
林下种植重楼和珠子参根际土壤与原生林地土壤特征差异△
2022-03-16 06:07高静王楠万修福徐蓉蓉关思静黄文静宋忠兴李铂张伟
中国现代中药 2022年2期
高静,王楠,万修福,徐蓉蓉,关思静,黄文静,宋忠兴,李铂,张伟
1.陕西中医药大学 药学院,陕西 西安 712046;
2.陕西中医药大学 陕西中药资源产业化省部共建协同创新中心/秦药特色资源研究开发国家重点实验室(培育),陕西 咸阳 712083;
3.中国中医科学院 中药资源中心/道地药材国家重点实验室培育基地,北京 100700;
4.宁强县中药材产业发展中心,陕西 汉中 724400
森林生态系统的功能健全离不开林下植被的有序参与,养分含量、水分资源及光照强弱等能够影响森林生态系统物种多样性和群落稳定性的关键因素都会随着林下植被的更替而发生变化[1]。已有研究通过对多个气候区域不同林分构成下的灌木层、草本层及苔藓层进行去除与添加植物物种试验发现,土壤理化性质和酶活性的差异是林下植被变化导致土壤微生物群落特征改变的关键因素[2]。因此,探查林下植被对土壤理化性质和酶活性的调控机制,对深入了解森林生态系统的运转模式具有重要的理论和实践意义。本课题组前期已经对重楼和珠子参根际土壤微生物量碳、氮、磷及其生态化学计量进行了相关探索[3],本研究通过将高海拔区域育苗的重楼和珠子参移栽到遮阴条件相似且林下植被均已去除的中海拔区域,对2 种移栽植物的根际土壤酶活性、养分含量及理化特征与中海拔区域原生林地土壤相应数据进行对比,分析2 种移栽植物对原生林地土壤特征的影响模式,为人工选择重楼和珠子参适宜生长区域土壤划分标准提供试验数据和理论参考。
1 材料
1.1 样品
试验样地位于陕西省西南隅秦岭和巴山两大山系交汇地带的宁强县鹰咀岩区域,此地区年平均降雨量1200 mm,年均温13 ℃。样地所处海拔1500 m左右,样地主要分布树种为白桦、麻栎、漆树、千金榆,华山松等,草本植物多为蒿类、白茅、莎草等。将样地草本全部挖除后,进行重楼和珠子参的移栽工作,移栽时间选取在9 月。本试验取样地点集中于上层乔木为千金榆和华山松的混交地带。实验材料由陕西中医药大学陕西中药资源产业化省部共建协同创新中心李铂副教授鉴定为重楼Paris polyphyllaSm.和珠子参Panax japonicusC.A.Mey.var.major(Burk.)C.Y.Wu et K.M.Feng。土壤采集于2019 年7 月上旬(当地高温时段,土壤化学过程较为剧烈),在重楼和珠子参林下栽培周边且未开发区域采集原生林地土壤。由于原生林地下层植物茂密,此土壤样本不区分是否为根际土。取样深度0~20 cm,在25 m×25 m 的区域内使用5 点法取样,混合土样除去植物残体和土壤动物。统一选择移栽苗龄3 年的重楼和珠子参根系不易抖落结合紧密的土壤部分(厚度1~3 mm)作为根际土,在连片种植区域为达到重复实验效果,3 组土壤获取区域间隔10 m 以上。上述土壤过2 mm 筛,置4 ℃冰箱保存待测定。
1.2 试药
盐酸、硫酸、甲基红、溴甲基绿、碳酸钠、酒石酸锑钾、抗坏血酸、钼酸铵、磷酸二氢钾、硫酸钾、氢氧化钠、柠檬酸、氢氧化钾、苯酚、甲醇、乙醇、丙酮、次氯酸钠、硫酸铵、葡萄糖、酒石酸钾钠、亚硫酸钠、草酸钠、高锰酸钾、脲、硝酸钾、硫酸银、氢氧化钙、碳酸钙均为分析纯,购自国药集团化学试剂有限公司;对氨基苯磺酸和α-萘胺盐酸盐均为分析纯,购自上海阿拉丁生化科技股份有限公司。
1.3 仪器
Vario MACRO cube 型元素分析仪(德国Elementar公司);K1160型凯氏定氮仪(海能未来技术集团股份有限公司);UV-1800PC型紫外分光光度计(上海美谱达仪器有限公司);FP640型火焰光度计(上海精工科技有限公司);Select TOC型总有机碳分析仪(德国Elementar公司);ICAP-7200型电感耦合等离子体发射光谱仪、Micro 17R型离心机均购于美国赛默飞世尔科技公司;230T型超声破碎仪(昆山市超声仪器有限公司);SpectraMax190型全波长酶标仪(美国分子仪器公司);JCS-2000型电子天平(中国凯丰集团有限公司);DIKW-4型水浴锅(北京中兴伟业仪器有限公司)。
2 方法
2.1 样品测定
土壤总碳(soil total carbon,TC)、有机碳(total organic carbon,TOC)、全氮(total nitrogen,TN)、全磷(total phosphorus,TP)、全钾(total kalium,TK)、速效钾(available kalium,AK)、速效磷(available phosphorus,AP)、有机质(total organic matter,TOM)、土壤微生物量碳(soil microbial biomass carbon,MBC)、土壤微生物量氮(microbial biomass nitrogen,MBN)、土壤微生物量磷(microbial biomass phosphorus,MBP)、铵态氮(NH4+-N)、硝态氮(NO3--N)采用元素分析仪、硫酸-重铬酸钾氧化容量法、浓硫酸消煮-凯氏定氮法、氢氧化钠熔融-钼锑抗比色法、火焰光度法、重铬酸钾外加热法、三氯甲烷熏蒸K2SO4法、三氯甲烷熏蒸-容量分析法、Brookes 法、分光光度法等进行测定[4-9]。土壤脲酶(urease,URE)、蔗糖酶(sucrase,SUC)、酸性磷酸酶(acid phosphatase,ACP)、过氧化氢酶(catalase,CAT)、硝酸还原酶(nitrate reductase,NR)、亚硝酸还原酶(nitrite reductase,NiR)活性采用苯酚钠-次氯酸钠比色法、3,5-二硝基水杨酸比色法、对硝基苯磷酸盐比色法、高锰酸钾滴定法、酚二磺酸比色法,苯磺酸-乙酸-α萘胺比色法测定[10-14]。土壤pH、电导率(electrical conductivity,EC)和含水量(water content,WC)分别使用pH计、电导率仪和称质量法测定。
2.2 数据统计与分析
使用主成分分析(principal component analysis,PCA)对数据进行降维处理,进行PCA 时将数据进行如下标记:TOC(X1)、TN(X2)、TP(X3)、TK(X4)、MBC(X5)、MBN(X6)、MBP(X7)、TOC∶TN(X8)、TOC∶TP(X9)、TOC∶TK(X10)、TN∶TP(X11)、TN∶TK(X12)、TP∶TK(X13)、MBC∶MBN(X14)、MBC∶MBP(X15)、MBN∶MBP(X16)、NH4+-N(X17)、NO3--N(X18)、AP(X19)、AK(X20)、pH(X21)、EC(X22)、WC(X23)、ACP(X24)、NiR(X25)、NR(X26)、CAT(X27)、URE(X28)、SUC(X29)、TC(X30)、TOM(X31)。采用单因素方差分析(One-way ANOVA)检验各指标不同处理间的差异,最小显著性差异法(LSD)进行多重比较,显著性水平为P<0.05。采用t-value 双序图研究土壤特征对酶活性的影响,当箭头完全落入空白或灰色圆圈,分别代表与限定酶活性呈现显著正相关或显著负相关。使用冗余分析(RDA)将3 种不同来源土壤特征的多变量集中于同一坐标系中,直观反映因子变化的贡献差异。由于TN 与TOM、TC 与TOC 在分析时有较大的重叠,因此在t-value双序图与RDA 中去除TOM 与TC 数据。MBC、MBN、MBP、生态化学计量等相关分析已在本课题组前期发表文章中进行讨论[3]。
3 结果与分析
3.1 不同来源土壤养分含量和理化性质比较
由图1 可知,珠子参根际土壤TC 和TOC 质量分数显著高于重楼根际和原生林地土壤的TC 和TOC 质量分数(P<0.05),TC 是后两者的1.59 倍和1.58 倍,TOC 是后两者的1.92 倍和1.86 倍,而重楼根际和原生林地土壤的指标之间差异无统计学意义。珠子参根际土壤TN 质量分数显著高于原生林地土壤TN 质量分数(P<0.05),达到后者的1.36 倍;原生林地土壤TN 质量分数显著高于重楼根际土壤TN 质量分数(P<0.05),是后者的1.07倍。珠子参根际土壤TOM 质量分数显著高于重楼根际和原生林地土壤TOM 质量分数(P<0.05),分别达到后两者的1.92倍和1.87倍,而重楼根际和原生林地土壤的指标之间差异无统计学意义。重楼根际土壤TP 质量分数显著高于珠子参根际和原生林地土壤TP 质量分数(P<0.05),分别达到后两者的1.11 倍和1.14 倍,珠子参根际和原生林地土壤TP质量分数之间差异无统计学意义。重楼根际土壤AP 质量分数显著高于珠子参根际土壤AP 质量分数(P<0.05),达到后者的5.5 倍;珠子参根际土壤AP 质量分数显著高于原生林地土壤AP 质量分数(P<0.05),是后者的2.57 倍。珠子参根际土壤TK质量分数显著高于重楼根际和原生林地土壤TK质量分数(P<0.05),达到后两者的1.12 倍,重楼根际和原生林地土壤TK 质量分数之间差异无统计学意义。原生林地土壤AK质量分数显著高于珠子参根际土壤AK 质量分数(P<0.05),达到后者的1.63 倍;珠子参根际土壤AK 质量分数显著高于重楼根际土壤AK质量分数(P<0.05),是后者的2.56倍。重楼根际土壤NH4+-N 质量分数显著高于原生林地土壤NH4+-N 质量分数(P<0.05),达到后者的1.12 倍;原生林地根际土壤NH4+-N 质量分数显著高于珠子参根际土壤NH4+-N 质量分数(P<0.05),是后者的1.04 倍。珠子参根际土壤NO3--N 质量分数显著高于重楼根际土壤NO3--N 质量分数(P<0.05),达到后者的3.74 倍;重楼根际土壤NO3--N 质量分数显著高于原生林地土壤NO3--N 质量分数(P<0.05),是后者的1.75 倍。由表1 可知,3 种土壤间pH、EC 及WC 差异均有统计学意义(P<0.05),pH 为原生林地土壤>重楼根际土壤>珠子参根际土壤,EC 和WC 为重楼根际土壤>珠子参根际土壤>原生林地土壤。上述结果表明植物会通过对碳、氮、磷、钾相关元素的获取间接影响土壤环境,进而改变土壤性质。
表1 不同来源土壤理化性质特征(,n=3)
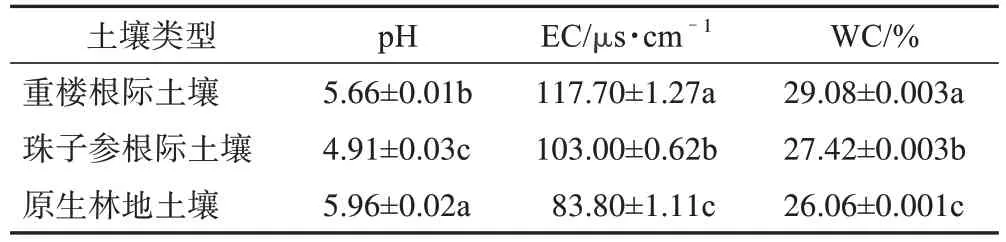
表1 不同来源土壤理化性质特征(,n=3)
注:不同小写字母表示组间P<0.05。

图1 不同来源土壤养分相关元素质量分数(, n=3)
3.2 不同来源土壤酶活性的比较
由图2 可知,原生林地土壤SUC 活性显著高于重楼根际和珠子参根际土壤SUC 活性(P<0.05),达到后两者的1.30 倍和1.23 倍,而重楼和珠子参之间差异无统计学意义。原生林地土壤URE 活性显著高于重楼根际土壤URE 活性(P<0.05),达到后者的1.33 倍;重楼根际土壤URE 活性又显著高于珠子参根际土壤URE 活性(P<0.05),达到后者的1.22 倍。原生林地土壤CAT 活性显著高于珠子参根际土壤CAT活性(P<0.05),达到后者的1.19倍;重楼根际土壤CAT活性与其他2种土壤之间差异无统计学意义。原生林地土壤NiR活性显著高于珠子参根际土壤NiR活性(P<0.05),达到后者的1.15倍;珠子参根际土壤NiR 活性显著高于重楼根际土壤NiR活性(P<0.05),达到后者的1.18倍。3种土壤间的NR 和ACP 活性差异无统计学意义。酶活性差异受到植株自身生理状态和环境条件的双重影响,一段时间内的酶活性只能表征当前时期不同植被类型和微环境小气候对不同酶类活动的影响。
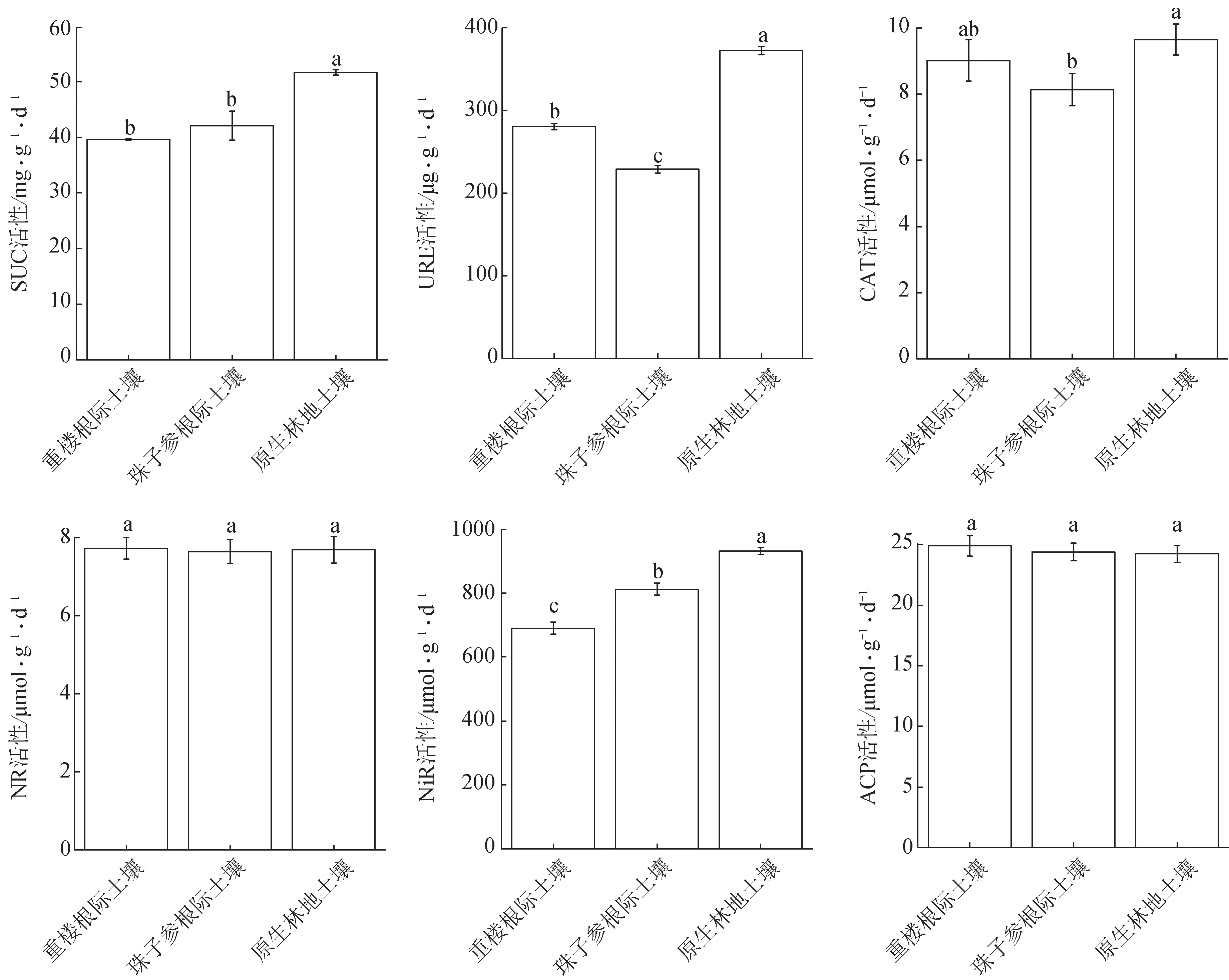
图2 不同来源土壤酶活性特征(, n=3)
3.3 土壤特征与酶活性的相关性
从t-value 双序图(图3)可以看出,TOC∶TP和TN∶TP 与ACP 活性呈现显著负相关关系。AK、MBC∶MBP、MBN∶MBP、TOC∶TP、TOC∶TK、TOC∶TN、TN∶TP、TN∶TK、TOC、TN、TK、NO3--N、MBN 与NiR 活性呈现显著正相关关系,pH、MBC∶MBN、TP∶TK、NH4+-N、MBP、TP、AP、WC、EC 与NiR 活性呈现显著负相关关系。TOC∶TP、TOC∶TK 和TN∶TP 与NR 活性呈现显著正相关关系,MBC∶MBN、TP∶TK 与NR活性呈现显著负相关关系。MBC∶MBN、TP∶TK与CAT活性呈现显著正相关关系,TOC∶TP、TOC∶TK、TN∶TP、TOC、NO3--N 与CAT 活性呈现显著负相关关系。pH、MBC∶MBN、TP∶TK、NH4+-N与URE 活性呈现显著正相关关系,MBN∶MBP、TOC∶TP、TOC∶TN、TOC∶TK、TN∶TP、TN∶TK、TOC、NO3--N、TN、TK、MBN与URE活性呈现显著负相关关系。没有指标与SUC 活性表现出显著相关性。指标间的相关关系往往会出现复杂的网状联系,酶活性虽然与养分含量及其相对占比有很高的关联度,但是由于植物生长状态及环境的不断变化,这种关系并不一定是时刻紧密对应的。
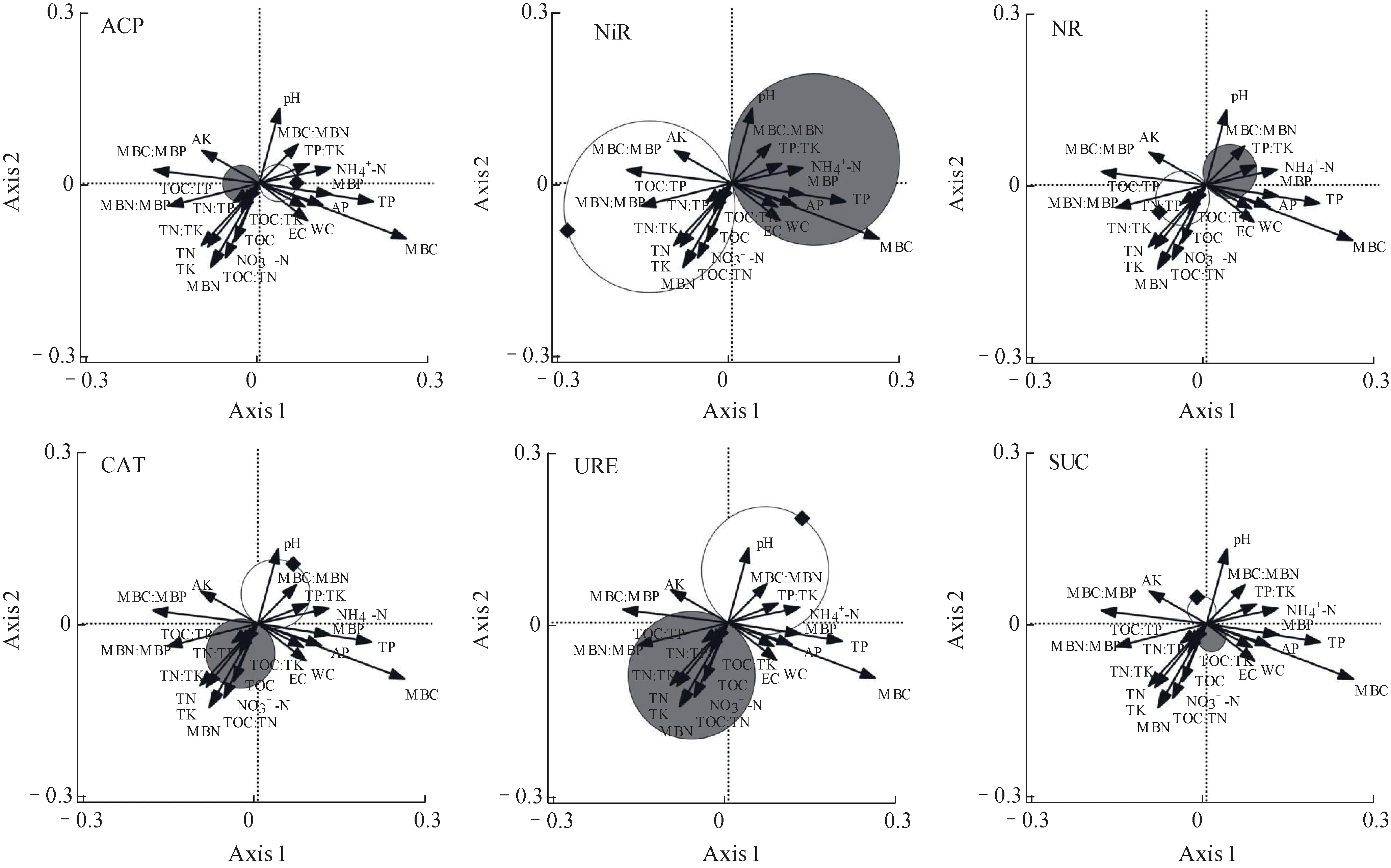
图3 土壤酶活性与土壤特征因子间的响应关系
3.4 土壤特征的PCA
从表2 可知,前三轴能够解释95.424%的变异(第一轴58.426%、第二轴33.063%、第三轴3.935%)。主成分表达式为公式(1)~(3),主成分综合模型为公式(4)。

表2 土壤特征的方差分解主成分提取分析
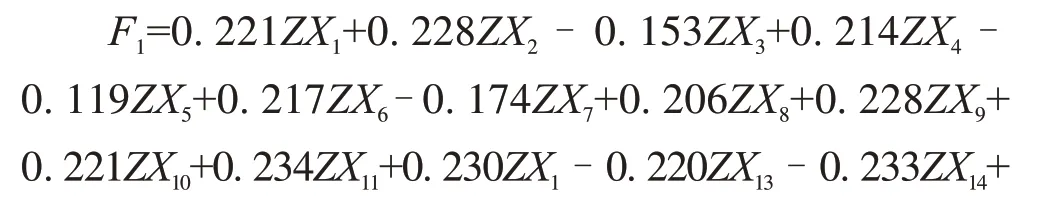
式中,ZX1~ZX31是X1~X31的标准化转换值。
主成分综合模型得分说明,3 种土样在养分状态和酶反应的综合条件下存在较大的差异(表3)。RDA 表明,重楼根际土壤特征与磷素类、NH4+-N 相关性较高,ACP 相比其他酶类有较高的参与度。珠子参根际土壤特征与大部分氮素类、TOC、TK 及NO3--N 相关性较高。原生林地土壤则是AK、SUC、URE、NiR、CAT较为活跃(图4)。
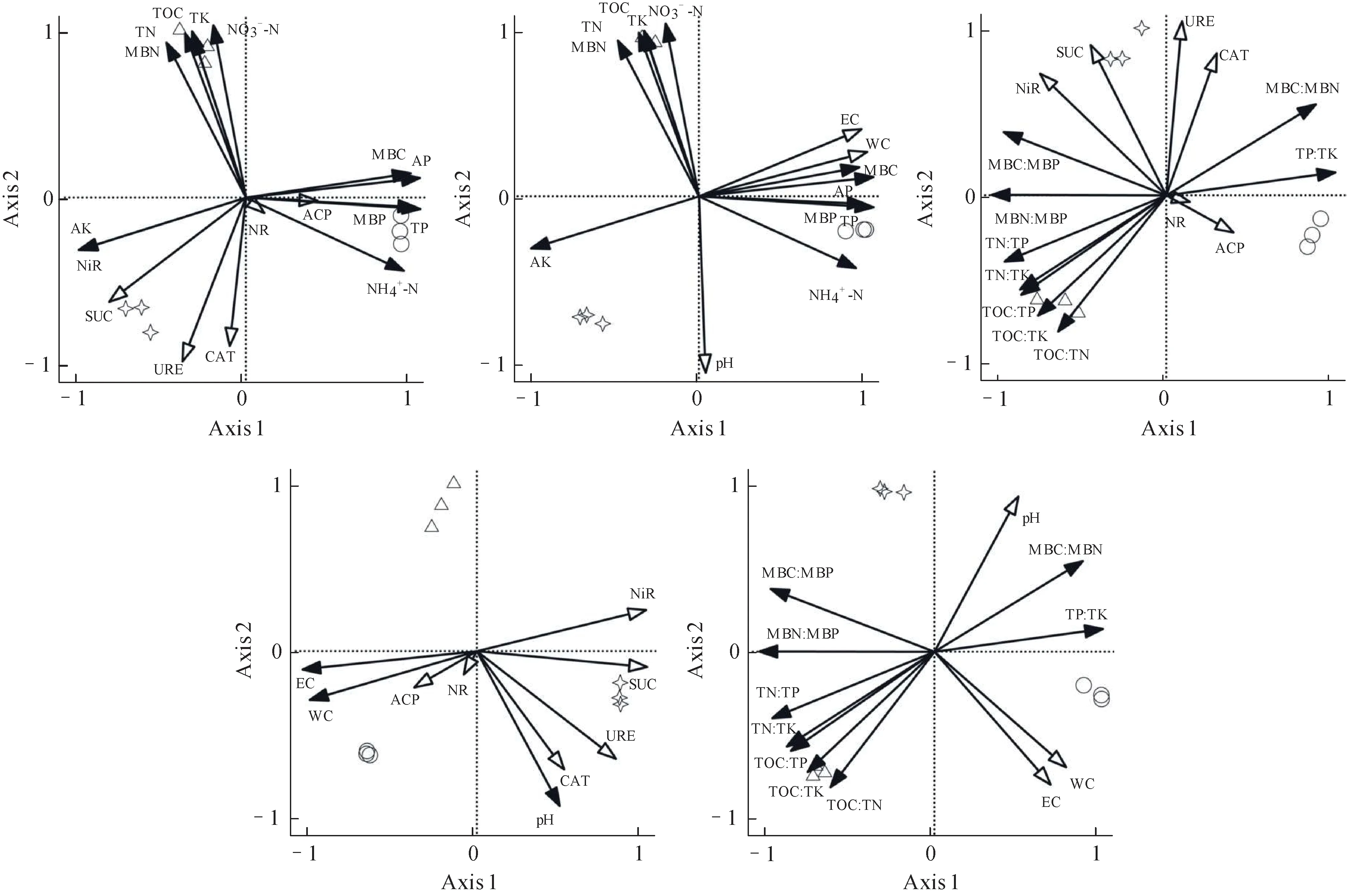
图4 不同来源土壤特征的RDA
4 讨论
根际是土壤养分流动与植物营养生理生态过程有机结合的重要区域。在本研究中,珠子参根际土壤TC 和TN 均高于重楼根际土壤和原生林地土壤。结合TOC 和TOM 数据可以推断,TOC 占比能够决定TC 总量,而TOM 对TN 含量上升具有较高的贡献。大量研究通过对不同地貌环境(山区、沙地、平原等)差异性植被类型(草地、作物、森林等)根际及土壤养分进行分析,同样得出TOC 与TC、TOM 与TN 间存在显著正相关关系[5,15-18]。林下移栽过程中深松处理属于保护性耕作,在此过程中的人为活动和耕作扰动使更多CO2和风积灰尘进入土壤,影响了土壤氮素的平衡和迁移转化,产生碳、氮元素表层富集效应[18],有利于浅根系植物的吸收。在本研究中,珠子参根际土壤TC 和TN 较高的原因可能是其相比于重楼对表层土壤碳、氮养分的利用率更高。有研究报道,通过对柠条Caragana korshinskiiKom.、短花针茅Stipa brevifloraGriseb.和蒙古冰草Agropyron mongolicumKeng 进行比较,发现根际与非根际TC 和TN 的含量存在不规律的变化形式,成因复杂,可能由植被差异介导的地表水资源汇流及再分配过程和根系对土壤理化性质及微生物种群的调节能力不同等方面原因导致[4]。根际土壤氮素含量既可以由根部土层养分决定,也可以被凋落物所含养分影响。然而,杉木Cunninghamia lanceolata(Lamb.)Hook.、林下中华鳞毛蕨Dryopteris chinensis(Bak.)Koidz.、淡竹叶Lophatherum gracileBrongn.、毛冬青Ilex pubescensHook.et Arn.和紫珠Callicarpa bodinieriLevl.等植物根际与非根际氮素含量被凋落物的差异化影响则说明,虽然凋落物也是林下土壤养分的主要来源之一,但凋落物所含养分的迁移是一个极其复杂的生态现象,有可能经历多次富集和再释放过程[19]。在本研究中,重楼根际土壤TP 和AP 均高于珠子参根际土壤和原生林地土壤。森林土壤磷素主要来源于矿物质风化,因此较难得到补充。磷素有向根际迁移的趋势,但是由于环境条件和植物吸收与转化能力差异,根际与非根际土壤磷素相对含量也随之变化[20]。对本试验样地分析后发现,珠子参种植密度大、间隙小,而重楼种植密度小且形态高瘦,由此可见,磷素的种间差异可能与淋溶程度有关[6]。钾离子移动性较强,AK 易受土壤质地和地形的影响[21],林下物种的差异加剧了本研究中AK含量的变化。
土壤酶来源广泛,植物根系、微生物及动物残体均能产生,且土壤中的多数生物化学与土壤环境运行、发展与转变过程均有土壤酶的参与[22-23]。由于土壤酶能转变固定态土壤养分,加速土壤有效态养分释放,因此土壤酶活性成为土壤肥力高低的衡量标准之一。土壤酶活性受到生物(植物、微生物与动物)和非生物(土壤水分、温度与理化性质)的综合影响[24]。在本研究中,重楼和珠子参根际土壤SUC 活性较为接近,但两者URE 活性差异较大。李毳等[11]对比了远志Polygala tenuifoliaWilld.和北柴胡Bupleurum chinenseDC.的根际土壤SUC 和URE的活性,也得到了与本研究相同的结果。土壤SUC能将蔗糖分解为葡萄糖和果糖,以便于植物和微生物吸收利用[22]。SUC 还能促进土壤易溶性营养物质增加,有利于为土壤微生物提供TOC,提高土壤微生物活性[13]。土壤URE 的酶促反应只能酶解尿素转变为氨,其活性与土壤氮素含量具有一定关联[23]。在本研究中,重楼根际土壤TN含量低于珠子参的现象可能暗示重楼对氮素的需求更大,导致微生物代谢更为活跃,这一点也可以被重楼根际土壤MBC含量较高所验证[3],所以造成重楼根际土壤URE 活性提高。土壤ACP 能对磷素进行分解,促进有机磷矿化,当解磷微生物感知环境磷素含量降低这一信号时,会以胞吐形式向土壤释放ACP,侧面帮助植物获得磷素[13]。然而,在本研究中ACP 活性高低与TP、AP 并没有直接的对应关系。SUC、URE 和ACP 是土壤中碳、氮和磷素流动与循环的关键酶,其活性大小时常受到土壤养分水平、微生物组成结构和植物生长状态的相互影响,因此ACP 活性与TP、AP含量的复杂关系及如何受到其他环境因素制约还需要进一步研究。过氧化氢类物质会对植物的生长发育产生危害,而CAT 能有效破坏土壤中这类物质[22]。有研究证实,CAT 活性和土壤有机质积累程度也有关联[25]。但也有文章和本研究结果类似,指出氮素含量不一定会影响到CAT 活性,由此可见氮素的转运效率差异也起到一定作用[20]。向土壤中添加肥料对NR活性通常有激活作用[25],然而也有研究证实毛竹Phyllostachys edulis(Carriere)J.Houzeau 原生林带土壤NR 活性最高且人工外源施加氮素对土壤NR活性具有显著抑制作用[13]。由于不同植被类型的根系形状和生理过程不同,引起土壤理化性质和微生物群落的改变,不同的反硝化作用体现差异化的土壤NR活性[26]。另一方面,底物和不同酶抑制物含量及底物分解速率是影响NR和NiR在氮素转化环节上相互关系的重要因素[27]。在本研究中,3 种土壤NR 活性差距不大,但NiR 活性表现出较大差异,可能是前期不同土壤NR 均能够促进反硝化作用,为NiR 提供了不同的底物数量[27],而此时的NR却因元素转化周期活性降低。此外,在本研究中原生林地土壤的6 种酶都保持不低的活性,可以表明植被物种多样性对酶活性具有一定的调节能力,能够有效提高土地生产力,增强生态系统组分的功能[28]。
土壤酶活性、养分含量和理化性质受到植被类型的显著影响,正、负响应均有发生。根据由主成分表达式得到的F1、F2、F3值和主成分综合模型得到的F值可以认为,本研究中的3 种土样在酶活性、养分含量和理化性质方面存在差异,重楼和珠子参移栽后对原生林地土壤特征的影响方向不同。在本研究中,RDA 显示重楼根际的磷素响应和珠子参根际的碳氮反应较为突出,而原生林地土壤的酶活性更加显著。本研究中3 种土样的SUC、URE、ACP活性与碳、氮、磷素含量变化不一致、相关性不显著的问题可能与移栽物种在适应新环境过程中能表现出类似入侵植物的特征,改变了养分循环效率有关[29]。通常,成熟林带土壤酶活性较高,这与本研究中原生林地土壤情况类似。然而,有研究表明,虽然近熟林酶促底物略有欠缺,但其部分酶活性和养分含量高于成熟林,原因归结于物种年轻、发育迅速且有效养分比重相对较高[30]。这一点也在本研究中重楼和珠子参的根际土壤中有所表现。因此,酶活性与养分含量相关性的变化,由植被组成、物种年龄、环境条件、采样时间等限制。此外,根部生理作用的周期变化也会带来酶活性与养分含量的剧烈变化,导致重楼与珠子参根际土壤养分相较于原生林地出现富集作用。本研究中RDA 结果显示,重楼、珠子参与土壤养分、酶活性的正、负相关关系和土壤p 值具有较大的关联。有研究也表明,常绿阔叶林下的土壤pH和养分含量适合重楼、珠子参的生长[31-32],且2 种植物的次生代谢产物与生长特性均与土壤养分含量呈现显著相关关系[33-36]。宁强县属于多雨气候类型地区,更应注意土壤养分的存在形式,NH4+-N有助于保护土壤肥力,NO3--N在潮湿的环境下容易流失,而NO3--N 增多伴随的土壤pH 降低对硝化细菌有抑制作用,影响酶活性[37]。
土壤特征与植被类型是一个相互影响的过程。与原生林地相比,进行林下种植后的土壤由于物种改变导致的养分需求变化必然会带来酶活性的差异。因此,应开展林下种植适应性研究,针对不同药用植物特征展开相应抚育工作,根据药用植物生长需求选择相应原生林地类别,缩短药用植物移栽后的适应周期,推动药用植物生态种植发展。
猜你喜欢
现代农业科技(2022年15期)2022-08-17
中国土壤与肥料(2021年5期)2021-12-02
四川农业科技(2021年7期)2021-12-01
建材发展导向(2021年9期)2021-07-16
中国果业信息(2020年10期)2020-12-15
考试与评价·七年级版(2020年5期)2020-10-29
红楼梦学刊(2020年2期)2020-02-06
诗潮(2019年10期)2019-11-19
当代陕西(2019年6期)2019-04-17
农民致富之友(2018年2期)2018-03-25
