链烃,这些成分协同作用,引发雄性的交配行为(Nojimaetal., 2007);C.analis的接触性信息素的活性成分为2,6-二甲基-1,8-二酸(2,6-Dimethyloctane-1,8-dioic acid)、一种单帖二羧酸(callosobruchusic acid)和一些碳氢化合物(Shimomuraetal., 2010);而C.rhodesianus的接触性性信息素比较特殊,为两种酮类(hexahydrofarnesyl acetone和2-nonadecanone),C27-C33的直链烷烃和甲基支链烃可起到增效剂的作用(Shimomuraetal., 2016; 2017)。蓼蓝齿胫叶甲Gastrophysaatrocyanea雌性产生的甲基二十七烷和甲基二十九烷(9-methylheptacosane、11-methylheptacosane、9-methylnonacosane和11-methylnonacosane)可以激发雄虫的交配行为(Sugenoetal., 2006)。猿叶甲Phaedoncochleariae体表的非极性成分(碳氢化合物)可以激发雄虫的交配行为,极性成分不能使雄虫产生交配行为,但极性成分和非极性成分混合可以显著提高交配活性(Geiselhardtetal., 2009)。
1.2 聚集信息素
为应对不利环境和食物缺乏的境况,很多昆虫有滞育现象,在这个阶段,常常大量个体聚集在一起,这有助于昆虫提高生存率和寻找交配对象(Sussetetal., 2013)。Susset等人的研究证实烷基甲氧基吡嗪(alkyl-methoxypyrazine)在瓢虫Adaliabipunctata中起到聚集信息素的作用,同时也发现滞育个体的表皮碳氢化合物可以延长瓢虫的停留时间(Sussetetal., 2013)。Durieux等对异色瓢虫Harmoniaaxyridis越冬聚集行为的研究表明,长链的碳氢化合物中一类物质可以引导瓢虫寻找聚集地点,另一类物质促使瓢虫聚集成群(Durieuxetal., 2012);对毛斑长足瓢虫Hippodamiaconvergens的越冬聚集行为的研究发现,瓢虫爬行足迹残余的碳氢化合物对同种个体有吸引作用,二十三烷是其主要活性成分,当碳氢化合物和甲氧基吡嗪(methoxypyrazine)共同使用时可维持瓢虫长时间的聚集(Wheeler and Carde, 2014)。甲虫的很多类群,比如叶甲类昆虫也常常具有群集越冬的现象,但表皮碳氢化合物是否起到聚集激素的作用尚未见到报道。研究还发现,接触性表皮碳氢化合物在异色瓢虫的非越冬聚集行为也起到重要作用(Durieuxetal., 2014)。
1.3 标记信息素
产卵地点选择不当可能会导致食蚜瓢虫后代面临饥饿以及由此引发的同种相残(cannibalism,即动物杀死并取食同种其他个体的行为)和集团内捕食(intraguild predation,即某物种以其竞争者(同营养级)为食的现象)(Meisneretal., 2011),因此产卵地点的选择对于雌虫至关重要(Magroetal., 2007)。
食蚜瓢虫利用幼虫足迹中的碳氢化合物作为识别信号,选择未被占领的区域产卵。这些化学物质是由位于幼虫第十腹节的臀片(anal disk)分泌(Laubertieetal., 2006),其主要成分为烷烃,包括直链、一甲基和二甲基烷烃(Hemptinneetal., 2001; Magroetal., 2007)。尽管同种和异种的幼虫印记都可能阻止雌性产卵,但雌虫往往对前者的反应更为明显,并且这种阻碍作用存在很强的种差异性。例如,对二星瓢虫Adaliabipunctata、十星瓢虫Adaliadecempunctata和七星瓢虫Coccinellaseptempunctata的比较研究发现,同种和异种幼虫的印记均会显著地阻碍二星瓢虫和十星瓢虫产卵,但同种幼虫的印记对七星瓢虫产卵仅有轻微的阻碍作用,而异种幼虫印记对七星瓢虫产卵的阻碍作用不明显。对5种瓢虫的研究发现,同种幼虫、蛹和两性成虫的化学信息素存在均会阻碍雌虫产卵,这种阻碍程度具有显著的密度依赖性和种特异性(Mishraetal., 2012);另一项研究显示,异种卵、幼虫、蛹和两性成虫的化学信息素存在也均会阻碍瓢虫Menochilussexmaculatus雌虫的产卵,且抑制程度具有密度依赖性,活跃阶段比静止阶段的虫态具有更强的产卵阻碍作用(Mishraetal., 2013)。
除了阻碍产卵外,幼虫化学印记对瓢虫的取食也具有阻碍作用。例如,七星瓢虫和异色瓢虫的幼虫均回避在留有同种幼虫化学印记的区域取食;七星瓢虫也回避在留有异色瓢虫幼虫化学印记的区域取食,但异色瓢虫对七星瓢虫的化学印记没有明显反应(Meisneretal., 2011)。另一研究表明,七星瓢虫和C.transversalis的幼虫和雌成虫留下的化学印记(碳氢化合物)会影响到其它幼虫的捕食效率和个体发育(Kumaretal., 2014)。
1.4 化学拟态
虽然化学通讯在很多生物中占主导地位,但由于化学通讯不易被察觉,由化学刺激引起的拟态现象往往受到忽视(Dettner and Liepert, 1994),前期大多数的研究主要集中于视觉拟态(Pasteur, 1982),
许多昆虫寄宿于其它群居昆虫的巢穴中,因为这些群居昆虫群体内部常常拥有复杂的通讯系统,可以识别同巢者并分辨异己,入侵者进化出一系列适应性机制,以躲避宿主识别。许多入侵者获得了模仿其宿主物种化学信号的能力。
嗜蚁的节肢动物种类繁多,目前据估算多达1万至10万种(Parker and Grimaldi, 2014; von Beerenetal., 2018),这里面包括甲虫的很多种类。甲虫为了适应于蚁巢的生活,其中一种常用的策略就是化学拟态。金龟科Myrmecaphodiusexcavaticollis的宿主是Solenopsis和Iridomyrmex两个属的蚂蚁,在寄主蚁巢内,M.excavaticollis可以通过交哺(trophallaxis)的方式直接从工蚁处获得食物,或者以蚂蚁幼虫、死去的蚂蚁以及蚂蚁的战利品等为食,并且可以自由活动而不受工蚁的攻击。M.excavaticollis通过各种和宿主接触的行为获取其宿主的表皮碳氢化合物,以此达到化学拟态的目的,其获得的碳氢化合物在离开宿主两周后会逐渐消失,当接触到新的宿主种类后又会开始新的化学模拟(Vander Meer and Wojcik, 1982)。阎甲Sternocoelishispanus和隐翅虫Chitosanigrita寄居于Aphaenogaster属蚂蚁巢穴内,两种甲虫均具有和寄主相同的特定碳氢化合物指纹图谱,但这些碳氢化合物并非获得性来自宿主,而是甲虫自身的合成(Lenoiretal., 2012)。瓢虫Diomusthoracicus幼虫专性寄生于蚂蚁Wasmanniaauropunctata的巢穴内,取食蚂蚁的幼体,并不会受到工蚁的攻击,化学分析结果显示,多数种类的表皮碳氢化合物为瓢虫幼虫和宿主共享(Vantauxetal., 2010)。隐翅虫科Tetradonia属的种类为多食性,每种隐翅虫可以捕食多种Eciton属蚂蚁,而Ecitomorpha属及Ecitophya属的隐翅虫为专性寄生,每个种专门寄生于Eciton属的一种蚂蚁。比较研究显示,专性寄生者表皮碳氢化合物成分和含量更接近于Eciton属宿主,而兼性寄生的种类表皮碳氢化合物的相似性和宿主相差较远,专性寄生甲虫更多采用了化学拟态的整合策略融入蚁巢(von Beerenetal., 2018)。
大花蚤Metoecusparadoxus寄生于胡蜂Vespulavulgaris的巢穴内,研究发现,M.paradoxus与其宿主V.vulgaris的表皮碳氢化合物组成相似,而与另一种近缘的胡蜂V.germanica则有明显差异,M.paradoxus通过这种化学模拟而不被宿主攻击。研究者推测,M.paradoxus可能是通过对所取食的胡蜂幼虫表皮碳氢化合物回收利用的方式达到化学模拟的目的(van Oystaeyenetal., 2015)。
另一类甲虫寄生于蜚蠊目的白蚁巢穴中,例如,隐翅虫Trichopseniusfrosti和宿主北美散白蚁Reticulitermesflavipes的表皮碳氢化合物在成分组成和相对含量上均非常相似,同位素示踪实验表明,隐翅虫T.frosti可以通过自身合成的方式来模仿宿主的表皮碳氢化合物(Howardetal., 1980)。进一步对另外3种隐翅虫(Philotermeshowardi、Trichopensiusdepressus和Xenistusahexagonalis)及其寄主南方散白蚁Reticulitermesvirginicus的研究发现,这3种隐翅虫与其白蚁寄主的表皮碳氢化合物指纹图谱均非常相似,并且至少其中的一个种(X.hexagonalis)也是通过自体合成的方式实现化学拟态(Howardetal., 1982)。
还有一些甲虫寄生于其它种类的甲虫洞穴之中,例如拟步甲Eremostibesopacus寄居于另一种拟步甲Parastizopusarmaticeps的洞穴之中(Heinetal., 1996),二者之间表皮碳氢化合物组成高度相似,E.opacus通过对宿主进行化学模仿的方式以达到防止宿主攻击的目的(Geiselhardtetal., 2006)。
芫菁Meloefranciscanus进化出非常奇特的“性欺骗”行为。其幼虫为寄生性,在美国加利福尼亚莫哈韦Mojave沙漠地区,寄主为沙丘打洞独居的回条蜂Habropodapallida。M.franciscanus的幼虫孵化后在植物茎秆上聚集成簇,可以模拟雌蜂表皮碳氢化合物成分,从而吸引雄蜂来交配。芫菁幼虫用特化的足抱握住雄蜂而被带入宿主巢穴,在蜂巢能取食蜜蜂储备的食物以及蜂幼体,完成个体发育(Hafernik and Saul-Gershenz, 2000; Saul-Gershenz and Millar, 2006)。进一步研究发现,M.franciscanus在俄勒冈地区的宿主为另一种同属的蜜蜂H.miserabilis,由于两个不同地理种群的宿主(蜜蜂)种类不同,M.franciscanus受到强烈的选择压力,芫菁幼虫进化出了模拟当地的雌蜂的化学信号的能力(Saul-Gershenzetal., 2018)。
1.5 宿主/猎物的定位和识别
对于寄生、共生和捕食者而言,昆虫宿主的表皮碳氢化合物常常被用作定位和识别的信号。专性寄生的肿腿蜂Cephalonomiawaterstoni雌虫可以追踪0.0001当量的扁谷盗Cryptolestesferrugineus幼虫体表化合物,而对非宿主Cryptolestespusillus和Triboliumcastaneum的幼虫体表化合物则无明显趋性。研究发现,自然留存在滤纸上的化学印记激发寄生蜂的追踪行为可维持一周的时间,而人工涂抹在平板玻璃上的化学物质则可以维持大约 2周(Howard and Flinn, 1990)。另一项研究显示,杂拟谷盗Triboliumconfusum爬行过后会留下表皮碳氢化合物印记,这种印记可以存留大约1 d的时间,体外寄生蜂Holepyrissylvanidis利用这种印记来寻找宿主,并最终通过直接接触的方式完成宿主识别(Füerstenauetal., 2017)。螨虫Grandiellarugosita专性共生于龟甲Acromissparsa体表,G.rugosita不仅对龟甲的体表化学物质具有趋性,还可以依赖于龟甲的体表化学物质识别宿主和非宿主(Beranetal., 2014)。甲虫的表皮碳氢化合物还可作为捕食者搜寻猎物的信号,跳甲属Altica种类是捕食性蓝蝽Zicronacaerulea最偏好的猎物,行为实验表明,老鹳草跳甲的表皮碳氢化合物可以激发蓝蝽的捕食行为,是其搜寻和识别猎物的关键信号(Xueetal., 2018b)。
在一些类群,表皮碳氢化合物还可以起到化学防御的作用。例如,二星瓢虫和七星瓢虫卵表面的碳氢化合物均可以阻碍其幼虫取食同种的卵(Hemptinneetal., 2000);异色瓢虫在欧洲属于入侵种,研究发现,异色瓢虫幼虫对欧洲本地物种Calviadecemguttata卵的取食偏好性明显高于Calviaquatuordecimguttata的卵,其原因是二者卵表面的碳氢化合物成分存在差异,前者表皮碳氢化合物的多样性明显低于后者,清除卵表面的碳氢化合物可以显著提高取食率,表明卵表面的碳氢化合物具有化学防御的功能(Wareetal., 2008; Katsanisetal., 2017)。拟谷盗Triboliumbrevicornis蛹表面的碳氢化合物可以阻碍被同种和同属其它种的捕食,其活性成分主要是二十八烷(Alabietal., 2011)。
2 影响甲虫表皮碳氢化合物组成的因素
2.1 性别
研究显示,甲虫表皮碳氢化合物存在着性别差异,这种差异常常表现在成分的相对含量上,例如在猿叶甲和三种跳甲Altica(Geiselhardtetal., 2009; Xueetal., 2016b)。而在天牛Pidoniagrallatrix、Pidoniatakechii和Xylotrechuscolonus,以及萤火虫Ellychniacorrusca,雌雄之间表皮碳氢化合物不但存在相对含量的差异,也存在成分种类的差异(Akinoetal., 2001; Ginzeletal., 2003a; Tanigakietal., 2007; Ming and Lewis 2010)。
2.2 发育阶段/繁殖状态
拟谷盗Triboliumdestructor幼虫和成虫的表皮碳氢化合物组成和相对含量均存在差异。成虫中正庚烷和3-甲基正庚烷含量最高,幼虫则以正构烷烃为主,幼虫含有正构烷烃,其中的C14-C17为成虫中所不存在的(Hebanowskaetal., 1990)。埋葬甲Nicrophorusvespilloides的繁殖状态对表皮碳氢化合物成分的相对含量有影响,和不育幼的个体相比,繁殖状态的个体具有更高含量的不饱和碳氢化合物成分(Steigeretal., 2007)。对3种跳甲Altica的研究显示,初羽化和性成熟状态的同性个体也存在显著的种内差异,这种种内个体的差异可以一定程度被雄虫作为交配识别的信号上(Xueetal., 2016b)。和正常阶段相比,滞育阶段的二星瓢虫Adaliabipunctata存在特异性的碳氢化合物成分(Sussetetal., 2013)。
2.3 食物
多个研究表明,食物对甲虫的表皮碳氢化合物指纹图谱具有明显影响。寡食性的猿叶甲取食大白菜和取食西洋菜(watercress)时表皮碳氢化合物表现出显著差异,并且这种碳氢化合物指纹图谱可随着饲喂植物的互换而发生逆转(Geiselhardtetal., 2012);进一步的研究发现,食物内脂肪酸的差异会影响叶甲P.cochleariae中直链和甲基化碳氢化合物的含量(Otteetal., 2014)。在饲喂不同的食物时,豆象Acanthoscelidesobtectus表皮碳氢化合物的指纹图谱表也现出可塑性(Stojkovic'etal., 2014)。两种近缘的专食性跳甲(Altica属)寄主植物相互隔离,均不取食对方的植物,在实验室条件下可实现种间杂交,其杂交一代可取食两个亲本的寄主植物而完成个体发育,当杂交一代用两种植物分别饲喂时,其表皮碳氢化合物表现出显著差异(Xueetal., 2016a)。
在上述情况下,甲虫的基因并未发生改变,但寄主植物的改变会影响昆虫的交配信号,即表皮碳氢化合物的成分,产生植食性昆虫的行为隔离,从而导致植食性昆虫的生殖隔离和物种形成,这在物种分化的初始阶段可能起到重要作用(Geiselhardtetal., 2012; Xueetal., 2016a)。
2.4 季节/温度
研究表明,甲虫表皮碳氢化合物的指纹图谱存在季节性变化。例如,拟步甲Eleodesarmata(Hadley, 1977)和异色瓢虫(Durieuxetal., 2013)表皮碳氢化合物的相对含量存在季节性差异;温度对拟步甲Eremostibesbarbatus表皮碳氢化合物的相对含量也具有一定程度的影响(Geiselhardtetal., 2006)。
3 碳氢化合物生物合成的分子机制
昆虫表皮碳氢化合物的合成场所是位于腹部的绛色细胞,其合成过程涉及到四步:(1)通过脂肪酸合成酶形成直链饱和、非饱和或者甲基支链脂肪酸前体;(2)通过延长酶将脂肪酸延伸形成长链脂肪酰辅酶a;(3)通过脂肪酰基辅酶a还原酶将长链脂肪酰辅酶a转化成醛;(4)通过氧化脱羧反应形成碳氢化合物(Blomquist and Bagnères, 2010; Ginzel and Blomquist, 2016)。
目前,关于碳氢化合物生物合成的分子机制的研究大多基于果蝇Drosophilamelanogaster和家蝇Muscadomestica等模式生物。昆虫碳氢化合物合成的最后一步受细胞色素P450超基因家族成员的调控(Reedetal., 1994)。细胞色素P450的4G家族为昆虫所特有,迄今为止已经测序的昆虫基因组都至少有一个CYP4G基因(Feyereisen, 2012)。其中山松大小蠹Dendroctonusponderosae中存在两个氧化脱羧反应的候选基因,即CYP4G55和CYP4G56。研究表明,脂肪乙酰辅酶a还原酶催化还原脂肪乙酰辅酶a生成醇,然后由CYP4Gs催化氧化生成醛,再氧化脱羧形成碳氢化合物,CYP4Gs可将短链的醇和醛转化成碳氢化合物。研究也证明了CYP4G酶具有高保守性,可以催化不同长度的碳链(MacLeanetal., 2018)。在另一项研究中,采用RNA干扰技术验证了CYP4G家族中两个基因(TmCYP4G122和TmCYP4G123)在黄粉虫Tenebriomolitor表皮碳氢化合物合成中的催化作用,发现沉默掉这两个基因中的任意一个或者同时沉默掉两个基因,在幼虫中或蛹中的表皮碳氢化合物含量均会显著减少(Wangetal., 2019)。
4 表皮碳氢化合物在化学分类中的应用
昆虫表皮碳氢化合物组成是昆虫化学分类的分类特征之一。近年来,昆虫表皮碳氢化合物在昆虫化学分类中的应用已经取得长足进展(Kather and Martin, 2012; 李群臣等, 2019)。
其中一类工作是将表皮碳氢化合物用于争议和疑难物种的鉴定及分类。例如,两种拟谷盗Triboliumcastaneum和T.confusum的形态和表皮碳氢化合物组成均相似,但其碳氢化合物成分相对含量的差异可以用来区分两个物种(Lockey, 1978);Sullivan等对墨西哥地区小蠹Dendroctonusfrontalis的两种不同形态类型的表皮碳氢化合物进行定量分析,结果发现不同形态类型的D.frontalis是两个独立的物种(Sullivanetal., 2012);3种近缘跳甲(蓟跳甲、蛇莓跳甲和老鹳草跳甲)的表皮碳氢化合物表型在种间和种内(雌雄之间,初羽化和性成熟个体之间)均存在差异,但统计分析结果表明其种间差异显著大于种内差异,该化学特征可作为近缘物种鉴定和界定的依据(Xueetal., 2016b)。类似的工作在萤叶甲、天牛和象甲等类群也有开展(Zhangetal., 2014; 王书平等, 2017; Duffyetal., 2018; Chenetal., 2019)。
另一类工作是将表皮碳氢化合物数据用于揭示甲虫的亲缘关系。例如,Lockey and Metcalfe(1988)对22种拟步甲表皮碳氢化合物的组成和含量进行分析,并尝试以聚类分析的方法对种间关系进行了探讨;Pageetal.(1997)对小蠹Ips属Grandicollis亚属的7个种进行了表皮碳氢化合物成分的鉴定,并基于系统发育的方法对种间关系进行了分析,由此对一些疑难种的系统分类地位提出了见解,发现基于表皮碳氢化合物数据得到的系统关系与基于形态学等其它方法得出的结论大致吻合;近期一项对地中海地区12种蜣螂的分析结果表明,表皮碳氢化合物具有种特异性,聚类分析结果显示,部分物种自身的所有个体聚在一起,聚类分析的结果和基于分子标记的系统发育结果部分吻合,说明了表皮碳氢化合物在一定程度上可以作为蜣螂物种分类和亲缘关系推定的工具(Niogretetal., 2019)。
5 结语与展望
甲虫种类繁多,目前仅有一小部分种类的表皮碳氢化合物被测定,对表皮碳氢化合物在不同类群中的功能尚知之甚少。比如,哪些甲虫类群利用表皮碳氢化合物作为性信息素仍不十分清楚。由于甲虫表皮碳氢化合物组成成分非常复杂,多数情况下,行为活性成分测定存在困难,除了部分重要林木害虫的研究较为透彻外,目前对大多数种类均不清楚。
对于一些甲虫的类群,表皮碳氢化合物在物种分化及生殖隔离中发挥重要作用。对于这类以表皮碳氢化合物作为性信息素的类群而言,碳氢化合物合成相关的基因可能受到性选择的影响,而对于不以碳氢化合物为信息素的类群而言,受到环境选择方面的压力更大,二者可能存在不同的演化机制,但目前这方面的研究鲜有涉及。鉴于上述原因,表皮碳氢化合物在昆虫物种界定和系统发育推断中的应用价值可能因类群而异。
在化学分类方面,由于昆虫表皮碳氢化合物存在种内变化,如何界定种内变化和种间差异是一个需要思考的问题;另外,不同的提取方法和分析方法,包括不同的溶剂和不同的色谱柱均可能导致结果的差异,因此不同研究之间的比较难以进行,怎样建立标准化的提取和分析规范是一个值得关注的问题。
以甲虫为对象进行表皮碳氢化合物生物合成机制/途径的研究还不多见,其主要活性成分差异的分子机制尚不明了。比如,蛇莓跳甲和老鹳草跳甲种间存在显著的行为隔离,这个生殖隔离是由表皮碳氢化合物的种间差异介导的(Xueetal., 2016a),那么哪些基因改变或以怎样的方式进行调控,从而导致二者表皮碳氢化合物指纹图谱的变化及其相应的选择行为的改变是一个非常值得探讨的问题。
猜你喜欢
植物保护(2021年4期)2021-11-12
四川蚕业(2021年1期)2021-02-12
红蜻蜓·低年级(2020年9期)2020-09-26
华夏地理(2019年2期)2019-07-24
小学生作文(低年级适用)(2018年4期)2018-11-29
理科考试研究·高中(2016年5期)2016-05-14
小朋友·快乐手工(2015年10期)2015-11-02
百科知识(2015年7期)2015-04-08
小朋友·快乐手工(2015年2期)2015-03-13
江苏农业科学(2014年3期)2014-07-16
