
外源钙缓解花生低温光合障碍的调控机制
2022-03-16 08:36刘欣悦刘轶飞易伯涛孙志宇张思威马明珠韩晓日
植物营养与肥料学报 2022年2期
刘欣悦,刘轶飞,易伯涛,孙志宇,张思威,马明珠,韩晓日
(土肥高效利用国家工程研究中心/沈阳农业大学土地与环境学院,辽宁沈阳 110161)
起源于南美洲热带地区的喜温作物—花生(Arachis hypogaea)是世界重要的油料和经济作物,也是重要的食用蛋白源和食用植物油源,在保障我国食用油安全方面具有举足轻重的作用[1]。特别是,基于后疫情时代“双循环”新发展格局下、中国大豆产需缺口加大等背景下,中央明确了油料作物结构调整主要策略之一,就是多措并举发展国内花生生产替代大豆进口。因此,我国油料作物供给(食用油)安全和国际新形势新挑战已然将花生的优化施肥技术与耐冷优质高效生产推到一个前所未有的国家战略安全高度[2]。
辽宁省是我国重要的优质花生出口基地和产区之一,花生在辽宁省大田作物中其种植面积仅次于玉米和水稻。辽宁省土壤中黄曲霉菌含量极低,辽宁产区的花生(黄曲霉毒素显著低于其他产区)完全符合出口创汇的优级标准,这一优势也使得“辽宁花生”在全国乃至全球花生产业中品牌效应强,不可替代。低温是限制花生地理分布和生产的主要环境因子之一,冷害在全球花生生产中普遍存在[3],在我国尤其以北方高纬度区域、南方高海拔地区为甚。研究表明,大多数花生在幼苗期环境温度低于
12℃~16℃时,植株生长缓慢,严重时停止生长,叶片会出现脱水、萎蔫甚至枯死等现象[4–5]。前人大量研究表明,低温导致植物光合障碍,生长发育停滞,甚至死亡[6];低温胁迫对植物生长发育和光合碳同化过程产生负面影响[7]。研究发现,低温胁迫显著降低了水稻、玉米、向日葵、高粱、苹果幼苗的叶面积[8–9],抑制了玉米根系生长和干物质积累等[10]。低温胁迫可破坏光合作用的主要机构,包括叶绿体的结构、类囊体膜功能、光合电子传递和气孔运动等[11–12],降低植物光合作用水平,进而导致生物量降低、叶片坏死等问题[13]。
钙素作为一种植物必需的矿质养分,在维持细胞壁和细胞膜的结构完整性和功能稳定性上具有多重作用,同时钙离子作为信号物质在调节植物应对非生物胁迫中也发挥重要功能[14]。虽然土壤中含有大量钙素,但是作物在低温逆境下根系活力和叶片蒸腾速率骤降,时常发生严重吸钙障碍[15]。为此,叶部抗寒增钙技术作为植物应对(寒)冷逆境的外源调控措施,现已在水稻、玉米、番茄、苹果、黄瓜、烟草等作物上广泛应用[16–18]。外源钙预处理有助于维持细胞膜的完整性和抗氧化系统的平衡,从而减轻膜脂的过氧化反应[19–20]。课题组前期研究发现,叶面施钙具有显著提升花生耐冷性的调控效应[12–13],但是外源钙素的调控作用是否与解除低温光合障碍有关,这有待于进一步探究。为此,本试验应用人工气候室盆栽培养花生苗,同时采用外源钙离子(钙素正向调控)及其钙抑制剂EGTA (钙素负向调控)对花生苗进行预处理,从正反两方面深入探讨外源钙素对花生低温依赖性光合反馈抑制的调控作用,这对于最终实现花生生产中科学施用钙肥,以及进一步开发寒区花生高产优质施肥配套技术等具有重要指导意义。
1 材料与方法
1.1 试验设计
试验于2019年在沈阳农业大学土地与环境学院人工气候室内进行,人工气候室光合光子通量密度(PPFD)为 600 μmol/(m2·s),CO2浓度为 (400 ± 5) μmol/mol,相对湿度为 (60 ± 5)%。
试验以东北主栽花生品种“小白沙”为材料,利用48孔穴盘进行花生育苗,待长出第一片真叶时选取100株长势均匀一致的花生苗移栽至直径为13 cm 的圆柱盆中,平均分为 4个处理组:1) CK (夜温/昼温 20℃/28℃ + 叶面喷施超纯水),2) +H2O (夜温/昼温 9℃/15℃ + 叶面喷施超纯水),3) +Ca (夜温/昼温 9℃/15℃ + 叶面喷施 15 mmol/L CaCl2),4)+EGTA (夜温/昼温 9℃/15℃ + 叶面喷施 5 mmol/L EGTA)。待真叶展开一周后,连续两天(下午3:00—4:00)分别喷施超纯水、15 mmol/L CaCl2和 5 mmol/LEGTA处理,以上调控处理条件均为课题组前期筛选得出的最佳条件[4, 12–13]。于叶面喷施试剂处理后的第二天夜晚开始,模拟常见的花生苗期低温冷害情况,低温持续处理7天。于第7天的夜间恢复常温20℃,继续生长7天,整个试验处理周期为14天。
1.2 测定方法
1.2.1 生长指标的测定方法各处理随机选取生长均一的3株花生,于低温处理第7天和恢复常温后的第7天上午9:00—10:00测量生长指标。株高与茎粗分别利用直尺与游标卡尺测量,叶面积和相对叶绿素含量(SPAD值)分别利用LI-3000C叶面积仪和CCM-200 plus叶绿素仪测定。
1.2.2 光合及荧光参数的测定方法光合和荧光参数分别在低温的第1、4、7天及其常温恢复过程中的第1、4、7天的上午9:00开始测定,每株选取“倒三叶”作为指定叶片,各处理随机选取3株作为重复。光合气体交换参数采用GFS-3000 (WALZCo.,德国)测定,其中瞬时水分利用率(WUEt)以及气孔限制值(Ls)参照以下公式计算:
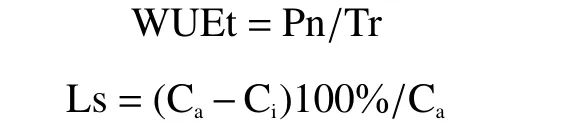
式中,Ca为大气CO2浓度,Ci为胞间CO2浓度。
最大光化学效率Fv/Fm、PSI实际量子产量Y(I)、PSI电子传递速率ETR (I)、供体侧限制引起的PSI处非光化学能量耗散的量子产量Y (ND)、受体侧限制引起的PSI处非光化学能量耗散的量子产量Y (NA)、PSII实际量子产量 Y (II)、PSⅡ电子传递速率ETR (II)、PSII处非调节性能量耗散的量子产量Y(NO)与PSII处调节性能量耗散的量子产量Y (NPQ)利用双通道叶绿素荧光仪Dual-PAM-100 (WALZCo.,德国)测定。
1.2.3 碳水化合物及活性氧含量的测定方法在低温处理的第7天和恢复常温的第7天取样,测定叶片碳水化合物和活性氧含量。各处理随机选取3株的倒三叶,105℃杀青30 min,65℃烘干,粉碎。葡萄糖、蔗糖、果糖、淀粉等含量测定参照薛应龙[21]的方法。丙二醛(MDA)含量采用硫代巴比妥酸(TBA)法测定,过氧化氢(H2O2)含量的测定参照Jana等[22]方法并加以改进。超氧阴离子的测定方法按照王爱国等[23]的方法并加以改进。
1.3 数据分析
利用 Excel 2016制表,Origin 2021作图。采用SPSS 19.0 软件进行单因素方差分析和最小显著差数法(LSD)检验,结果以3次生物学重复的平均值和标准误差表示。
2 结果与分析
2.1 低温胁迫及常温恢复条件下外源钙素对花生幼苗生长的影响
由表1可以看出,在低温处理7天时,喷施H2O和EGTA处理株高显著低于CK,而喷施Ca处理与CK无显著差异;而在恢复常温7天后,+H2O、+Ca和+EGTA 3个处理的株高之间无显著差异,但均显著低于CK。低温胁迫下,喷施H2O处理茎粗最大,而恢复常温后,施Ca处理茎粗最大,喷施EGTA处理最小。对照叶面积在低温胁迫和恢复常温后均最大,其他3个处理低温下无明显差异,但喷施Ca处理比施H2O和EGTA处理分别高42.9%和22.6%。低温胁迫显著降低了相对叶绿素含量SPAD,而恢复常温后,除了喷施EGTA处理SPAD值显著低于CK之外,其余处理与CK差异不显著。低温胁迫下,喷施Ca处理的根冠比虽然显著高于CK,但显著低于喷施H2O和EGTA处理;恢复常温后,喷施EGTA处理的根冠比依然显著高于CK。
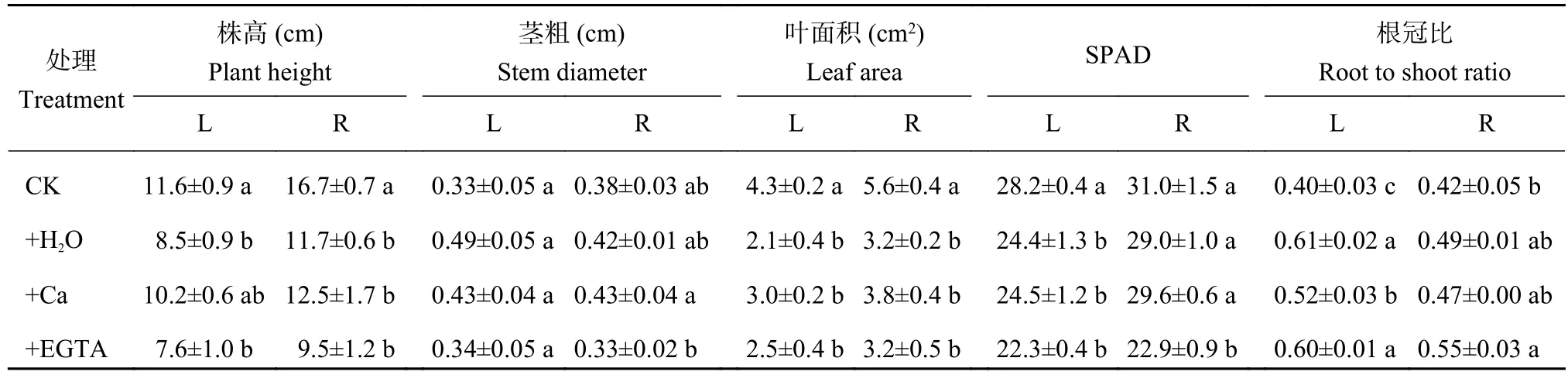
表1 低温胁迫7天(L)及恢复常温7天(R)时不同处理花生生长状况Table 1 Peanut growth at 7 days of low temperature stress (L) and at 7 days after recovery to regular temperature (R)
2.2 低温胁迫及常温恢复条件下外源钙素对花生叶片光合气体交换水平的影响
从图1可以看出,低温处理及恢复常温期间,净光合速率(Pn)整体呈先下降后上升逐渐趋于平缓的趋势,在低温胁迫7天时,喷施Ca和施H2O处理Pn低于CK,但高于喷施EGTA处理;恢复常温7天后,喷施Ca处理的Pn与CK无显著差异。细胞间二氧化碳浓度(Ci)随着低温胁迫天数的增加,整体呈先增长后下降的趋势。低温胁迫下喷施EGTA处理的气孔导度(Gs)最低,喷施Ca处理与CK差异最小,恢复常温后,喷施H2O与喷施EGTA处理Gs有增加趋势。蒸腾速率(Tr)随低温胁迫天数增加逐渐降低,恢复常温后逐渐趋于平稳,喷施Ca处理Tr值与CK无显著差异。瞬时水分利用率(WUEt)随低温胁迫天数增加逐渐降低,恢复常温后逐渐趋于平稳,低温胁迫下,以喷施Ca处理最大。低温处理及恢复常温期间,气孔限制值(Ls)整体呈先下降后上升的趋势,低温胁迫1~4天快速下降,恢复常温后上升趋势明显。综上,随着低温胁迫时间的延长,花生叶片Pn、Gs、Tr也逐渐降低,Ci逐渐上升,恢复常温7天后,喷施Ca处理的Pn、Tr、WUEt与CK无显著差异。
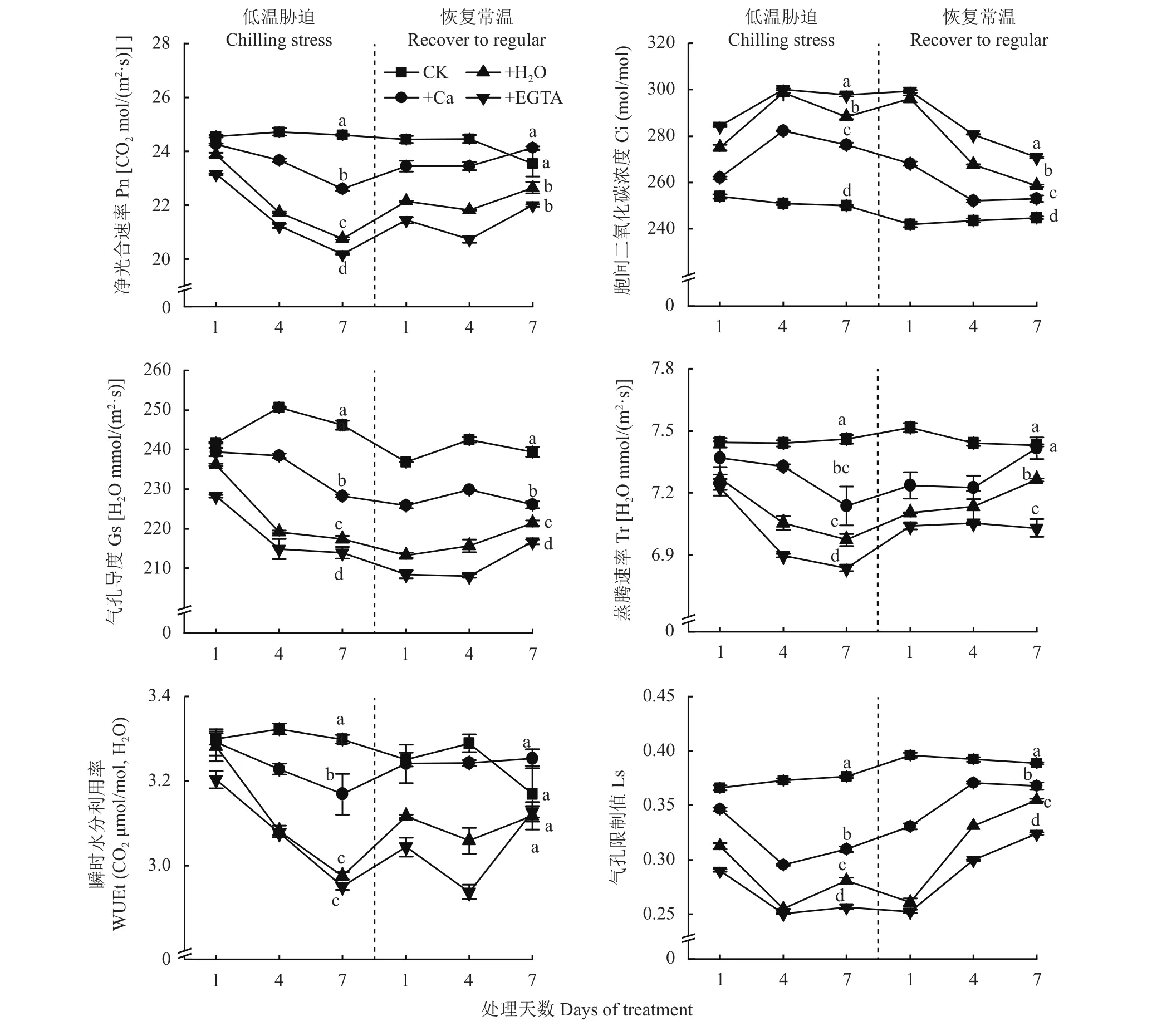
图1 低温胁迫及恢复常温后各处理花生气体交换参数Fig. 1 Leaf gas exchange parameters at different days of chilling stress and recovery to normal temperature under different treatments
2.3 低温胁迫及常温恢复条件下外源钙素对花生最大光化学效率Fv/Fm的影响
图2表明,PSII最大光化学效率Fv/Fm随着低温胁迫时间的延长呈现逐渐降低趋势,3个低温处理变化趋势相似,随低温胁迫时间延长,喷施EGTA处理Fv/Fm下降速率最大,喷施H2O和喷施Ca处理Fv/Fm下降的趋势与喷施EGTA处理相比较为缓慢;在低温胁迫第7天,3个低温处理与CK相比有显著差异,其中施Ca处理Fv/Fm大于喷施H2O和EGTA处理;在恢复常温的1~4天,除CK外的3个处理Fv/Fm呈快速上升的趋势,喷施Ca处理在恢复常温第7天Fv/Fm恢复到CK水平。
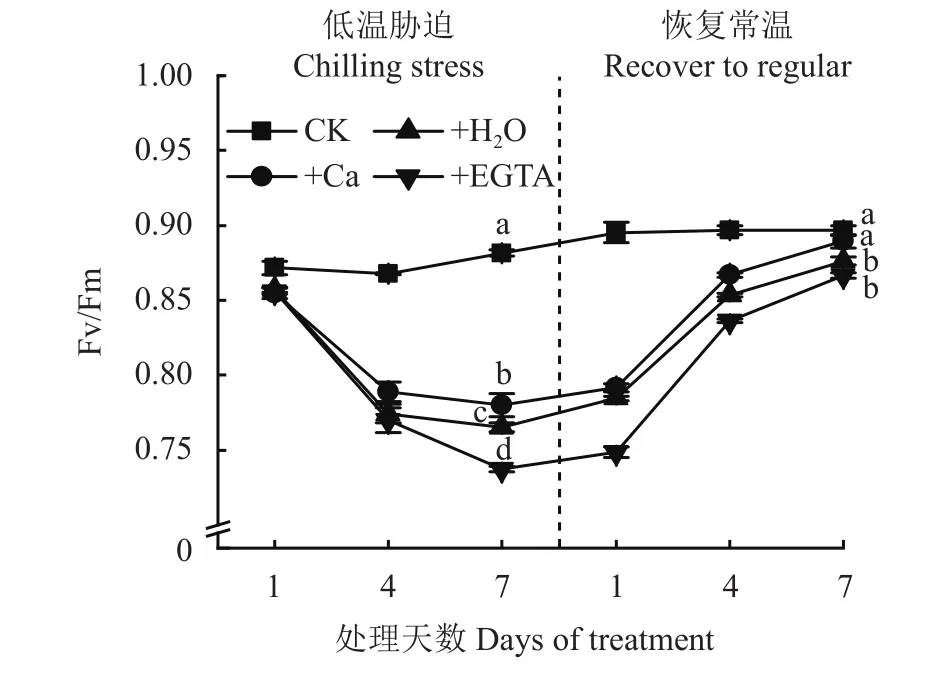
图2 低温胁迫及其常温恢复条件下各处理花生叶片PSII最大光化学效率(Fv/Fm)Fig. 2 The maximum quantum yield of PSII (Fv/Fm) in peanut leaves at different days of chilling stress and recovery to normal temperature under different treatments
2.4 低温胁迫及常温恢复条件下外源钙素对花生叶片PSI和PSII光化学活性的影响
由图3可以看出,低温胁迫1~7天时,+H2O、+Ca和+EGTA 3个处理的 Y (I)和 ETR (I)都显著降低,但喷施Ca处理降低最少;恢复常温后,+H2O、+Ca和+EGTA 3个处理的 Y (I)和 ETR (I)均呈上升趋势,且在恢复常温后的1~4天ETR (I)增加速率最大。在低温胁迫1~7天时,Y (ND)和Y (NA)呈上升的趋势,其中施Ca处理的Y (ND)最大,CK最小。Y(II)和ETR (II)总体趋势与Y(I)和ETR(I)相似,其中在1~4天的低温胁迫期间ETR(II)下降速率最大,Y(II)在恢复常温第7天时达到CK水平。随着低温胁迫时间延长,Y (NO)显著上升,喷施H2O和EGTA处理的Y (NPQ)与Y (NO)高于施Ca处理,低温条件下的+H2O、+Ca和+EGTA 3个处理间差异显著。
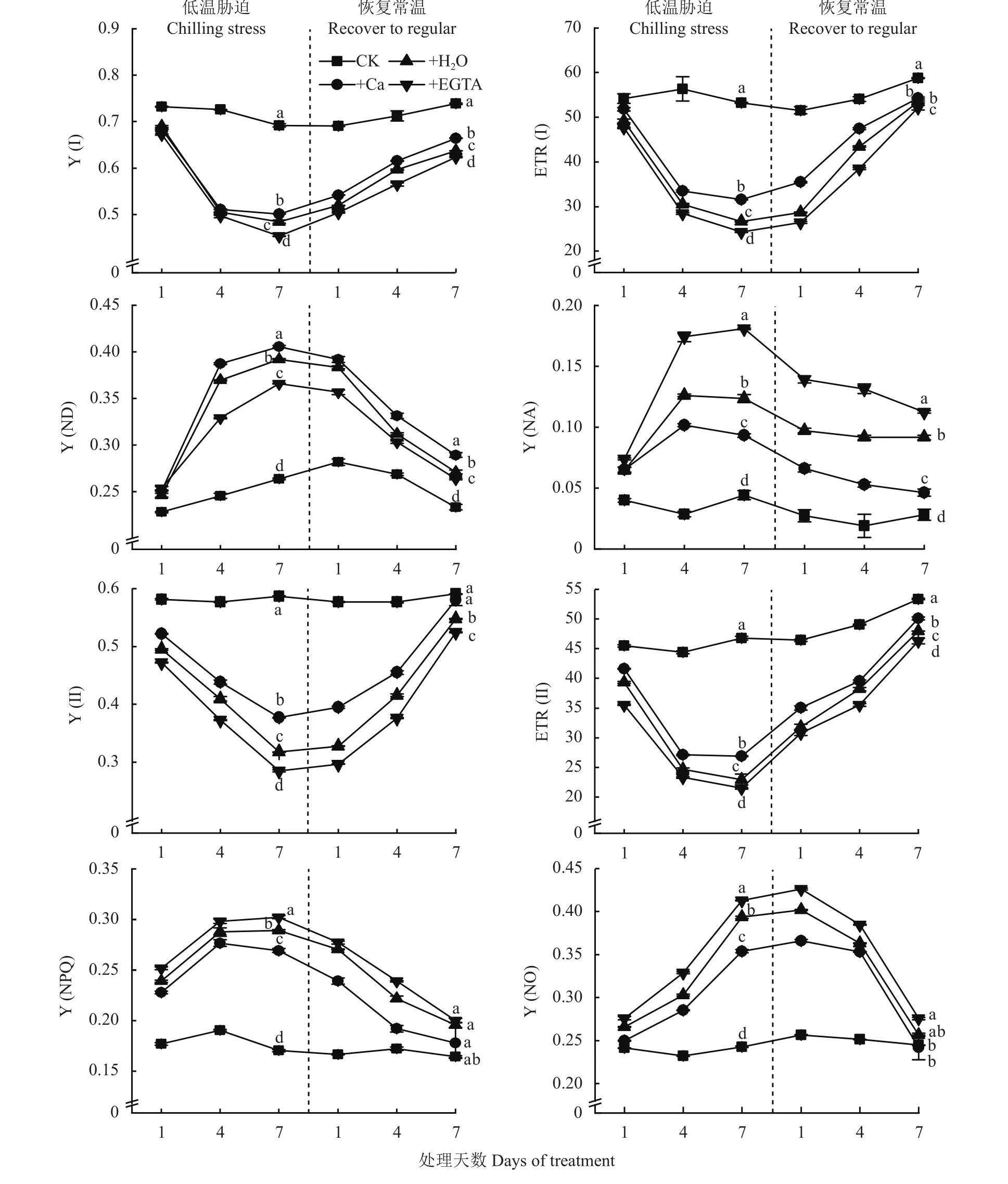
图3 低温胁迫及其常温恢复条件下各处理花生叶片PSI和PSII光化学活性Fig. 3 PSI and PSII photochemical activities in peanut leaves at different days of chilling stress and recovery to normal temperature under different treatments
2.5 低温胁迫及常温恢复条件下外源钙素对花生叶片非结构性碳水化合物积累的影响
由图4可以看出,在低温处理7天时,葡萄糖含量+Ca处理显著高于其他处理,+H2O和+EGTA处理显著低于对照;在常温恢复7天,+H2O、+Ca和+EGTA显著低于对照。与CK相比,3个低温处理的蔗糖含量显著增加。在恢复常温后,蔗糖含量都大幅度降低,其中+Ca处理与对照最相近。胁迫3个低温处理叶片果糖含量明显高于对照,其中+Ca处理最高,其次为+H2O和+EGTA处理。恢复常温7天,果糖含量大幅度降低,但处理间仍存在显著差异,以+EGTA处理的果糖含量最高。低温胁迫下+H2O处理淀粉含量最高,+Ca处理与对照无显著差异,恢复常温后淀粉含量增加,+Ca处理与对照无显著差异。
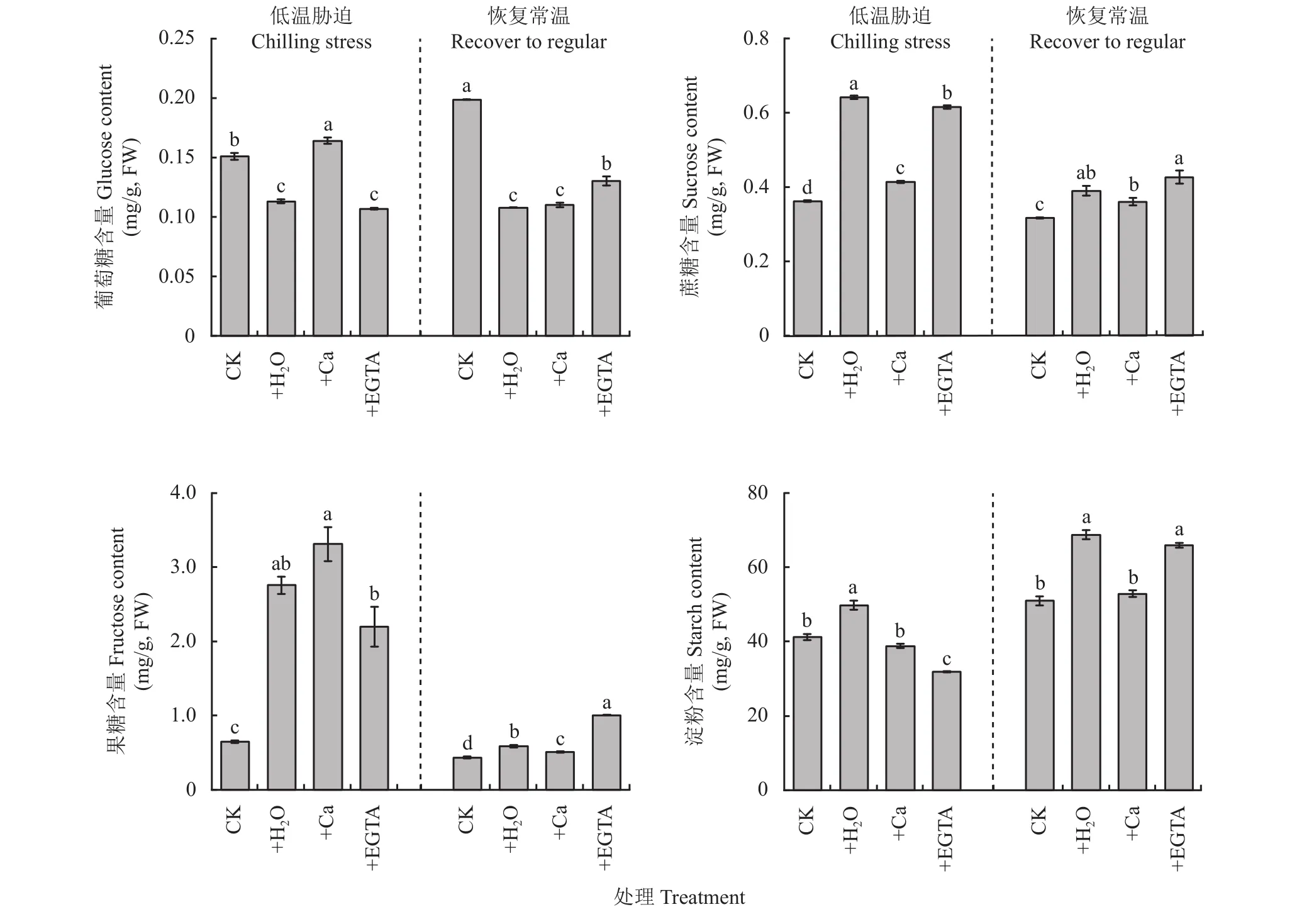
图4 低温胁迫7天及恢复常温7天各处理花生叶片非结构性碳水化合物积累量Fig. 4 Non-structural carbohydrate accumulation in peanut leaves at 7 days of chilling stress and recovered to regular temperature under different treatments
2.6 低温及常温恢复条件下外源钙素对花生叶片活性氧积累的影响
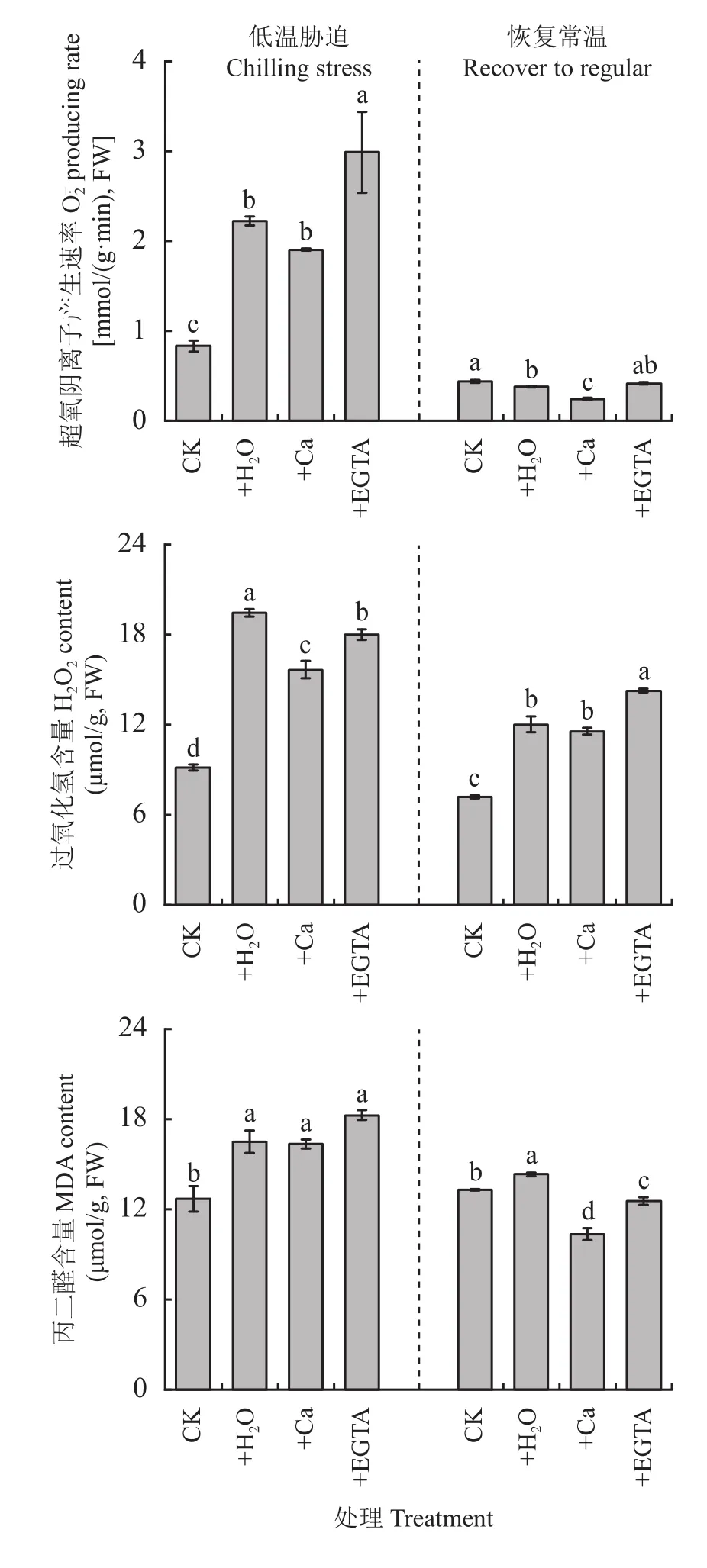
图5 低温胁迫7天及恢复常温7天各处理花生叶片活性氧积累量和丙二醛含量Fig. 5 The reactive oxygen accumulation and MDA content in peanut leaves at 7 days of chilling stress and recovered to regular temperature under different treatments
3 讨论
低温影响植物的生长发育,减少生物量积累,低温胁迫严重时造成不可逆伤害。花生属于典型喜温作物,低温环境显著影响花生生长发育,甚至造成缺苗断垄、空壳减产,导致严重经济损失。很多研究表明,在逆境条件下外源钙能有效提升气孔导度和净光合速率等[23–24];同时也可提升Rubisco[25]与1,5-二磷酸核酮糖羧化酶的活性、增强类囊体中电子传递速率等进而增强光合效率[26]。本研究表明,低温胁迫下与喷施H2O相比,叶面喷施外源Ca2+增加了花生株高、叶面积和叶绿素含量,也显著缓解了低温依赖性光损伤;有效缓解了花生叶片中非结构性碳水化合物的过量积累,增强了光合能力;有效降低了、H2O2和丙二醛的过量积累,保护了光合机构。前人研究表明,外源Ca2+间接缓解了夜间低温胁迫造成的Gs和Tr的骤降,维持了Ci,防止了Pn的下降,Ca2+改善了光合碳水化合物在韧皮部的装载和转运等,外源Ca2+可以显著修复低夜温造成的光合膜损伤,缓解番茄、苹果、烟草、花生等植物的光系统损伤,特别是缓解PSI光抑制[27–28]。以往的观点认为,植物PSI比PSII更稳定而不易受到光损伤,然而新的研究表明,PSI复合物基质侧的铁硫簇极易受到光系统中活性氧簇(reactive oxygen species, ROS)的破坏[29–30],尤其是波动光强和光强–温度交叉胁迫下PSI的光损伤更为严重,甚至PSI的光损伤要早于PSII。本研究发现,外源Ca2+有效缓解了低温胁迫对花生生长的不利影响,特别是有效减缓了Fv/Fm的降低,这说明外源Ca2+有效恢复了低温条件下花生叶片PSII的功能,降低了叶片中蔗糖的积累,降低活性氧带来的损伤,有效提高了花生叶片的光合活性[31]。
前人研究表明,植物可溶性碳水化合物的积累在冬季抗寒生存中起着重要的作用[32],通常蔗糖是在低温条件下积累的游离糖[33],另有国内外学者认为过多的碳水化合物积累使光合碳同化作用水平降低,并认为造成生物量减少的原因是碳水化合物的反馈抑制作用[34]。本研究发现,低温胁迫使花生幼苗叶片中蔗糖、果糖及淀粉大量积累,Pn降低,ETRI和ETRII下降;喷施外源Ca2+减缓蔗糖、果糖、淀粉等的过度积累,减缓了ETR和Pn的降低,说明喷施Ca2+可减缓由非结构型碳水化合物过度积累造成的光合效率降低,从而提高了低温胁迫下花生叶片的光合能力。Zhou等[35]、何洁等[36]研究认为,形成的大量淀粉粒会改变二氧化碳向细胞内扩散的途径,影响叶绿体中二氧化碳的吸收与运输使光合效率降低,出现光合速率和呼吸速率降低、同化物质运转缓慢等应激反应,导致淀粉及可溶性糖的过度积累。由我们低温胁迫后的恢复常温的实验结果可知,花生叶片中蔗糖、葡萄糖和果糖含量都显著降低,淀粉含量依旧较高;外源施Ca2+处理有效降低非结构型碳水化合物积累量,并可恢复至对照水平。低温阻碍叶片中蔗糖的运输,大量非结构型碳水化合物的堆积会显著抑制光合碳同化过程[37–38],这是使光合作用下降的主要原因之一;磷酸丙糖转化成更多的淀粉,淀粉来不及转化为蔗糖向外运出,导致蔗糖外运受到阻碍,进而叶片中积攒大量淀粉[39–41],最终产生严重的抑制作用。
有研究表明,碳水化合物与活性氧的关系密不可分,认为碳水化合物中的某些可溶性糖是活性氧代谢系统中重要的成分,可使植物抵抗过氧化损伤[42],也有研究表明活性氧的积累会上调可溶性糖的积累,从而适应非生物胁迫[43],活性氧在植物发育和生长过程及胁迫环境中起到调节作用,是不同胁迫与植物发育和生长控制之间的重要联系[44–46]。本试验结果显示,在低温胁迫后产生速率、H2O2含量以及MDA含量显著增加,这与罗娅等[47]研究结果一致,外源钙有效降低了、H2O2和丙二醛的大量积累,降低了光合机构的损伤。Ca2+在应对非生物胁迫及调节胞内信号也起着重要作用,可以抑制植物细胞膜脂过氧化,减轻活性氧的毒害作用。钙和ROS最初被认为是相互作用的,因为钙直接或者间接地对RBOHs(植物爆发式氧化酶,活性氧的主要来源)产生负调控,并可通过激活/抑制激酶/磷酸酶活性介导钙通道的开闭[48]。本试验结果显示,外源Ca2+有效缓解ROS大幅度增加,喷施钙离子抑制剂(EGTA)会显著增加产生速率,进而反向证明了Ca2+可有效降低活性氧的产生。在低温下ROS水平升高,ROS从源头抑制了蛋白质的合成,特别抑制了D1蛋白的合成,D1蛋白是光系统II反应中心的组成部分[49]。在不利于CO2固定的环境条件下,光合色素吸收过多的光能会抑制光损伤PSII的修复,从而增加光抑制的程度。因此,ROS不仅抑制PSII蛋白的合成,而且抑制PSII修复速率[50–51]。此外,钙可以稳定脂质层的基本结构,通过调节自身浓度减少ROS对膜脂过氧化损伤[52]。恢复常温后,喷施钙处理花生叶片丙二醛含量迅速下降[53],说明低温胁迫及其恢复过程中,叶面喷施Ca2+有助于降低花生叶片丙二醛含量,增加植株冷适应性。
4 结论
低温胁迫及其恢复过程中,外源Ca2+可以有效提升花生生长发育和光合作用水平,显著降低非结构型碳水化合物和ROS的过度积累,全面缓解低温光抑制。
猜你喜欢
现代农业科技(2022年5期)2022-12-14
舰船科学技术(2022年11期)2022-07-15
科学导报(2022年11期)2022-03-03
中国土壤与肥料(2021年5期)2021-12-02
农业资源与环境学报(2021年5期)2021-10-06
保鲜与加工(2021年1期)2021-02-06
智慧健康(2020年9期)2020-12-03
爆笑show(2015年3期)2015-05-08
妇女生活(2014年11期)2014-09-10
食品工业科技(2014年21期)2014-03-11
