
槲寄生植物(Mistletoe):森林中的重要资源
2022-03-17 05:30黄大明王思玥朴宙元DavidWatson
中国科技纵横 2022年2期
黄大明 王思玥 朴宙元 David M.Watson
(1.清华大学生命学院生态学实验室,北京 100084;2.澳大利亚查尔斯特大学约翰斯通中心和环境研究组,新南威尔士州巴瑟斯特 2795)
0.引言
很久以前人们就关注到槲寄生(Mistletoe)植物与动物之间的关系,达尔文把它作为进化适应的典范[34]。Linne首先描述了槲寄生植物的生活史,注意到画眉鸟吃槲寄生果实后会将黏黏的种子吐在后来的栖木上[112]。Pliny描述了发生在1600年前类似的观察结果[160]。实际上,大多数槲寄生植物都依靠动物(主要是鸟类)传播种子[37,83,105,173,177,209],这种密切关系是典型的种子传播模式[89,92,93,127,207,242]。在一些区域,有研究鸟类利用槲寄生植物的记录:印度[49,51]、北美[84,220]、澳大利亚[173,330]、拉丁美洲[183,195,202];和研究食果过程[177,209]。达尔文曾指出:槲寄生植物也通过动物传粉(主要是鸟类和昆虫),但关于其生活史的研究较少[34,51,67,159,173,244]。实际上,大多数槲寄生植物与动物相互作用的研究都是附带的。已经进行系统整理的类似工作如:地衣[198]和棕榈树[248]。
本文主要研究脊椎动物与槲寄生植物的关系。尽管我们知道许多昆虫为槲寄生植物传粉(主要为鞘翅目、双翅目、膜翅目和鳞翅目)[85,244]。据记载,有200多种昆虫为一种槲寄生植物传粉[159]。鞘翅目、双翅目、半翅目、同翅目、膜翅目、鳞翅目、直翅目和缨翅目内的许多昆虫都以槲寄生为食,所有目都含有槲寄生植物的寄生种类。除了一些顺带研究,专门研究槲寄生—昆虫关系的工作很少,并且没有比较综合性研究[55,85,132,244]。
1.槲寄生植物概述
1.1 组成和分布
槲寄生是多类有花植物有1300多种,分布在除南极洲以外的所有大陆[37,38,105,138,240]。专营半寄生:所有水和矿物质都是通过特殊的寄生根从宿主获得[59,110,111,156]。它们分别属于檀香目(Santales)的5个科[104,105,141],且气生形式至少有4~5个分支(见图1)[138,139,140]。因此,槲寄生(Mistletoe)不是指植物分类群,而是表示功能群(类似红树林),代表檀香目中所有半寄生植物。其中2个科:Misodendronaceae和Eremolepidaceae鲜为人知,仅分布在新热带森林,而檀香科(Santalaceae)的几个气生属则分布于从拉丁美洲到东南亚的热带雨林中[104,105,240]。相比之下,桑寄生科(Loranthaceae)和槲寄生科(Viscaceae)世界范围分布,研究较多,大约分别包括940和350种,包括绝大多数槲寄生植物(>98%)[138,240]。这些科不是姐妹分类群,其气生形式是独立进化的(见图1)[37,105,138,139]。根据白垩纪的化石,以及澳大利亚、新西兰和南美洲发现的亲缘属[16,105],认为:桑寄生科最先出现在冈瓦纳大陆,后来传播到非洲、欧洲和北美。而槲寄生科则起源于东亚,在第三纪早期通过劳亚大陆辐射,二次传播到南部大陆[16,161,170]。
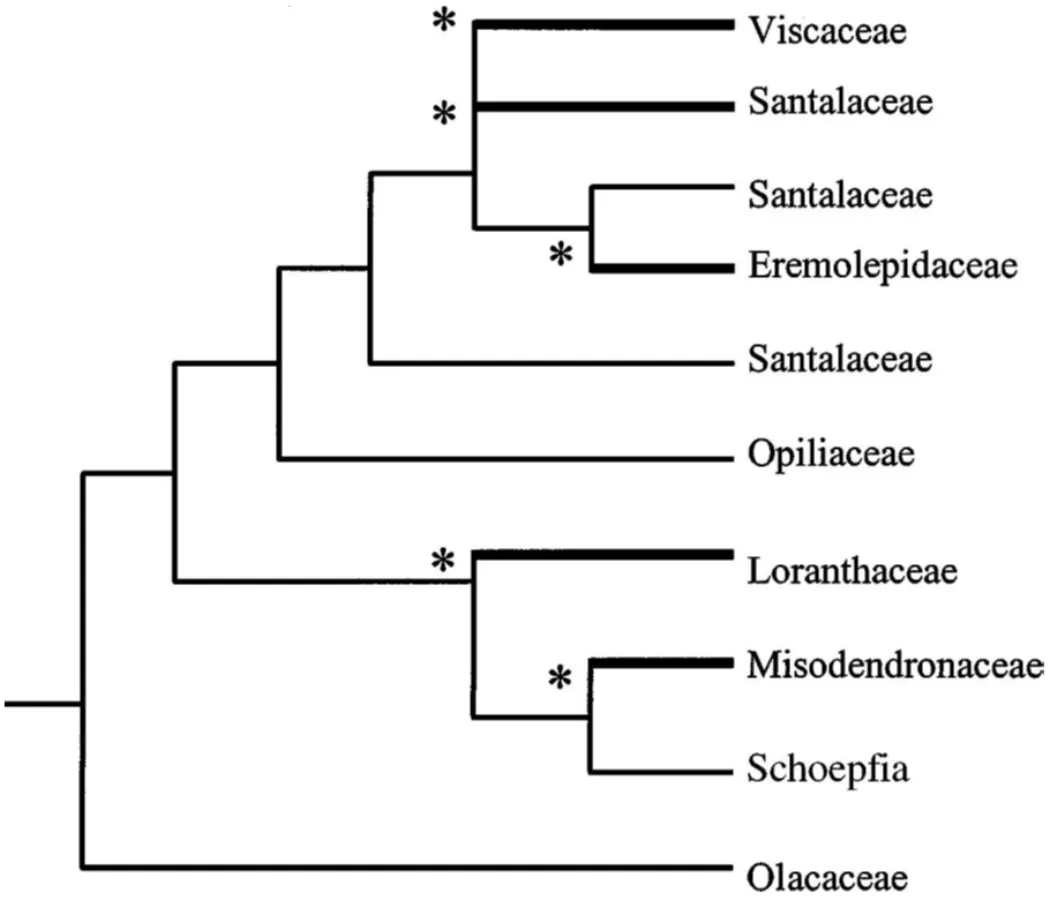
图1 基于来自核18s rDNA和叶绿体rbcL序列的檀香目分支树粗线的分支表示槲寄生植物类群,星号表示空中半寄生形式的独立进化起源。[138]
目前,桑寄生科和槲寄生科广泛分布于欧洲、美洲、非洲、亚洲和澳大利亚大陆(塔斯马尼亚岛除外),从北方气候带到温带,热带和干旱地区,仅在极端干燥或寒冷地区不存在[16,105,170]。在海洋岛屿上也有很广泛的分布,亚速尔群岛、马达加斯加、阿尔达布拉、科摩罗群岛、马斯克林群岛、加拉巴哥群岛、夏威夷群岛、新喀里多尼亚、豪勋爵岛、诺福克岛、新西兰、斐济群岛、亨德森岛,大、小安的列斯群岛和伊斯帕尼奥拉等都具有代表性的地方种类[16,85,105,161]。
虽然槲寄生通常被认为是森林植物,但拥有广泛的宿主:北方森林中的针叶树[83,85]、非洲和拉丁美洲沙漠中的肉质植物和仙人掌[124,161],澳大利亚沿海荒地的草本植物和一年生禾本植物[64],甚至是中美洲云雾林中的兰花和蕨类植物[106]。然而,大多数槲寄生植物寄生于树木和灌木,其多样性因地而异[37,83,105]。
1.2 生活史与物候学
除了几种根寄生外,槲寄生植物的特点是它们的生境[64,105]。它们通常在宿主树冠上长成密丛[见图2(a)和(c)],这些密丛通常由半肉质的槲寄生茎和叶组成[见图2(a)和(b)]。矮小的槲寄生(Viscaceae,Arceuthobium spp.)密丛(称为a witch's broom,VA矮小密丛)表示宿主对寄生感染的反应:由加厚和扭曲的枝条组成[见图2(c)和(d)][85,197]。通常认为槲寄生植物危害树木健康[43,83,86,180],但并不一定如此[178,179,216]。槲寄生植物的年生存率很低[133],2种澳洲槲寄生植物年生存率估计分别为19%和31%[178],被认为是“低级K选择森林寄生物”[8]。许多槲寄生植物对火[85,101,193]和霜[95,206]敏感。由此限制了槲寄生植物在一些区域的分布[82,85,214]。槲寄生植物种子需要高水平光照才萌芽、定植和生长成熟[103,108,109,161],但随着宿主冠层的发育,经常被遮蔽[108,186]。因此,尽管传播成功率和发芽率很高,但定植的很少,目前研究的所有槲寄生植物的微生境都很狭小[85,103,108,109,151,195,196,247]。

图2 槲寄生植物
槲寄生通过寄生根从宿主获得所有的水、矿物质和营养[37,163]:见图2(b)。肿胀的固着部分既可以将槲寄生植物固着在宿主上,又可以将水和矿物质等转移到寄生植物中。使用半寄生物一词是因为大多数槲寄生植物尽管可以从宿主获得高达60%的碳水化合物,但还能进行光合作用[96,110]。这种生长形式的结果使槲寄生植物很少受到土壤、降水和营养因素的影响,宿主植物缓冲了生境资源大规模波动对槲寄生植物的影响[59]。
在哥伦比亚雨季云雾林中,5种槲寄生植物具有连续的结果期和广阔的峰值,花期具有同样的连续性,全年都有花蜜和果实。开花和结果期10~12个月不等,具有明显的潮湿季高峰[62,113,183,195,202,217]。非洲南部的槲寄生植物也有较长的结果期,从3个月到全年不等[72,161]。而Burkhina Faso的槲寄生植物全年表现出交错的结果模式[31]。在Palearctic的矮小槲寄生植物开花和结果季节非常有限,持续4~6个月:开花通常在初秋,中秋达到顶峰,在秋末到初冬结果率最高[85]。印度南部6种槲寄生植物的物候,开花期为3~9个月,结果期为4~7个月。其果实和花蜜的丰盛度都呈资源分割状,因此任何时候都有至少2种的花蜜和果实[50]。13种澳大利亚桑寄生科的植物:几种是全年花期,其他不同模式与区域气候有关。在潮湿地区,明显的夏季高峰值与较干旱冬季低峰值形成梯度,此时开花较少,果期全年。一个区域内至少有一种槲寄生植物在花果期[173]。
1.3 果实散布和授粉
几种槲寄生植物使用液压爆方式扩散种子(特别是Viscaceae科),接着是鸟类进行远距离传播[84]。一般认为槲寄生种子是鸟类传播[37,83,105,173]。对新世界槲寄生植物研究表明:脊椎动物的种子传播在槲寄生植物多样性上起着关键作用[182]。由脊椎动物传播种子的槲寄生植物具有更高的多样性。大多数进行槲寄生植物果实种子扩散的鸟类是几种高度特化种类[175,176,195,208,235]。这些特化的生理,解剖和行为等特征相互协同适应[72,73,116,174,175,177,182,187,208,209,234]。许多槲寄生植物果实也表现出一系列协同性状:大、甜、成熟时颜色明显,有小花梗或直接长在枝上。果肉组成不同,但大多数具有高比例的可溶性碳水化合物,通常干重为40%至60%[73,110,183],有的高达74%[209]。一些loranthaceous种类的果实含有高脂质(干物质含量高达35%)[73,183],Loranthus europaeus果实含有纯脂肪滴[41]。粘性果实种类往往比其他果实具有更高的蛋白质含量[243]:3种Phoradendron的平均含量为22%[183]。含有多达18种游离氨基酸[41,73]。槲寄生果实的详细生化分析已确定了10种必需氨基酸,尤其是精氨酸,赖氨酸和苯丙氨酸[36,41,73,191,246]。与其他槲寄生组织一样,果实还含有高浓度的矿物质(特别是P和K)和微量元素(如Mn和Fe)[110,156]。据说槲寄生植物有毒,但仅在Viscaceae中的少数种类中分离出毒素,且毒性很小[18,36]。许多槲寄生植物果实不连续成熟,单株植物在整个结果期都有成熟的果实,不同植物不同种类高峰期不同[173,183]。这些种内种间的适应辐射,使一些区域全年都有果实和花[31,50,161,173]。
尽管大多数槲寄生种类都是鸟类传粉,但槲寄生科的种类主要通过风和昆虫传粉[85,105,161,244]。确定具体喜风或喜昆虫传粉的属性,还很不容易[85,159]。桑寄生科主要由鸟类传粉[37,51,105,107,173,189],具有与鸟类传粉相关的特化性状:大无味花,通常颜色鲜艳(黄色、橙色、红色),具有坚固的花冠,短花柄,并且通常有大花序(图2b)[173,240]。通常花蜜等分泌物丰富,且富含糖:葡萄糖、蔗糖,尤其是果糖(总糖含量高达60%[13,173,218])。与果实扩散不同,目前没有鸟类,昆虫或其他物种完全依赖槲寄生花蜜作为主要食物来源[50,51,173]。更确切地说,各种各样的物种为槲寄生植物传粉,其中一些可能只在特定季节依赖于花蜜[51,62,67,107,189,217,237]。
2.槲寄生植物与脊椎动物的关系
2.1 槲寄生作为食物来源
一些鸟类和哺乳类以槲寄生植物为食,包括采食其花蜜、花、果实、种子或叶。以科为单位列于表1。
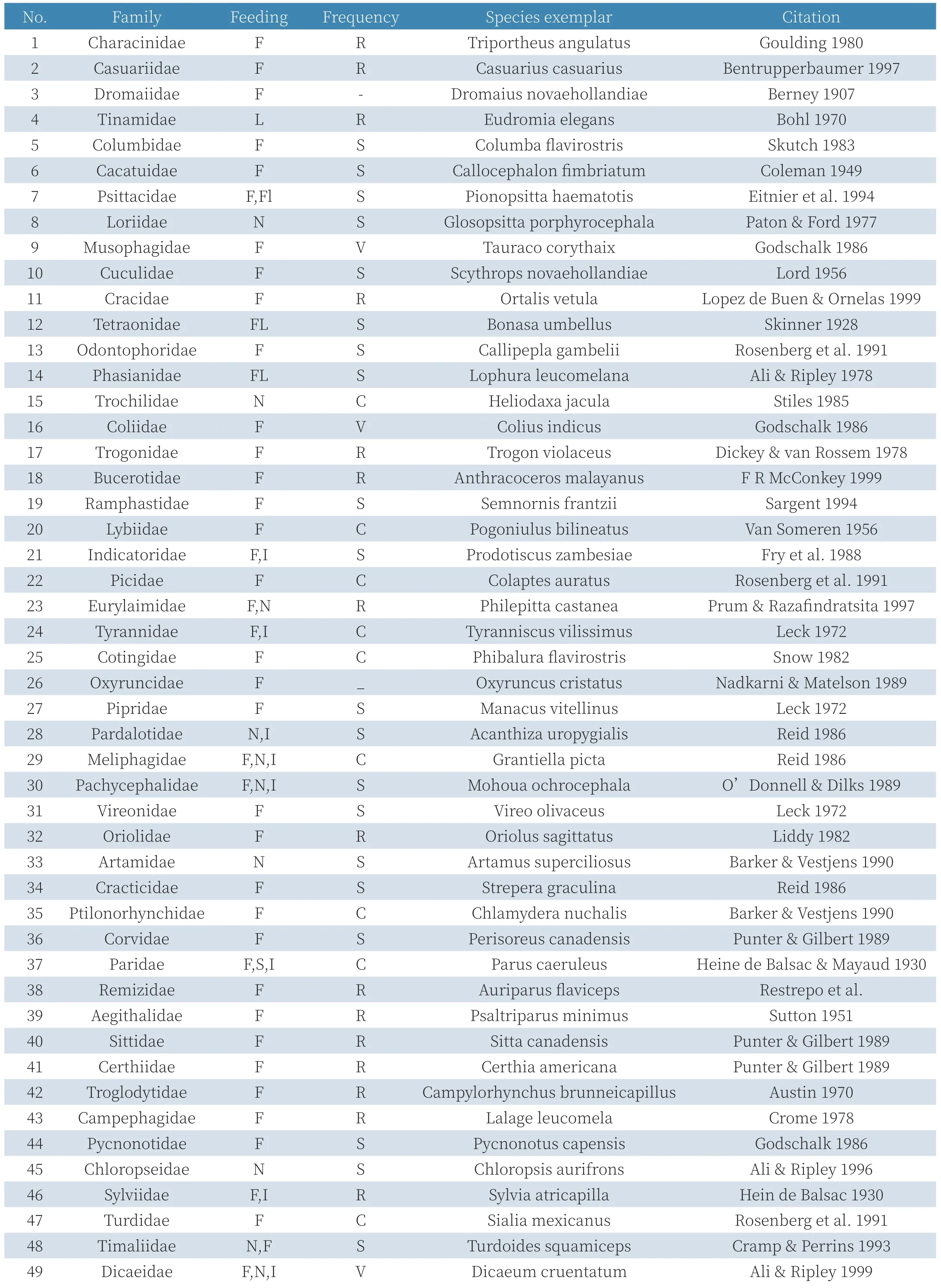
表1 以槲寄生植物为食的科

(在营养模式栏中,F果实,Fl花,N花蜜,S种子,W全株及槲寄生的昆虫。在频率方面,R表示科中罕见,S表示几个记录,C表示常见科,V是非常常见的——并且是针对单型科。)
有66个鸟类和30个哺乳类科利用槲寄生植物,分属12和10个目。还有在亚马逊洪水高发季节以槲寄生植物为食的特殊鱼类(Triportheus angulatus)[76]。鸟类一般采食槲寄生植物的果实和花蜜,而大多数哺乳类采食果和叶。一般认为只有鸟类专吃槲寄生植物果实,但在南安第斯云雾林中,古代有袋动物目Microbiotheria中的一种负鼠Dromiciops australis是槲寄生植物果实的专性扩散者。二者起源于冈瓦纳大陆,在早期就存在关联[5]。虽然许多爬行动物是果食性或叶食性,但食性分析中没有发现采食槲寄生植物[168]。
许多区域全年都有槲寄生植物的果实,很多物种都会来采食,但槲寄生植物的种类并不多,这些消费者大多采食果实,可能偶尔起到传播作用。这种随机性是不可预测的,出人意料的是槲寄生植物具有高度特异的扩散系统,在进化过程中逃避广食果动物[89,127,129,177,242]。许多槲寄生植物果实大都具有大而粘、相对隐形的特点,以逃避泛食果动物,有利于特化种类[127,242],许多区域的槲寄生植物依靠少数特化食果鸟类完成种子扩散[32,72,74,87,116,165,173,175,176,183,195,208,239]。
槲寄生植物还有丰富的花蜜,来觅食的不仅有许多食蜜种类[50,107,173,207,217],还有肉食性昆虫和广食性种类[14,15,47,147,173,192],和几种哺乳动物(常见的是吃整花)[58,88,98,125,150]。没有哺乳动物传粉的记录。
许多食叶和浏食动物非常喜欢采食槲寄生植物的叶子,从鹿和犀牛到大猩猩和负鼠(见表1)。一般认为从澳洲引进的刷尾负鼠(Trichosaurus vulpeca)摧毁了新西兰当地槲寄生植物的数量[143,148],当然还有传粉鸟类种类减少的因素[189],实际上,食叶哺乳动物是槲寄生植物叶子宿主拟态的自然选择压力[18,42,99,155,245],没有哺乳动物专门采食槲寄生植物,但有几种表现出季节性依赖[166,188],有几种以槲寄生植物叶为食的鸟类,但类群很少[2,30,200,224,131]。
槲寄生密丛常被食虫类作为觅食场所。如前所述,密丛中有大量与槲寄生植物有关的如传粉昆虫、植食昆虫群落[21,55,132,244],几个高度特化的食虫鸟类利用槲寄生密丛进行觅食活动(如,Prodotiscus regulus,Arachnothera longirostris)[4,21,69,87,147,230]。然而,这些食虫动物基本不食槲寄生植物,这类物种超过10个科。还有一种捕食性蜥蜴(Varanus albigularis)利用槲寄生密丛作为猎鸟场所[190]。突显了槲寄生密丛栖息地的重要性和复杂性。
2.2 以槲寄生植物营巢与栖息
槲寄生植物被广泛用作营巢和栖息场所[27,33,39,61,69,120,142,154,167,184,201,228,231,241]。相关的鸟类与哺乳动物科列于表2。
有43个鸟类和7个哺乳动物科利用槲寄生植物营巢或栖息(见表2),没有专性类群,但有几种鸟类喜欢在槲寄生密丛中营巢(特别是Coccyzinae,Musophagidae,Striginae,Accipitrinae,Ptilogonatidae和 Ptilonorhynchidae)[27,69,142,154]。尚不清楚其选择因素[29,97,126,192,223,247]。例如,长耳鸮(Asio otus)使VA矮小密丛为巢的主要结构支撑,一项研究发现20个巢中有19个与槲寄生植物相关[33]。有许多雀形目的巢较小,隐藏在多叶槲寄生密丛中[6,23,29,32,126,142]。以槲寄生植物营巢除了结构因素外,还有微气候因素。如豪猪(Erethizon epixanthum),松貂(Martes americana)和几种松鼠会在密丛中休眠[120,153,154,204,225]。在炎热的天气里,槲寄生密丛是几种鸟类和哺乳动物隐蔽场所[32,97,220],此外,非洲和北美的鸟类用新鲜槲寄生枝条作巢衬。虽然鸟类使用绿叶作巢衬的行为很普遍,但至少有7种猛禽选择槲寄生科植物作巢衬,经常每天更换[32,69,154,231]。槲寄生植物(Viscum spp.)提取物具有抗菌活性[77],临床试验发现其中一些化学物质可以诱发免疫反应[68,181,219,233]。紫翅椋鸟(Sturnus vulgaris)的实验表明:巢中有各种芳香草本植物时,雏鸟具有较高的免疫功能[79],槲寄生科植物在巢中可能与清洁与抗感染,改善微气候有关[21,23,85],槲寄生植物很多生长在大尺度的林缘[169],直接或间接成为脊椎动物的营巢和栖息场所,槲寄生植物丰盛度与营巢鸟类的多样性和丰盛度有密切关系[21-23,84,120,153]。
3.槲寄生植物的群落特征
关键种概念已经应用于各种群落群,包括捕食者、猎物、寄生蜂、附生物、过渡群落和优势种群落[152]。一般认为关健种对群落或生态系统的影响较大,其丰盛度却很低[162],许多学者认为槲寄生植物是森林的关健种或重要资源[9,70,128,135,172,179,180,199,216,227],有2个研究较多的群落。
3.1 美国西南部的豆科灌丛
代表区域是以豆科类(Prosopis spp.),金合欢类(Acacia spp.)和石炭酸类(Larrea tridentata)等灌木和小树为主,与河岸带和高地之间形成的半干旱河谷[7,28,97,192,215]。这里有槲寄生科的两个属:Arceuthobium和Phoradendron,前者只寄生在针叶树上,很少在豆科灌丛上出现。Phoradendron寄生在很多树上,其中P.californicum和P.tomentosum在豆科灌木上最常见,昆虫(主要是双翅目和膜翅目)是主要的传粉者[244]。鸟类主要扩散果实[171,192,214]。丝鹟属鸟类(Phainopepla nitens)主要以Phoradendron浆果为食[234],其他几种鸟类(西部蓝鸟Sialia mexicanus,雪松太平鸟Bombycilla cedrorum,小嘲鸫Mimus polyglottos和几种啄木鸟)根据果实丰盛度部分采食[171,192,223]。虽然这些种类是果实的主要传播者,但许多其他食虫和泛食鸟类也采食这些果实。冬季很少有其他果实资源可利用[7,12,185],因此一些鸟类与槲寄生植物协同适应[235],在夏季,槲寄生植物果实会被当作水源[97,234]。几种哺乳动物采食果实和叶子(例如,美洲叉角羚Antilocapra americana[194],北美黑尾鹿Oedocoileus hemionus-Riney[188],大盆地小囊鼠perognathus parvus[35])。槲寄生植物还是土著人的食物和药品[48,229],一些哺乳动物季节性依赖于槲寄生植物:在结果高峰期,中美蓬尾浣熊(Bassariscus astutus)的粪便中有很多Phoradendron种子[226]。P.tomentosum叶子可以占白尾鹿(Oedocoileus virginianus)冬季采食量的65%[166]。Phoradendron密丛被许多物种用于营巢[29,32,171,241],并被认为是几个物种的优选营巢地。在研究亚利桑那州的phainopeplas时,80%在豆科植物中营巢穴,其中80%是在槲寄生密丛中或之下[171]。18% 在仙人掌(Campylorhynchus brunneicappilus)中槲寄生P.californicum密丛营巢[6]。红腹唧鹀(Pipilo aberti),50%的巢位于槲寄生密丛(P.californicum)[63]。此外,几种猛禽通常在槲寄生密丛中营巢,还有动物使用新鲜的Phoradendron作巢衬,包括一些啮齿动物[32,220],还有研究指出槲寄生植物P.californicum与几种鸟类的丰盛度有关[12],是15/20种鸟类的重要栖息场所[7,185]。P.californicum浆果是该地区唯一可靠的果实来源,它决定了整个豆科灌从中果实的分布,冬季有以其浆果为主的11种重要食物[192],此群落Phoradendron植物密度差异很大:每公顷1至17棵植物[28];0.5%至8.7%植物有槲寄生[215]。
3.2 澳大利亚东南部的桉树林
常绿硬叶的桉树林遍布澳洲东部,延绵到内陆其他林地和稀树草原,冠层以桉树(Eucalyptus spp.)为主,小树和灌木(如:Acacia spp.Leptosoermum spp.Casuarina spp.)一般会形成亚冠层,在与沟谷和干山坡过渡处会有其他群落[205,228,230]。此地有20多种桑寄生科植物,其中7种广泛寄生在桉树上[Amyema bifurcatum,A.miquelii,A.pendulum,Dendrophthoe glabrescens,D.vitellina,Diplatia grandibractea,和Muellerina eucalyptoides][57]。都营鸟类传粉[67,173],食蜜昆虫也有一定传粉作用(主要在Meliphagidae)[157,173],很多鸟类会随机采食槲寄生花[14,15,67,173,230],特别在夏季森林中花蜜量最低的时候[157]。鸟类是唯一的果实传播者,林子里还有2种专门利用槲寄生植物的特化鸟(槲寄生鸟Dicaeumhirundaceum和画食蜜鸟Grantiella picta),还有许多其他物种采食槲寄生果实[14,15,173]。槲寄生是林中为数不多的可利用果源之一[173],几种哺乳动物经常采食槲寄生的叶和花[40,42,99,173],抑制一些区域槲寄生的丰盛度。槲寄生密丛还是许多食虫类的捕食场所,可以指示昆虫密度[230]。槲寄生密丛明显比桉树叶片密集[见图2(a)],许多物种用其营巢[27,39,142,228],虽然大多是随机的,但有些物种喜欢在槲寄生密丛中营巢[66]。新南威尔士州北部28%的叽喳采蜜鸟和29%的红食蜜鸟在槲寄生密丛中营巢。奇卷尾袋貂(Pseudocheirus peregrinus)几乎完全生活在槲寄生密丛中[116,205,228,230,237],一般在退化和多干扰的区域,长槲寄生的桉树密度范围从每公顷不到3株到超过100株。而纯自然林地,通常少于每公顷10株[86,146,206]。
4.结语
在森林中,一般槲寄生植物的生物量和丰盛度都非常小,但能提供优质营养资源,支持几种专性和许多广食性物种,特别是在资源稀缺季节,为许多物种提供营巢、栖息和捕食场所[162]。在以上2个群落中,一个是半干旱,以灌木为主的群落,其中粘性槲寄生主要是昆虫传粉。另一种是温带中等森林,主要由鸟类传粉,矮小的槲寄生密度是影响鸟类丰盛度的重要因素,28种鸟类中24种的丰盛度受槲寄生密丛丰盛度的影响[21-23,84]。槲寄生植物与脊椎动物关系的研究有许多:南部非洲大草原的槲寄生植物与脊椎动物[54,69,72-74,231],澳洲中部的金合欢灌丛[14,15,27,142,175,176],印度的热带森林[3,4,49,50,51,132],智利以仙人掌为主的沙漠[123,124],哥斯达黎加的亚热带云雾林[62,195,196,201-3,217]。
有研究表明:脊椎动物丰盛度与槲寄生植物密度正相关。在澳洲南部林地中,29种常见林地鸟类的12种分布模式与槲寄生植物的密度有关。随着槲寄生植物密度的增加,鸟类的丰盛度也增加,直至达到每公顷约20棵树的阈值,但不是简单的线性关系。高密度林斑(每公顷多达93棵树被寄生)一般都严重过牧,有状态不佳的鸟类群体[86,144-146,236,238],有4种槲寄生植物的林斑中鸟类的平均丰盛度比只有1种的林斑高63%[236]。鸟类与果实性槲寄生植物协同演化,促进附近区域的α多样性[208,235]。南部非洲的槲寄生分布与土壤类型密切相关,槲寄生在较肥沃的土壤类型更为常见[54]。影响澳洲与新西兰槲寄生分布的因素非常复杂,包括环境、宿主,传粉者、果实扩散者等的综合作用[144-145]。随着欧洲移民定居点的增多,槲寄生在澳洲东部变得越来越普遍[86,180],而在新西兰则变得越来越少[143,148],在西澳洲的林斑中或缺或剩[144,145]。
移除槲寄生植物对群落的影响:
(1)专性食槲寄生果实和叶的动物丰盛度降低,区域种群数量下降到局部灭绝。
(2)特化槲寄生采食者(食果,食叶和食蜜动物)的丰盛度降低。
(3)长期效果会减少枯枝、树洞和树疤等。
(4)降低在槲寄生密丛和树洞中营巢的鸟类和哺乳动物的丰盛度。
(5)一般会降低脊椎动物的丰盛度。
(6)群落对干旱和其他干扰的敏感度增加。
(7)斑块状森林生境的槲寄生密度通常异常高或低。此方法可以用于商业种植园。澳洲东部商业林资料表明:槲寄生植物养殖和林业可以共存,该主题值得进一步探讨[7,23,86,144,145,179,180,189,216,230]。
人们普遍认为槲寄生是破坏性的有害生物。许多土地所有者、管理者甚至生物学家都认为槲寄生是入侵的有害生物,对个体树木造成破坏,危害森林健康,真正理解槲寄生植物在生态系统中的地位与功能不是件容易事。本研究认为槲寄生植物是全世界许多森林和灌丛的重要种类,是重要的森林资源。尽管槲寄生植物占群落多样性不到1%,对群落多样性的影响难以确定,但作为营养源、营巢地、栖息地、隐蔽场所、捕食场所、营巢材料、特殊生境等,在2个新热带森林群落资源匮乏季节支撑了大多数食果脊椎动物[6,21,114,145,173,208,230,227]。
猜你喜欢
中国科学探险(2022年8期)2022-12-31
疯狂英语·新悦读(2022年12期)2022-03-08
东坡赤壁诗词(2022年1期)2022-02-25
中国民间疗法(2021年13期)2021-08-30
新疆大学学报(自然科学版)(中英文)(2020年2期)2020-07-25
林业调查规划(2020年3期)2020-06-08
语文教学与研究(2014年18期)2014-11-07
语文教学与研究(读写天地)(2014年6期)2014-07-09
知识窗(2011年11期)2011-05-14
生物进化(2010年3期)2010-09-27
