
不同水分条件对甘孜州大渡河干旱河谷区巨菌草植株形态的影响
2022-03-22 09:34宋思梦彭彬陈梁婧周旭张茂娟梁蔡佳王欣然
农业与技术 2022年5期
宋思梦 彭彬 陈梁婧 周旭 张茂娟 梁蔡佳 王欣然
(四川民族学院横断山区生态修复与特色产业培育研究中心,四川 康定 626001)
甘孜州位于青藏高原东部,是四川盆地西缘山地向青藏高原过渡的地带,地势高峻,北高南低,中部突岀,东南缘深切,山川平行相间,地域差异显著,大渡河流域在州境内干流长239.2km,河床比降约5.8%,年径流量约153.8亿m3,流域面积约2km2。甘孜州大渡河流域干旱河谷区是我国西南山地一种十分特殊的脆弱生态系统[1],面临生态恢复挑战难度最大、经济稳定发展困难程度最高[2]、垂直气候变异机理最为复杂,有着低阈值生态安全与高风险生态退化的隐患[1]。自20世纪60年代开始,人们对资源的恶意掠夺又加剧了该地区的干旱与退化程度,迫使植被趋于低矮多刺的旱生性演替,林线和稀疏灌草上限抬升,部分森林植被消失,地表温度最高可达75℃,土壤破碎风化愈演愈烈,保水保肥能力极差,严重侵蚀后土壤进一步向恶性循环方向发展,由此引发的崩塌、滑坡、泥石流等自然灾害愈发频繁[3,4],在甘孜州大渡河流域干旱河谷区寻找因地制宜的植被恢复措施亟待开展。前人对该地区的生态恢复研究多以“植树造林”为主,如岷江柏(Cupressus chengiana)、刺槐(Robinia pseudoacacia)、臭椿(Ailanthus altissima)、山杏(Armeniaca sibirica)、梨(Pyrus bretschneideri)、红豆杉(Taxus chinensis)等,栽一棵保一棵,忽略了灌木和草本的巨大生态效益[5]。其中,草本生长快速、覆盖面积广泛、可选材料较多[2,4],用以开展干旱河谷区(造林难度大)生态恢复重建意义较大。
巨菌草(Pennisetum sinese),也叫巨能草,隶属被子植物门,单子叶植物纲,禾本科,狼尾草属(Pennisetum),多年生禾本科直立丛生型草本,是由二倍体珍珠谷作母本与四倍体象草杂交选育而成的三倍体C4植物[6]。原产地在北非,驯化成功后,已在食用菌栽培、动物饲养和生态治理等领域得到广泛应用。其植株高大(株高最高能达到7~8m)、生物量高(单株重达3.25kg,年产鲜草达200~400t·hm-2)、适口性好、根系发达[7-9],种植一次可连续收割15a以上,不仅可用于保持水土,还可作为当地特色畜牧的青(干)饲料以及食(药)用菌栽培原料[10],具有生态、经济、社会综合效益,是我国当代深化农业供给侧结构性改革,推进农业优质、特色、绿色发展,助力山区乡村振兴的重要作物。在甘孜州大渡河流域干旱河谷区引种巨菌草对生态环境保护、退化生态系统修复、畜牧业发展、提高当地农牧民收入意义重大[11]。巨菌草对热量要求较高,较耐贫瘠,生长效果对水分有一定要求[12-14]。选取甘孜州大渡河流域干旱河谷区域代表性土壤,通过设定不同水分梯度的巨菌草盆栽试验,观察其株高、叶宽、叶长的变化过程,拟解决以下科学问题:不同水分条件对巨菌草植株形态是否有影响;若有,不同水分条件对巨菌草植株形态有什么影响;筛选适宜于甘孜州大渡河流域干旱河谷区土壤引种巨菌草的最佳水分条件。以期为甘孜州大渡河流域干旱河谷区脆弱生态系统引种巨菌草进行植被恢复提供科学依据。
1 材料与方法
1.1 研究地概况
甘孜州位于四川西部,主要流域有金沙江、雅砻江、大渡河,其中大渡河流域主要途径丹巴县、康定市、泸定县等区域(E101°17′~102°38′,N29°39′~31°23′)[15]。属于典型的干旱河谷气候,辐射强烈,干湿季分明,焚风效应显著。年平均日照时数1324~2079h,年平均气温7.0~15.5℃,年平均无霜期177~279d,年平均降雨量69.35~664.4mm,年平均蒸发量1500~2200mm,是降雨量的2~7倍,降水少而蒸发大,地形的高山峡谷闭塞形式与气流团下沉增温减湿,加之人为干扰植被,使得干旱程度加剧。土壤贫瘠,水土流失严重,岩石易在外营力频繁作用下风化剥离。研究选择来自泸定德威村的典型黄壤,在康定黑日村四川民族学院农学楼3楼温室大棚(温湿度受控并遮蔽雨水,无降雨干扰)中进行盆栽试验,该地区(E102°17′,N30°12′)海拔1400m,是典型的干旱河谷气候。
1.2 试验材料
于2021年5月从福建农林大学菌草综合开发利用技术国家地方联合工程研究中心引进4月龄健康、成熟、无病虫害的饱满巨菌草,将其砍切为20cm左右的种节。土壤取自泸定德威,其田间持水量25.6%,容重1.43g·cm-3,通气度31.7%,总孔隙度43.7%,毛管孔隙度28.4%。
1.3 试验方法
1.3.1 育苗
砍切后经低浓度尿素浸泡、高温高湿催芽,扦插1节于1个聚乙烯圆台型花盆(口径40cm,深25cm)中,共扦插90盆,每盆装16kg土壤,均放置于温室大棚中自然育苗。育苗1月后选择长势相近、健壮无病害的巨菌草幼苗60株进行水分控制试验。
1.3.2 水分控制试验
采用盆栽控水的方法。清除盆中杂草等因素,采用控制变量法,设置水淹胁迫(100%田间持水量)、正常(75%田间持水量)、轻度干旱胁迫(50%田间持水量)、重度干旱胁迫(25%田间持水量)4个水分梯度。自6月开始,采用每日称重补水与土壤水分仪结合的方式控制盆中土壤水分含量。期间不施肥,以便监测水分条件的单因素对巨菌草生长影响。
1.4 指标监测
每7d(6月16日、6月23日、6月30日、7月7日、7月14日)监测每盆巨菌草的株高、叶宽、叶长重要植株形态指标。其中,株高用皮尺测定从根颈至叶心的垂直高度;叶宽用钢卷尺测量植株顶端已完全展开的第3片叶的宽度;叶长用钢卷尺测量植株顶端已完全展开的第3片叶的长度。每个梯度选择代表性植株4株,连续时间对其进行测量,每个梯度取平均值。
1.5 数据处理
运用Microsoft Excel 2007进行数据整理;运用SPSS 24.0计算数据平均值和标准误差;运用One-way ANOVA验证不同次测量前后植株形态差值变化及不同水分梯度对植株形态特征的影响,利用Levene′s test检验方差齐性与否,方差齐性时差异程度用Duncan多重比较,反之则用T2 Tamhane′s test多重比较,显著性水平设置为P=0.05;运用Origin 9.0制图。
2 结果与分析
2.1 巨菌草植株形态受水分条件调控的总体变化
巨菌草株高、叶宽、叶长受水分条件变化的显著影响(P<0.05),水分的增减调控着巨菌草植株形态的变化特征。其中,在正常水量(75%田间持水量)状态下巨菌草累计株高增量最大(70.45±22.82cm)、累计叶长增量最大(31.58±13.67cm),在水淹状态(100%田间持水量)状态下巨菌草累计叶宽增量最大(1.17±0.41cm),而在重度干旱胁迫(25%田间持水量)状态下巨菌草死亡。从株高增额来看,正常水量组>水淹组>轻度干旱胁迫组>重度干旱胁迫组,均有显著性特征;从叶宽增额来看,水淹组>正常水量组>轻度干旱胁迫组>重度干旱胁迫组,均有显著性特征;从叶长增额来看,正常水量组>水淹组>轻度干旱胁迫组>重度干旱胁迫组,均有显著性特征。因此,不同水分条件对巨菌草植株形态的影响中,正常水量组巨菌草生长最好,水淹组其次,轻度干旱胁迫组次之,重度干旱胁迫组巨菌草无法正常存活。
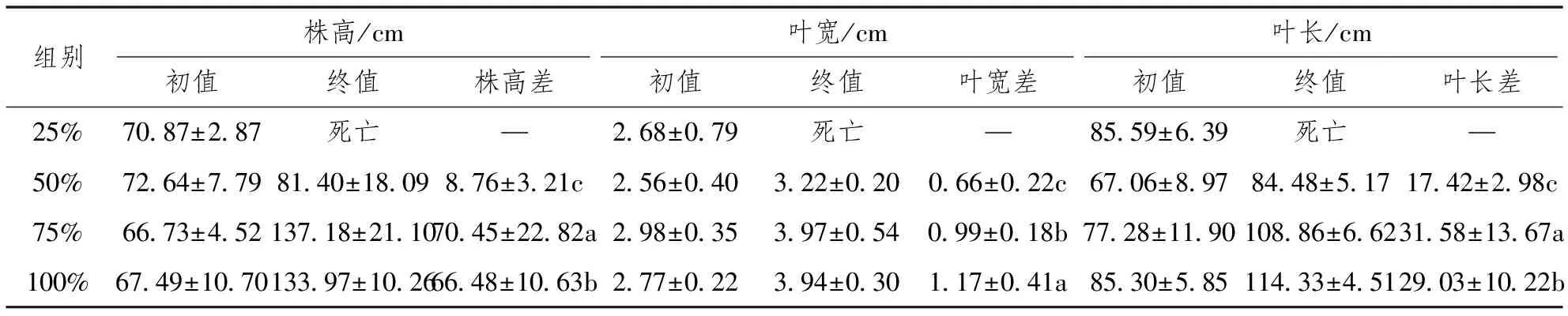
表1 不同水分条件对甘孜州大渡河干旱河谷区巨菌草植株形态的影响
2.2 巨菌草株高差受水分条件调控的变化
追踪水分调控后巨菌草累积株高生长随时间的变化如图1所示。分析图中数据可知,各水分梯度下巨菌草株高差随时间先增大后减小,且正常水量组和水淹组增幅更明显,累积量更高,阶段性增幅最大可达到29.10cm(6月30日—7月7日水淹组),而轻度干旱胁迫组在6月30日—7月7日出现负增长(-10.12cm),说明轻度干旱胁迫已经出现使巨菌草凋萎的现象。此外,重度干旱胁迫组在第1个7d(6月16—23日)内缓慢积累株高,此后逐渐死亡,无观测值。
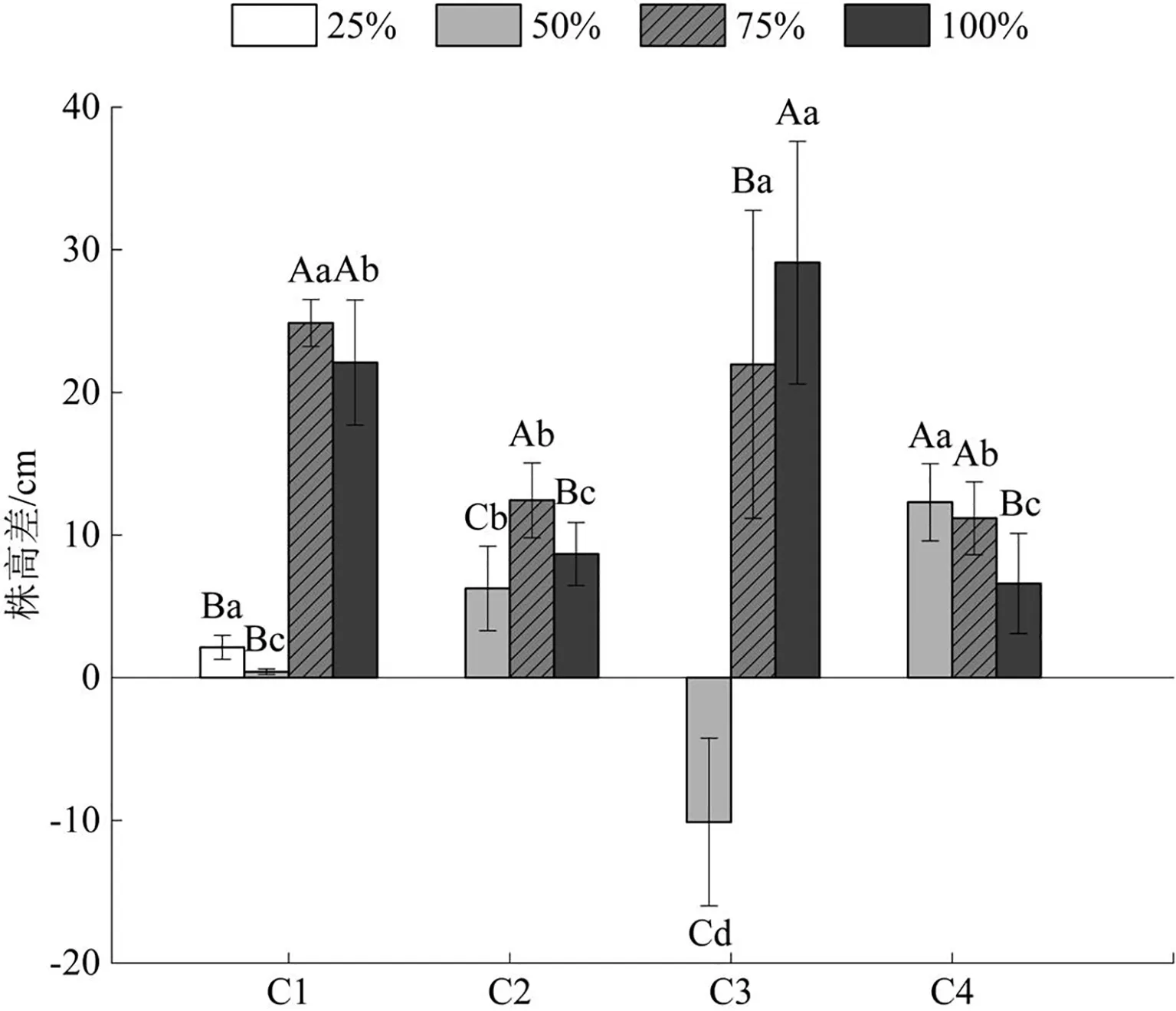
图1 不同水分条件对甘孜州大渡河干旱河谷区巨菌草株高差的影响
2.3 巨菌草叶宽差受水分条件调控的变化
追踪水分调控后巨菌草累积叶宽生长随时间的变化如图2所示。分析图中数据可知,各水分梯度下巨菌草叶宽差随时间变化不一,正常水量组先增后减,在6月30日—7月7日有高峰值;水淹组先增后减,在6月23—30日有高峰值;轻度干旱胁迫组先增后减,在6月30日—7月7日有高峰值。且正常水量组和水淹组变幅更明显,总累积量更高,阶段性增幅最大可达到0.68cm(6月30日—7月7日正常水量组),而轻度干旱胁迫组在6月16—30日出现负增长,说明轻度干旱胁迫已经出现使巨菌草叶片萎焉的现象,而后随巨菌草渗透性物质调节作用,从6月30日—7月7日逐渐恢复。此外,重度干旱胁迫组在第1个7d(6月16—23日)内叶宽急剧缩减,此后逐渐死亡,无观测值。
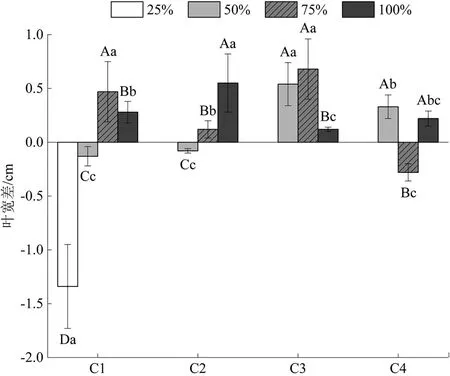
图2 不同水分条件对甘孜州大渡河干旱河谷区巨菌草叶宽差的影响
2.4 巨菌草叶长差受水分条件调控的变化
追踪水分调控后巨菌草累积叶长生长随时间的变化如图3所示。分析图中数据可知,各水分梯度下巨菌草叶长差随时间变化不一,正常水量组逐渐减少,在6月16—23日有高峰值;水淹组呈先减后增再减的“W”型,在6月16—23日有高峰值;轻度干旱胁迫组先增后减,在6月30日—7月7日有高峰值。且正常水量组和水淹组变幅更明显,总累积量更高,阶段性增幅最大可达到15.64cm(6月16—23日正常水量组),而轻度干旱胁迫组在7月7—14日出现负增长,说明轻度干旱胁迫已经出现使巨菌草叶片萎焉的现象。此外,重度干旱胁迫组在第1个7d(6月16—23日)内叶长急剧缩减,此后逐渐死亡,无观测值。
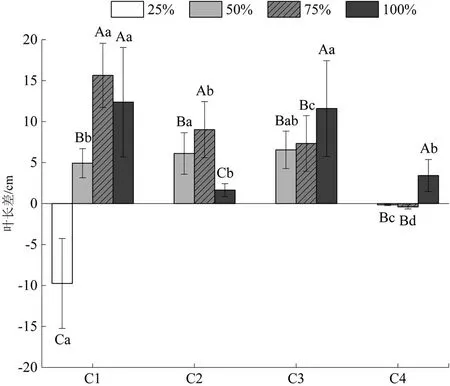
图3 不同水分条件对甘孜州大渡河干旱河谷区巨菌草叶长差的影响
3 结论与讨论
巨菌草植株形态(株高、叶宽、叶长)均受水分调控的显著影响。甘孜州大渡河流域干旱河谷区引种巨菌草有一定耐水淹、耐干旱胁迫的能力,在50%田间持水量的轻度干旱胁迫中可继续生长,但25%田间持水量的重度干旱胁迫中无法正常生长,超过7d后逐渐死亡。在该地区栽培巨菌草时正常水量(75%田间持水量)生长最好,水淹(100%田间持水量)其次,轻度干旱胁迫(50%田间持水量)次之。
巨菌草是福建农林大学国家菌草工程技术研究中心近年来选育的产量高与抗逆强的新草种,通过在黄河沿岸护土固沙与修复生态的集成示范,显示了良好成效与应用前景,实现了当年种草即可发挥防控水土流失的功效。其根系可延伸至地下2m,且每株须根可覆盖直径1m以上,抱沙聚土,充分发挥了生态修复与保土固沙作用,展示了大规模应用潜力与实施草牧菌业延伸开发的良好效益。甘孜州水土流失严重,草地荒漠化速度加剧,土壤有机质锐减,土壤肥力水平持续降低,大渡河流域干旱河谷区受冬夏海陆季风交替影响,干湿季分明,热量下沉,水分降低,水文状况恶化[2,3]。水淹胁迫与干旱胁迫是干旱河谷区雨季及旱季时常出现的严酷逆境挑战,可限制植物的生长[1],尤其是长达8个月的旱季(9月—4月)。研究区土壤容重较大,土壤紧实且土壤孔隙度小,说明土壤入渗速率和蓄水量较低,土壤的通透性可能较差,而在此土壤中发生侵蚀的可能性变大,危害程度更强烈,退化趋势越强[16-18]。引种巨菌草后,其地下根系生长迅速,能在较短时间内形成须根网络,牢固地锁住水分和泥土,防止水土流失,对甘孜州绿化荒山荒坡、防风固沙都具有积极作用;且根系生物量发达,盘根错节,对土壤团聚体具有一定固持作用[19,20],可提高土壤保水性,同时其自身对干旱环境会促进分泌防御系统的保护酶来提高抗逆性[21],这些都将表现在其植株形态特征上。轻度的干旱胁迫中,巨菌草的生长发育无明显受限,而重度干旱胁迫使巨菌草活性氧和防御系统的动态平衡受到破坏,地上部分(叶、茎等)受害症状加重[22-24],失去弹性,色泽变暗,植株损伤直至死亡。后期应保持观测物候期及其它农艺性状,并增加电解质渗透率、相对电导率、丙二醛、可溶性糖、过氧化物酶、过氧化氢酶、超氧化物歧化酶、脯氨酸、可溶性蛋白等生理生化指标的监测,以更明晰地了解巨菌草抗旱抗涝的响应机理。此外,巨菌草抗旱性是较为复杂的综合性状表现,不仅与干旱胁迫程度相关,还可能与植物生育期及干旱胁迫时间有关,今后需增加幼苗期、分蘖期、拔节期、成熟期的巨菌草对干旱胁迫的抵抗能力进一步的研究。
猜你喜欢
小学生学习指导(高年级)(2023年10期)2023-12-10
汽车实用技术(2022年16期)2022-09-03
高原山地气象研究(2020年3期)2020-07-16
小哥白尼(军事科学)(2018年3期)2018-06-15
现代园艺(2017年23期)2018-01-18
廉政瞭望(2016年13期)2016-08-11
河南科技(2015年3期)2015-02-27
湖北科技学院学报(2014年6期)2014-07-12
天然气勘探与开发(2012年2期)2012-03-25
军事历史(1986年2期)1986-01-18
