
遮阴对烤烟烟碱合成调控的影响
2022-03-23 06:28罗俊周宏钱发聪李军营
中国农业科技导报 2022年2期
罗俊,周宏,钱发聪,李军营
(1.南京农业大学生命与科学学院,南京 210032;2.云南烟草农业科学研究院农艺部,云南 玉溪 653100;3.中国烟草总公司湘潭中等专业学校,湖南 湘潭 411103;4.云南省烟草昆明市公司,昆明 650051)
光照强度是生态系统过程的基本驱动因素,影响作物光合作用和初级生产速率[1]。光照强度在农业生态系统中作物的生长和发育方面起着重要作用,其变化可能导致作物中化学物质合成代谢的重新调整[2-3]。优质烤烟生长需要充足且适宜的光照[4],在云南6—7月,此时云烟87正处于烤烟旺长期,烤烟所需适宜光强为3.0~5.0万lx,光照不足会导致烤烟株高降低,叶片长和宽等农艺性状指标降低,不利于烟碱合成,从而降低烤烟烟叶内在品质[4-5]。
弱光照对烟株生长特性有显著影响[6]。适宜的遮阴可调节烟叶的蒸腾速率、气孔导度和胞间CO2浓度,有利于烟叶的光合作用,促进烟叶内含物的形成[7]。刘柏林等[6]研究表明,遮光显著降低烟株HY06和K326株高和茎围,现蕾期有效叶片数显著降低,烟叶净光合速率、气孔导度、叶片光合放氧速率显著降低,胞间二氧化碳浓度显著升高。弱光照对烟株化学物质合成代谢有显著影响[8]。强光照处理会使烤烟植株中烟碱总含量降低,中上部烟叶尤其如此[9]。研究表明,适度遮阴会提高烟叶烟碱含量,重度遮阴到采收后期烟碱含量则降低,而3层白色聚乙烯纱网遮阴的上部叶和中部叶烟碱含量维持在一个比较稳定的范围[10]。适宜的光照有利于提高烤烟评吸质量,初烤烟叶化学成分更协调,利于优质烟叶的形成,而弱光照条件显著影响烤烟叶片烟碱水平[11]。前期研究仅局限于光照强度对烟株农艺性状、光合参数及常规化学成分含量的影响[12-15],但遮阴度对烤烟烟碱合成的影响机理仍不明确。
本课题组推测遮阴对烟叶烟碱含量影响是由烟碱前体、中间物质含量及烟碱合成酶基因相对表达水平的差异所致。为了验证此假设,本研究在大田试验中通过覆盖不同层数遮阴网以设置不同遮阴强度,从烟碱合成通路的角度分析遮阴度对烟碱前体、中间物质合成及烟碱合成酶基因表达量的影响,进一步探究遮阴对烟碱合成的影响机制,以期为烤烟烟碱的合成调控提供理论依据及生产指导。
1 材料和方法
1.1 试验区概况
试验时间为2018年10月到2020年8月(包括预实验),试验地设在云南省烟草农业科学研究院研和基地。供试红壤情况:pH为6.2,速效氮8.04 mg·kg-1,速效磷 12.6 mg·kg-1,速效钾 70.4 mg·kg-1。供试材料为云南主栽烤烟品种云烟87。
1.2 试验设计
不同遮光强度通过覆盖不同层数的遮阳网(尼龙网,孔径50 mm,透光率为60%)来实现。试验设4个处理,对照组(CK)为100%自然光,光强在 1 250~2 000 μmol·m-2·s-1;L1处理遮 1层遮阳网,控制光强分别在 890~1 380 μmol·m-2·s-1,约70%自然光,轻度遮阴;L2处理遮2层遮阳网,控制光强分别在 560~910 μmol·m-2·s-1,约 45% 自然光,中度遮阴;L3处理遮3层遮阳网,控制光强分别在 260~420 μmol·m-2·s-1,约20%自然光,重度遮阴。各处理设置3个重复,随机区组排列。烤烟采用漂浮育苗,于6叶1心时移栽。在移栽20 d后进行遮阳网覆盖处理,遮阴40 d后打顶,直至遮阴后75 d。烤烟大田试验管理按照云南省优质烤烟生产栽培管理技术标准执行。
1.3 测定项目和方法
1.3.1 光合特性测定 光合特性的测定时间为遮阴后20 d(旺长期)、40 d(打顶期前1 d)、60 d(下部叶采烤期前1 d)的10:00—12:00,分别选取倒第6、8、10片叶,使用LI-6400XT便携式光合作用测量系统(美国LI-COR公司)测定。光合特性测定指标包括净光合速率(μmol·m-2·s-1)、胞间CO2浓度(μmol·mol-1)、气孔导度(mmol·m-2·s-1)、蒸腾速率(mmol·m-2·s-1)等。
1.3.2 烟碱及烟碱前体、中间物质含量测定 分别于烟株遮阴处理后15、25、35、45、55、65、75 d(除烟碱中间物质含量)取样,进行生化指标的测定。用连续流动分析仪[法国Alliance(AMS集团)公司]测定云烟87根、茎、叶的烟碱含量。用Agilent 1100 HPLC高效液相色谱(美国安捷伦公司)测定根系烟碱前体和中间物质含量。
1.3.3 烟碱合成酶基因表达水平测定 采用TRIZOL法提取0.1 g根尖细胞总RNA,提取方法参照Total RNA Extraction Kit(北京索莱宝科技有限公司)说明书。随后参照Universal RT-PCR Kit(北京索莱宝科技有限公司)说明书合成烟草根尖细胞cDNA。用RT-PCR方法分析烟碱合成酶基因的表达水平,包括腐胺-N-甲基转移酶(putrescine N-methyltransferase)基因NtPMT、喹啉酸磷酸核糖基转移酶(quinolinate phosphoribosyl transferase)基因NtQPT、N-甲基腐胺氧化酶(N-methylputrecine oxidase)基因NtMPO和精氨酸脱羧酶(arginine decarboxylase)基因NtADC。RTPCR反应采用荧光染料SYBR Green I(上海萌芽生物科技有限公司)在ABI7500荧光定量PCR仪上进行,PCR程序如下:95℃30 s预变性;95℃5 s,60℃34 s,循环40次。然后用熔解曲线检验扩增效率,基因引物用PrimerPremier5.0设计,见表1。最后用2-△△Ct的方法计算基因的相对表达水平。

表1 荧光定量PCR引物设计Table 1 List of primers used for the qRT-PCR analysis
1.4 数据分析
应用SPSS 24.0软件对试验中所得到的数据做Duncan多重检验(Duncan’s multiple range test),采用Origin2018作图。
2 结果与分析
2.1 不同遮阴处理对烤烟叶片光合特性的影响
由表2可知,遮阴20 d后,与对照相比,L1处理的烟叶净光合速率、气孔导度、胞间CO2浓度分别增加10.42%、35.85%、5.44%,而L3处理的相应指标分别降低38.80%、52.83%、3.89%,表明适当遮阴有助于烤烟叶片进行光合作用,重度遮阴时,光合作用反而降低,不利于烤烟生长。遮阴40 d后,烤烟叶片净光合速率、气孔导度、蒸腾速率随遮阴度增大而逐渐降低,L3处理降幅最大,分别为39.94%、71.43%、40.41%。遮阴处理60 d与遮阴处理40 d的相应光合生理指标结果相反。所有处理烟叶胞间CO2浓度随烤烟生育期的推进呈先降后升趋势,整体呈单峰曲线,这可能是由遮阴40 d后烤烟处于现蕾期光合作用较大所致。
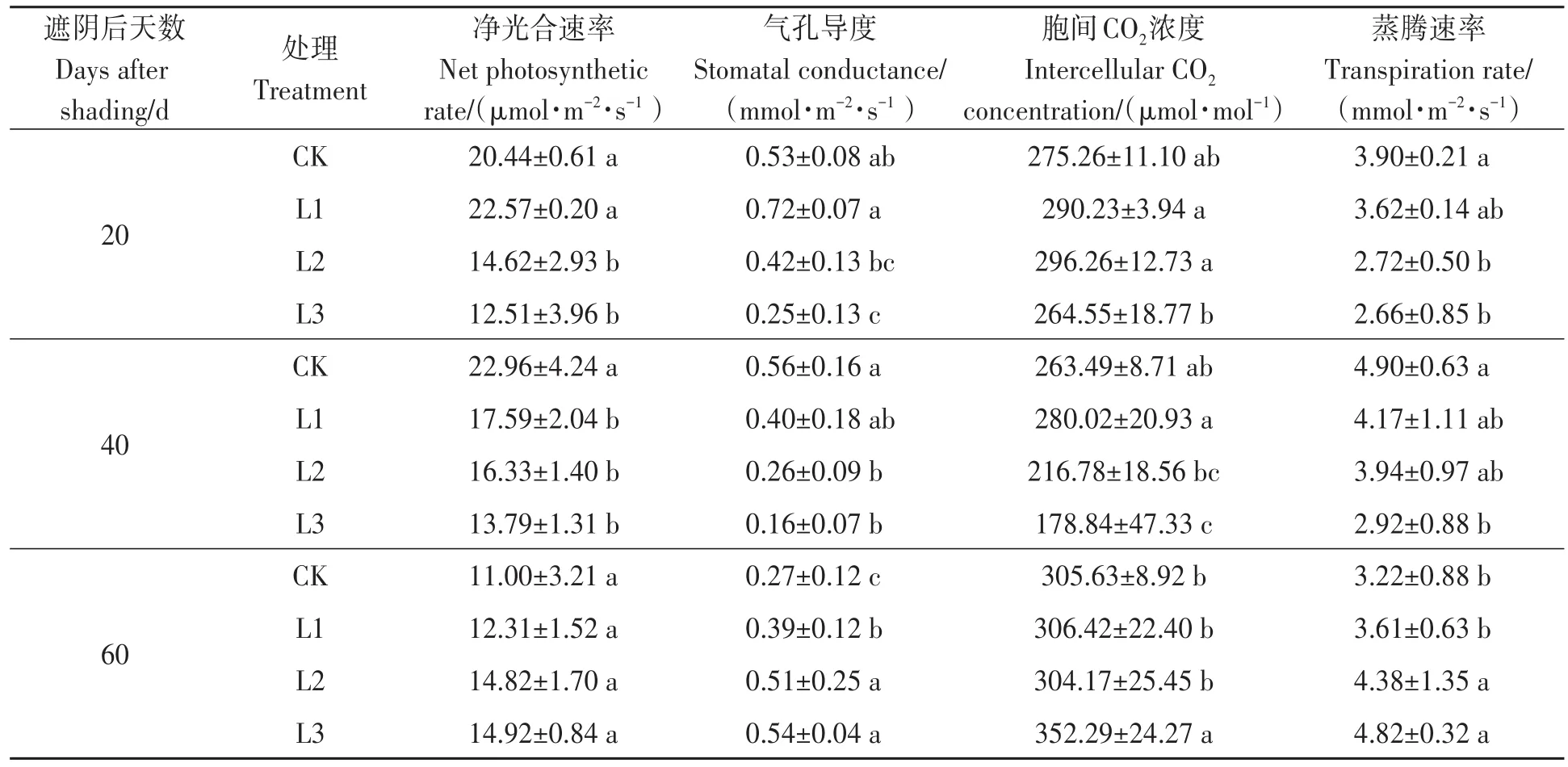
表2 不同遮阴处理对烤烟中部腰叶光合特性的影响Table 2 Effect of different shading treatments on photosynthetic characteristics of middle waist leaf of flue-cured tobacco
2.2 不同遮阴处理对烤烟各器官中烟碱含量的影响
图1显示,遮阴15~55 d,根部烟碱含量随生育期推进呈现先升高、后降低、再升高、最后降低的趋势,即呈双峰曲线。遮阴后45 d,由于打顶操作刚完成,处理L1、L2、L3的根部烟碱含量比对照高,L2处理增幅最大,为53.19%。遮阴55~75 d,处理L3的根部烟碱含量最低,与对照相比分别降低3.33%、16.92%、11.48%,表明重度遮阴降低现蕾期烤烟根系烟碱含量。遮阴15~55 d,与对照相比,L3处理显著降低烤烟茎部烟碱含量,分别为29.87%、43.97%、15.83%、21.05%、6.91%。对下部叶而言,遮阴后15 d,叶片烟碱含量随遮阴度增大而逐渐降低,最大降幅为45.05%;而遮阴75 d后叶片烟碱含量与遮阴15 d后的结果相反,最大增幅为160.44%。对中部叶而言,除遮阴45、65 d后,与对照相比,L1、L2处理烤烟叶片烟碱含量各生育期均无显著差异,表明轻中度遮阴对中部叶烟碱含量几乎无影响。值得注意的是,烤烟各器官中,所有处理在遮阴55 d时,烟碱含量与其他天数相比都较低,说明处于现蕾末期的烤烟烟碱合成在急剧降低。
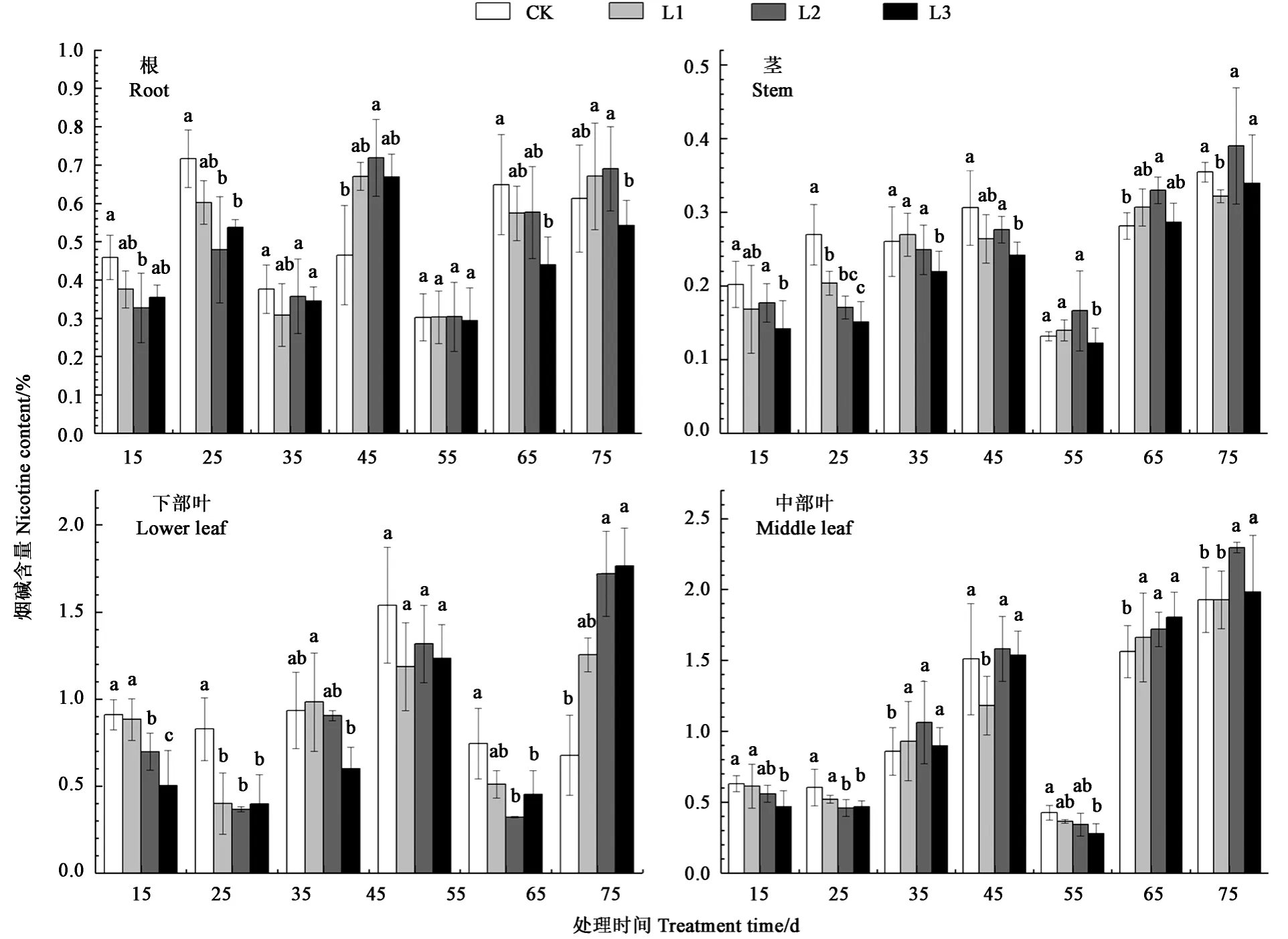
图1 不同遮阴处理对烤烟各器官烟碱含量的影响Fig.1 Effect of different shading treatments on the nicotine content in organs of flue-cured tobacco
2.3 不同遮阴处理对烤烟根部烟碱中间物质含量的影响
从图2结果可知,遮阴15~45 d,与对照相比,L1处理降低根部腐胺含量分别为36.77%、11.37%、2.84%、5.71%,但差异不显著。而遮阴55~65 d,L1、L2处理根部腐胺含量都高于对照CK,L2处理增幅最大,分别为14.31%、6.44%。遮阴25~65 d,与对照相比,所有遮阴处理烤烟根部精胺含量都较高,说明遮阴有利于精胺的合成。CK、L1、L2处理的根部精胺含量随生育期推进呈先升、后降、再升趋势,呈双峰曲线,且分别在遮阴后45、65 d达到高峰值。遮阴15~25 d,与对照相比,L1处理的根部亚精氨含量分别降低30.47%、4.46%,而L2、L3处理显著增加亚精氨含量,分别是对照的2.16、1.99倍。遮阴35~65 d,与对照相比,L1、L2、L3处理的根部亚精氨含量增加,说明烤烟旺长期以后遮阴处理有利于亚精氨合成,最大增幅分别为147.64%、173.78%、73.89%、82.69%。遮阴25~65 d,与对照相比,L2、L3处理显著增加多胺含量,最大增幅分别为 40.05%、36.14%、36.27%、23.47%、162.40%。
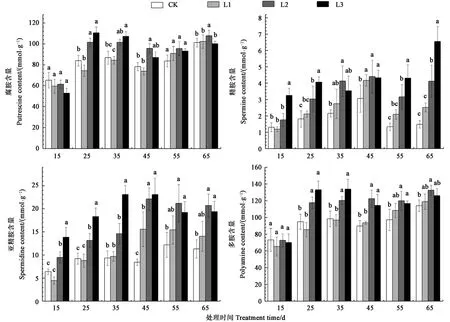
图2 不同遮阴处理对烤烟根系烟碱中间物质含量的影响Fig.2 Effect of different shading treatments on the nicotine intermediate content at root of flue-cured tobacco
2.4 不同遮阴处理对烤烟根部烟碱前体物质含量的影响
图3结果显示,遮阴15~55 d,L2、l3处理的烤烟根部天冬氨酸含量都高于对照,最大增幅分别为7.01%、25.65%、253.85%、22.70%、89.28%。遮阴65~75 d,L3处理降低天冬氨酸含量幅度最大分别为27.20%、37.52%。遮阴15~65 d,所有遮阴处理的根部精氨酸含量随生育期推进呈先降、后升、再降的趋势,第1个低峰值和高峰值分别出现在遮阴后35、45 d。除遮阴35 d外,L1、L3处理甲硫氨酸含量都比对照降低。在遮阴55 d后,所有遮阴处理的烤烟根部甲硫氨酸含量急剧下降,根部烟碱含量急剧下降相呼应。遮阴35~55 d,与对照相比,L2、L3处理烤烟根部游离氨基酸含量都显著增加,最大增幅分别为38.88%、25.67%、20.72%。
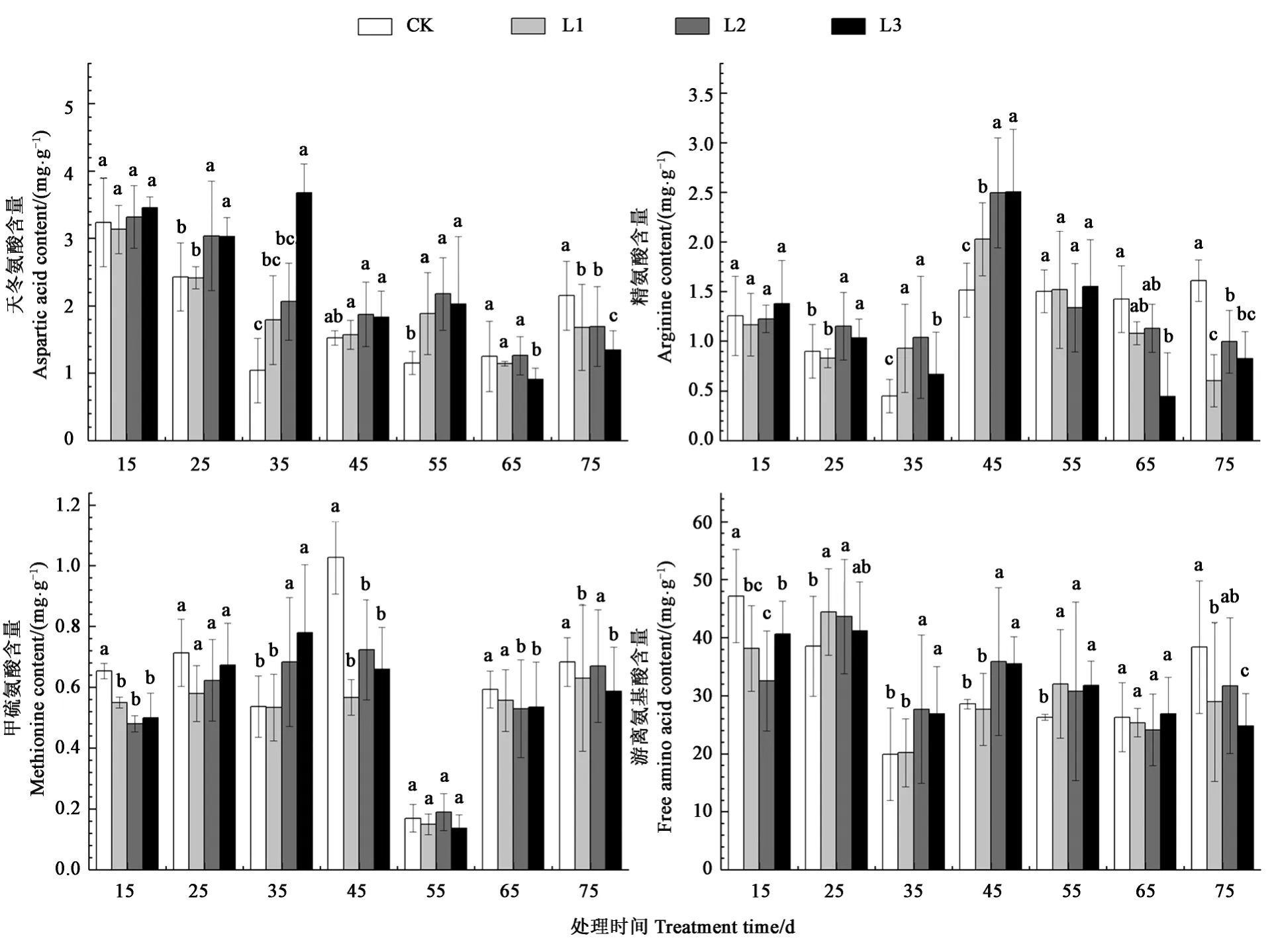
图3 不同遮阴处理对烤烟根系烟碱前体物质含量的影响Fig.3 Effect of different shading treatments on the nicotine precursor content at root of flue-cured tobacco
2.5 不同遮阴处理对烤烟根系烟碱合成酶基因表达的影响
从图4可以看出,遮阴15、55 d后,QPT基因相对表达量随遮阴度增大而逐渐降低,最大降幅分别为50%、83%;遮阴35 d时,所有遮阴处理现蕾期烤烟根系QPT基因相对表达量都高于对照,分别比对照增加74%、31%、27%。遮阴25~55 d,与对照相比,L2或L3处理显著增加PMT基因相对表达量,最大增幅分别为283.80%、85.97%、67.17%、54.59%。遮阴25~65 d,与对照相比,L3处理MPO基因相对表达量显著增加,最大增幅分别为155.62%、164.26%、67.17%、98.59%、34.40%。遮阴15~45 d,与对照相比,L2或L3处理显著增加ADC基因相对表达量,最大增幅分别为85.97%、181.83%、103.93%、54.59%;遮阴45~65 d,与对照相比,L1处理显著增加ADC基因相对表达量,增幅分别为70.28%、110.53%。
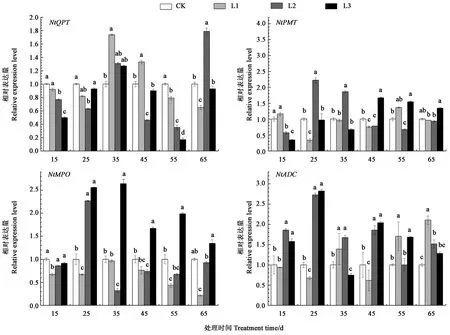
图4 不同遮阴处理对烤烟根系烟碱合成酶基因表达的影响Fig.4 Effect of different shading treatments on nicotinic synthase gene expression at root of flue-cured tobacco
3 讨论
不同遮阴程度对烟株叶片光合特性产生不同影响[16]。王子腾[17]研究认为,烤烟上部叶光合强度、气孔导度和蒸腾速率均随光照强度升高呈先升后降趋势,而本研究结果表明,烤烟旺长期中部叶净光合速率、气孔导度、蒸腾速率随遮阴度增大而逐渐降低,重度遮阴处理降幅最大,各指标降幅分别为39.94%、71.43%、40.41%。这可能是由本研究中光强范围处于烟叶光饱和点以下所致。
烟碱在根系中合成,且根系和叶片中烟碱都能通过去甲基化降解,但主要在衰老叶片中降解[18]。烤烟中烟碱的累积受到环境因素、栽培技术、植物激素水平等影响[19-20]。适度遮阴可提高烟叶烟碱含量,重度遮阴到采收后期烟碱含量则降低,而3层遮阴上部叶和中部叶烟碱含量维持在比较稳定的范围[10]。遮阴15~55 d,根部烟碱含量随生育期推进呈现先升高、后降低、再升高、最后降低的趋势,呈现双峰曲线。第1个高峰值出现在遮阴25 d,可能因为在其中一条通路下NtADC和NtPMT表达量增加,导致由精氨酸合成精胺的含量升高,最后在PMT酶的作用下合成烟碱;另一条通路下随腐胺合成量和NtPMT表达量的增加,烟碱合成量增加,由于两条通路的共同作用导致烟碱合成量急剧增加[21]。第2个低峰值出现在遮阴55 d,可能因为天冬氨酸含量和NtQPT表达量大幅度下降,导致烟碱合成量急剧下降[21]。遮阴25~55 d,中重度遮阴处理增加烤烟根部腐胺、精胺、亚精胺及多胺含量,表明该处理导致前体物质精氨酸含量较低,而精氨酸脱羧酸基因NtADC表达量较高,由此合成的中间物质(多胺类)含量高。遮阴15~55 d,中重度遮阴处理的烤烟根部天冬氨酸含量都高于对照,中重度遮阴最大增幅分别为 7.01%、25.65%、253.85%、22.70%、89.28%,这与刘典三[22]的研究结果基本一致,表明中重度遮阴处理不利于烤烟叶片的氨基酸向蛋白质的同化及蛋白质的转运,降低了氮素同化水平。本研究结果证实,遮阴25~55 d,重度遮阴处理的烤烟根部天冬氨酸、精氨酸、腐氨、精氨、亚精胺含量都高于对照,烟碱含量却降低,可能因为其中一条通路为由天冬氨酸在NtQPT作用下合成烟酸,再合成烟碱;另一条通路为精氨酸经过NtADC合成精胺,再由精胺合成烟碱[23]。当精氨酸、精胺和天冬氨酸含量累积时,烟碱的合成受阻。
烟碱生物合成是代谢途径中多基因共同作用的结果[23-24]。目前为烟碱合成提供吡咯烷环的通路有3条,第1条通路为精氨酸经过ADC酶的作用合成精胺转化为N-氮甲酰基腐胺,其为烟碱合成提供吡咯烷环;第2条通路为由鸟氨酸经过鸟氨酸脱羧酶作用合成腐胺,再由PMT和MPO酶的作用合成N-甲基吡咯啉,其为烟碱提供吡咯烷环;第3条通路为由蛋氨酸经过S-腺苷甲硫氨酸合成酶的作用合成S-腺苷蛋氨酸,再由PMT和MPO酶的作用合成N-甲基吡咯啉,其为烟碱提供吡咯烷环。目前为烟碱合成提供吡啶环的通路有1条,先由天冬氨酸转化为喹啉酸,后在QPT酶的作用下转化为烟酸,其为烟碱提供吡啶环[24]。但是目前尚无关于遮阴度对烤烟烟碱合成调控影响机理的报道。本研究阐明,遮阴15~45 d,与对照相比,中重度遮阴显著增加NtADC基因相对表达量,最大增幅分别为85.97%、181.83%、103.93%、54.59%,由此说明了中重度遮阴处理下精氨酸含量高而精胺含量也高的原因。遮阴25~55 d,中重度遮阴处理显著增加NtPMT基因相对表达量,最大增幅分别为283.80%、85.97%、67.17%、54.59%,且重度遮阴处理显著增加NtMPO基因相对表达量分别为155.62%、164.26%、67.17%、98.59%,由此说明了中重度遮阴处理下腐胺含量高而烟碱含量低的原因。遮阴35 d时,所有遮阴处理烤烟现蕾期根系NtQPT基因相对表达量都高于对照,分别比对照增加74%、31%、27%,由此说明了不同遮阴处理下天冬氨酸含量高而烟碱含量低的原因。
猜你喜欢
现代农村科技(2022年8期)2022-11-18
安徽农业科学(2022年16期)2022-09-02
安徽农业科学(2022年10期)2022-06-06
西藏文学(2020年5期)2020-09-21
消费电子(2020年12期)2020-01-19
中国食品(2018年18期)2018-09-10
发明与创新·大科技(2017年12期)2017-12-22
环球时报(2017-12-20)2017-12-20
科学种养(2017年3期)2017-03-23
董事会(2016年8期)2016-05-14
