
新型冠状病毒(SARS-CoV-2)变异的研究进展
2022-03-18 10:01:18蒋潘虹阮嘉雯俞慕华卢次勇
中国人兽共患病学报 2022年2期
蒋潘虹,阮嘉雯,俞慕华,卢次勇
2019年12月底,湖北省武汉市暴发不明原因肺炎疫情。2020年1月,科学家们分离患者体内的病毒,并进行基因组测序,发现是一种新型的β冠状病毒,且与严重急性呼吸道综合征冠状病毒(SARS-CoV)具有78.7%的同源性[1]。基于其与SARS-CoV在基因上的关联性,2020年2月11日,国际病毒分类委员会将该病毒命名为“SARS-CoV-2”,世界卫生组织将感染该病毒引起的疾病正式命名为“COVID-19”[2]。2020年3月初,研究者在S蛋白变异分析中唯一确定G614为符合他们所设定阈值标准的位点。当时,G614形式在全球范围内还很少见,但在欧洲越来越突出;同时全球共享流感数据倡议组织(GISAID)也在追踪携带 D614G 替代的进化分枝,并将其命名为“G clade”[3]。随着时间推移,全世界公布的SARS-CoV-2的基因组信息迅速增加,越来越多的基因突变和突变株被发现。2021年2月25日,世界卫生组织发布了关于“关切变异株”VOC和“关注变异株”VOI的定义,其中VOC应符合:1)流行病学上传播力增强或流行特点出现有害变化;2)致病力增强或临床表现趋重,或公共卫生、社会措施或现有诊断、疫苗、治疗方法的有效性降低。VOI相比早期参考株,应符合:1)病毒表型发生变化,或氨基酸变异引起或潜在引起病毒表型发生变化;2)病毒变异株引起社区传播、或在多个地区或国家传播[4-5]。到目前为止,VOC包括了Alpha、Beta、Gamma以及Delta 4种突变株,SARS-CoV-2已经演变出超过800种的不同亚型或分支[6-7]。新变异株的不断出现,致使疫情不断复燃、蔓延,截至2021年6月27日,全球累计确诊人数超1.8亿,死亡人数近400万[6]。为了控制疫情的发展,世界各国采取了各种防控措施。从防控效果来看,封锁策略一定程度上能有效遏制病毒的传播,但同时长时间的封锁也会给国家的经济发展、公民的心理健康等各方面带来负面影响[8-9]。由此看来,在疫情常态化的今天,接种疫苗才是避免感染COVID-19的根本方法。因此,本文对SARS-CoV-2基因组结构及功能、国际上主要流行的变异株的一些特点、SARS-CoV-2疫苗对变异株保护结果以及应对SARS-CoV-2变异的疫苗策略等方面进行简要概述。
1 SARS-CoV-2的基因组结构及功能
SARS-CoV-2为单股正链RNA病毒,基因组包含约30 000个核苷酸,可编码9 860个氨基酸,主要由2个侧翼非翻译区和整段可编码蛋白的开放阅读框(ORF)组成[10]。其中,5′端的开放阅读框(ORF1a/b)约占整个基因组的三分之二,可在宿主细胞的粗面内质网内被翻译为pp1a和pp1ab蛋白,这2种复制酶多聚蛋白被3C样蛋白酶(3CLpro)和木瓜蛋白酶(PLpro)切割为16种非结构蛋白;3′端的开放阅读框约占整个基因组的三分之一,可编码4种结构蛋白:刺突蛋白(S)、包膜蛋白(E)、膜蛋白(M)和核衣壳蛋白(N)及一些辅助蛋白[11]。
在SARS-CoV-2编码的相关蛋白中,S蛋白是介导病毒入侵宿主细胞的关键蛋白[12]。S蛋白由S1和S2两个亚基组成,可介导病毒识别宿主细胞受体,促进膜融合,并诱导免疫反应产生中和抗体[10]。S1亚基上的受体结合域(RBD)能与靶细胞上的受体特异性结合,从而决定病毒对细胞的趋向性和致病性,因此,RBD也被作为SARS-CoV-2药物和疫苗研发的重要作用靶点。S2亚基上的融合肽(FP)可插入宿主细胞膜表面,在七肽重复序列1(HR1)和2(HR2)之间形成超螺旋结构,介导病毒蛋白发生构象变化,促使病毒与宿主细胞膜融合,从而进入宿主细胞内释放病毒基因组并进行复制[10,13]。
2 SARS-CoV-2主要的突变株
2.1 D614G突变株 D614G突变是指野生型SARS-CoV-2的S蛋白中发生错义突变,原来在614号位编码天冬氨酸(D)的残基突变为甘氨酸(G)。该突变株于2020年1月28日在德国巴伐利亚州的一次小规模疫情中首次被发现,随后在欧洲流行,不断扩散传播到北美洲、大洋洲、南美洲和亚洲[3,14]。但有研究基于GISAID上共享的SARS-CoV-2基因组序列,回顾病毒样本发现1月下旬在中国的广州、四川和上海已经出现这种突变[14]。在2020年3月之前,D614G突变株仅占全球公布病毒株测序序列的不到10%,综述3月,数字增长到了67%,截止到5月底已经上升至78%,到2020年6月,该突变株已经成为全球范围内的主要传播类型[3-4]。假病毒模拟实验分析发现,G614假病毒比D614假病毒更容易进入表达人血管紧张素转换酶2(ACE2)的细胞[15];在体外细胞及动物试验中,D614G突变株对原代人气道上皮细胞的形态及对ACE2转基因小鼠和叙利亚仓鼠的呼吸道组织和肺部疾病的病毒滴度的影响与野生型毒株相似,但该突变株似乎复制能力更强,更具感染性和竞争适应性[16]。 Zhou等也发现D614G突变能增强体外结合和复制,并在体内特别是在传播瓶颈期间更具竞争优势[17]。有研究表明,D614G突变株传播力和感染性的增强似乎与其引起的上呼吸道病毒载量增加、总S蛋白掺入病毒粒子增多、S1脱落减少以及导致更多ACE2结合和融合状态的构象改变有关,但是该突变与疾病严重程度的增加或逃避中和抗体似乎无关[14,18]。在对COVID-19患者的临床数据及SARS-CoV-2基因组序列的研究中也发现,虽然感染含有G614病毒的患者的病毒RNA水平较高,但住院结果并没有差异,D614G突变似乎并不会影响疾病的严重程度[3,19-20]。在突变株的免疫应答方面,Weissman等评估了来自刺突免疫小鼠、非人类灵长类动物和人类的血清对携带D614或G614刺突的假病毒的中和作用。在所有情况下,G614假病毒的中和的敏感性较高,也更容易被RBD结合[21]。Klumpp-Thomas等发现来自高发人群的88名捐赠者的抗体对原始刺突和D614G突变株的刺突均有反应[22]。这些研究均表明了D614G突变并不能逃避中和抗体,对原有疫苗的效果并不会产生重大影响。
2.2 Alpha(VOC 202012/01、 501Y.V1或B.1.1.7)突变株 2020年12月,英国当局首次报告出现一种新型变异毒株——Alpha突变株,该突变株最早一例可识别的临床样本可追溯到2020年9月[6-7]。截至2021年6月27日,该突变株已经传播至172个国家[6]。与野生型SARS-CoV-2相比,Alpha突变株累积了24个突变。其中,S蛋白上有7个错义突变和2个缺失(N501Y、A570D、D614G、P681H、T716I、S982A、D1118H、H69-V70del和Y144del)(图1-A);ORF1ab上有3个错义突变和1个缺失(T1001I、A1708D、I2230T和SGF 3675-3677del);ORF8上有3个错义突变(Q27stop、R52I和Y73C);N蛋白上有2个错义突变(D3L和S235F)[23]。S蛋白上N-末端结构域的H69-V70del与免疫受损患者的免疫逃逸有关,在体外还可增强病毒的感染性,同时,这一缺失还与针对刺突蛋白的ThermoFisher TaqPath探针诊断测试失败有关;P681H替换紧邻Furin裂解点,对病毒的感染和传播可能产生影响;受体结合域的N501Y替换可以增强病毒与ACE2结合的亲和力和细胞感染性[24-25]。这些关键位点的突变可能改变了病毒的感染力、传播力和致病力,影响机体对病毒的免疫应答。有研究表明,Alpha突变株的传播力比之前的毒株高70%[26]。这与Davies等研究推测的Alpha突变株的传播率比英国之前流行的毒株高43%~90%相符[24]。Davies另一项分析英国SARS-CoV-2社区检测阳性和COVID-19死亡相关数据集的研究发现,Alpha突变株相关的死亡风险可能比以前存在的突变株高出61%,该突变株不仅更具传染性,还可能导致更严重的疾病[27]。在Alpha突变株的免疫应答方面,Nuez等对正常未染毒及康复的叙利亚仓鼠(>10月龄)进行了感染和再感染研究,发现自然感染诱导的免疫对叙利亚仓鼠有保护作用[28]。Wang等的假病毒模拟试验发现,康复者血清和Moderna或辉瑞的mRNA疫苗免疫者血清对Alpha毒株中和活性没有显著的影响,预计Alpha毒株对当前疫苗效果不会产生严重影响[29]。Supasa和其同事的研究显示虽然Alpha毒株对阿斯利康疫苗的中和效价降低2.5倍,对辉瑞疫苗的中和效价降低3.3倍,但这2种疫苗依然能保护人体免受Alpha毒株的感染[30]。综上所述,Alpha突变株可能具有更高的传染性和致病性,但并不会逃脱当前疫苗的免疫。
2.3 Beta(501Y.V2或B.1.351)突变株 Beta突变株于2020年10月在南非纳尔逊·曼德拉海湾流行的COVID-19的疫情中首次被发现,随后进行溯源发现2020年8月份该地就已存在该突变株[31]。截至2021年6月27日,已经有120个国家发现存在该突变株[6]。与野生型SARS-CoV-2相比,Beta突变株存在12个错义突变和2个缺失,其中约79%的突变都位于S蛋白上。S蛋白上有9个错义突变和1个缺失(L18F、D80A、D215G、LAL 242-244del、R246I、K417N、E484K、N501Y、D614G和A701V)(图1-B);ORF1a上有1个错义突变(K1655N);ORF1ab上有1个缺失(SGF106-108del);E蛋白上有1个错义突变(P71L);N蛋白上有1个错义突变(T205I)[23-24]。研究表明,Beta突变株具有选择性优势,更具竞争性,可迅速扩散传播至多个地区并成为主流毒株,其传播能力比野生型高约50%,但其是否会影响疾病的严重程度目前还没有相关的证据证明[23,31]。与D614G突变株相比,Beta突变株在携带人类ACE2的细胞中没有表现出更高的感染性,但在携带小鼠ACE2的细胞中感染性增强,表明该突变株有可能感染小鼠,存在跨物种传播的潜能[32]。假病毒模拟实验分析发现,大多数康复者血清对Beta突变株的中和活性降低了2.5倍,多种RBD靶向的单克隆抗体以及多克隆抗体对Beta突变株的中和活性显著下降,这种中和活性的下降与S蛋白RBD上的K417N、E484K以及N501Y的同时突变有关,而RBD外的突变对其影响很小[29,32]。Kuzmina等发现,辉瑞疫苗免疫者血清对Beta假病毒刺突蛋白的中和活性降低了6.8倍,同时该研究也证实了K417N、E484K以及N501Y这3个突变的重要性,这3个突变的组合可导致传染性和中和抗性的增强,还可以提高转导靶细胞的能力,最高可达13倍[33]。此外,Beta突变株对Novavax疫苗、强生单剂疫苗以及阿斯利康疫苗的效果也有影响。Novavax疫苗的有效率从89.3%降至49.4%;强生单剂疫苗的有效率从美国的72%下降到南非的57%;阿斯利康疫苗对Beta突变株的有效率仅为10%[34-35]。以上研究结果显示Beta突变株似乎存在免疫逃逸的可能,但是Edara等研究发现,尽管急性感染和恢复期患者血清及Moderna疫苗免疫者血清对Beta突变株的中和活性有所下降,但其仍能中和Beta突变株,对Beta毒株的感染仍具保护作用[36]。
2.4 Gamma(P.1或B.1.1.28.1)突变株 2021年1月,首次报道在4名从巴西亚马逊到日本东京的游客中发现新的SARS-CoV-2突变株即Gamma突变株,随后有研究表明2020年12月在巴西亚马逊州首府马瑙斯就已检测到该突变株,且基于基因组测序结果推测,2020年11月中旬该地可能就已经存在该突变株[37]。截至2021年6月27日,Gamma突变株已经扩散至72个国家[6]。与野生型SARS-CoV-2相比,Gamma突变株包含17个错义突变,1个缺失和4个同义突变。其中,S蛋白上有12个错义突变(L18F、T20N、P26S、D138Y、R190S、K417T、E484K、N501Y、D614G、H655Y、T1027I、V1176F),是目前S蛋白上累积突变数最多的变异株(图1-C);ORF1ab上有3个错义突变和1个缺失(S1188L、K1795Q、E5665D、SGF 3675-3677del);ORF8上有1个错义突变(E92K);N蛋白上有1个错义突变(P80K)[23,37]。Faria等[37]整合基因组和死亡率数据构建动力学模型发现,Gamma毒株的传播性可能是非Gamma谱系的1.7~2.4倍,这与Coutinho等[38]发现的Gamma突变株的可传播性是以前流行病毒株的2.6倍的这一结论相一致,均表明Gamma突变株的传播性更高。此外,Faria等的研究还发现Gamma突变株出现后感染病毒出现死亡的概率是Gamma出现前的1.2~1.9倍,但由于医疗系统的压力等因素也可导致死亡风险的增加,因此无法确定死亡风险的增加是否是由于感染了Gamma突变株,还需要进一步的研究[37]。该研究还发现Gamma突变株可以规避21%~46%以前感染非Gamma谱系突变株引起的保护性免疫,这表明之前感染过非Gamma谱系的患者可能再次感染Gamma毒株[37]。此外,它还对多种治疗性单克隆抗体无效,对康复者血清和疫苗接种者血清的中和具有相对抗性,其中,康复者血清对Gamma突变株的中和活性下降约3.1倍,疫苗接种者血清对该突变株的中和活性下降了2.2~2.8倍[39-40]。这与Hoffman等的研究结果相似,均表明Gamma突变株可能逃避中和抗体反应,对当前的抗体疗法产生威胁[41]。
2.5 Delta(B.1.617.2)突变株 Delta突变株最早于2020年10月在印度被发现,截至2021年6月27日,已经有96个国家明确发现存在该突变株[6]。该突变株S蛋白的RBD上含有T478K和L452R双重突变,Furin裂解位点上含有P681R替换(图1),研究表明这些突变可能会增加传染性,增加S1-S2裂解速率以及降低抗体介导的中和作用,并可能具有免疫逃逸能力[42]。世界卫生组织对GISAID上提交的该突变株的序列进行初步分析也发现,Delta突变株的增长率明显高于印度其他流行的突变株[43]。日本的一项研究也表明Delta毒株的传播率是Alpha毒株的1.23倍,该毒株不仅传播至多个国家还迅速成为当地的主流毒株[44]。Chaudhari 等通过计算分析、蛋白质建模和分子动力学模拟方法对Delta突变株的基因组和结构基础进行探索,发现该突变株具有向更高传播性和致病性进化的可能[45]。Arora等的研究发现Delta突变株具有更强的进入和融合肺细胞的能力,可能有助于增加其传播性和致病性,而且该突变株对用于COVID-19治疗紧急使用的单克隆抗体Bamlanivimab具有抗性[42],这与Planas等[46]的发现相一致。此外,Planas等还发现,恢复期患者血清和辉瑞Comirnaty疫苗接种者血清对Delta突变株的中和活性比Alpha突变株低3~6倍,接受一剂阿斯利康疫苗的个体的血清几乎不抑制 Delta突变株,该突变株似乎具有免疫逃逸的可能[46]。Yadav等评估了Delta突变株与恢复期患者血清及接种两剂BBV152疫苗者血清的中和活性,发现与D614G突变株相比,Delta突变株对恢复期患者血清的中和活性降低4.6倍,对接种两剂BBV152疫苗者血清的中和活性降低2.7倍,但该疫苗对Delta突变株的感染仍具保护作用[47]。
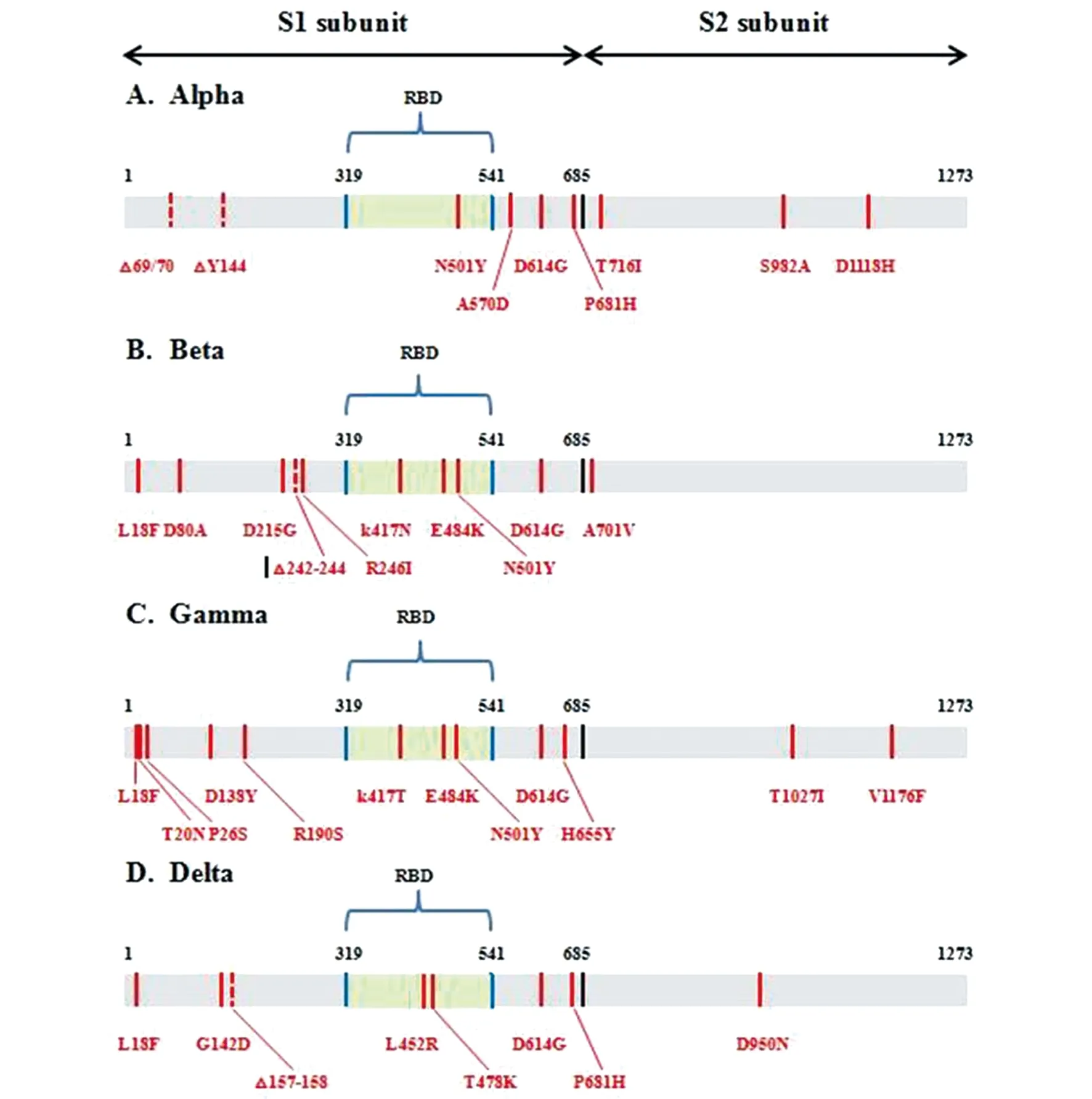
A为Alpha突变株S蛋白中氨基酸替换示意图;B为Beta突变株S蛋白中氨基酸替换示意图;C为Gamma突变株S蛋白中氨基酸替换示意图;D为Delta突变株S蛋白中氨基酸替换示意图。
2.6 其它SARS-CoV-2突变株 Epsilon(B.1.427/B.1.429)突变株最早于2020年5月左右出现, 直至12月才在美国首次被检测报道,其S蛋白上包含3个错义突变(S13I、W152C 和 L452R),ORF1ab上有1个错义突变(D1183Y)[48]。研究显示这2种突变株的传播性比野生型SARS-CoV-2增加了18.6%~24.0%,与恢复期患者血清和疫苗接种后血清结合的中和效价分别降低了4.0~6.7倍和2.0倍,S蛋白RBD上的L452R突变减少了与RBD 特异性单克隆抗体 (mAbs)的中和活性[48-49]。Iota(B.1.526)突变株最早于2020年11月在美国出现[6]。 S蛋白上包含6个错义突变( D614G、L5F、T95I、D253G 、 A701V以及 E484K 或 S477N),其中含S477N的Iota突变株可以被恢复期患者及疫苗接种者的血清完全中和,含E484K的Iota突变株中和活性降低3.5倍[50]。Annavajhala等的研究显示Iota毒株导致再感染或逃脱疫苗免疫的风险可能低于Beta,但高于Gamma突变株[51]。Zhou等的研究显示目前的疫苗和治疗性单克隆抗体对Iota突变株仍具保护作用[52]。B.1.616突变株最早于2021年1月在法国出现[6]。 其S蛋白上含有9个错义突变(H66D、G142V、D215G、V483A、D614G、H655Y、G669S、Q949R和N1187D)和1个缺失(Y144del),研究发现该突变株可能逃避鼻咽拭子核酸检测(RT-PCR检测),导致难以及时诊断和甄别,对于其能逃避传统检测的原因,可能是由于病毒载量低于上呼吸道检测极限,而不是常规RT-PCR靶点与此突变株之间的基因组不匹配,研究还显示该突变株似乎具有较高的致死率[53]。
3 SARS-CoV-2疫苗
3.1 SARS-CoV-2疫苗对变异株的保护结果 当前,世界卫生组织紧急使用清单主要包括2种腺病毒载体疫苗(阿斯利康疫苗和强生疫苗),2种mRNA 疫苗(Moderna疫苗和辉瑞疫苗)以及2种灭活疫苗(中国国药北京生物和科兴疫苗)[54]。体外试验结果显示,疫苗接种者的血清对突变株的中和活性均有不同程度的下降,在临床试验以及真实世界中的数据也表明疫苗对突变株的保护力有所下降[55]。其中,阿斯利康、Moderna和辉瑞疫苗对Alpha突变株的保护力分别为74.5%、100%和93.7%[56-57];阿斯利康、强生、Moderna和辉瑞疫苗对Beta突变株的保护力分别为10.4%、57%、96.4%和75.0%[35,55,57-58];科兴疫苗对Gamma突变株的保护力估计为56.2%[59];阿斯利康和辉瑞疫苗对Delta突变株的保护力分别为67.0%和87.9%[56]。相关研究显示疫苗对突变株的保护效果大体上为:mRNA疫苗>腺病毒载体疫苗>灭活疫苗,这与以上研究的结果相一致[60];而对于疫苗接种后的不良反应发生率,研究表明腺病毒载体疫苗>mRNA疫苗>灭活疫苗[61]。
3.2 应对SARS-CoV-2变异的疫苗策略 首先,应该建立类似全球流感监测的监测SARS-CoV-2变异的网络系统,以及时监测SARS-CoV-2的变异以及变异对现有疫苗效果的影响,根据现有疫苗对突变株的保护效果,可以考虑通过增加接种剂次以及不同疫苗混合接种来提高疫苗的免疫原性,但需要进一步的研究以确保混合接种疫苗有效性及安全性[55,62];其次,针对新的变异株应该要能够快速实现疫苗的更新迭代;最后,研发通用型的冠状病毒广谱疫苗将是控制SARS-CoV-2突变株传播的有效方法。有研究发现不同属冠状病毒融合肽(FP)疫苗具有相似的保护作用,提示FP可能是开发广谱冠状病毒疫苗的有效靶点,还需要进一步研究证实[63]。
4 结 语
SARS-CoV-2仍在全球传播,由于选择压力和病毒在人群中的不断复制,新的变异也将继续出现[64]。目前已经有研究报道存在接种疫苗后的突破性感染,国内广东境外输入Delta毒株的传播链中也存在少数的突破性感染,但感染者的疾病严重程度明显减轻,这也提示现有疫苗对突变株仍具有保护作用[65]。在针对变异株的新疫苗没有出台前,普及接种现有疫苗,提前建立好群体免疫屏障便很重要。Lancet一项研究显示,在非自然感染情况下,达到群体免疫需要将近100%的人群接种1剂疫苗或80%的人群接种2剂疫苗[66]。因此,现阶段在控制疫情传播扩散的同时世界卫生组织也在强烈倡导普及疫苗接种,世界各国也都正在积极进行中,加速扩大疫苗覆盖率,建立群体免疫屏障,争取在出现超级变异病毒前将病毒压制住,将其转变为“常态化”病毒。
利益冲突:无
引用本文格式:蒋潘虹,阮嘉雯,俞慕华,等. 新型冠状病毒(SARS-CoV-2)变异的研究进展[J]. 中国人兽共患病学报,2022,38(2):157-164. DOI:10.3969/j.issn.1002-2694.2021.00.030
猜你喜欢
中学化学(2024年2期)2024-06-17 04:01:47
广东药科大学学报(2023年5期)2023-12-30 00:08:39
时代报告·奔流(2022年1期)2022-04-29 04:10:56
昆明医科大学学报(2022年3期)2022-04-19 13:59:42
科学大观园(2022年2期)2022-01-23 11:05:15
中国兽药杂志(2021年7期)2021-08-13 02:37:22
艺术品鉴(2020年5期)2020-07-27 02:43:08
丹青少年(2017年2期)2017-02-26 09:10:51
动物医学进展(2015年10期)2015-12-07 05:46:18
浙江医学(2014年17期)2014-04-13 10:13:16
