
高温对冬小麦旗叶光合机构的伤害机制*
2022-03-11 03:38:24李向东杜思梦张德奇时艳华王汉芳邵运辉方保停程红建
中国生态农业学报(中英文) 2022年3期
杨 程,李向东**,杜思梦,张德奇,时艳华,王汉芳,邵运辉,方保停,程红建,位 芳
(1.河南省农业科学院小麦研究所/小麦国家工程实验室/农业部黄淮中部小麦生物学与遗传育种重点实验室/农业部中原地区作物栽培科学观测实验站/河南省小麦生物学重点实验室 郑州 450002;2.郑州大学农学院 郑州 450052)
小麦()是我国第二大粮食作物,小麦的稳产和高产对于我国粮食安全具有重要意义。黄淮麦区是我国冬小麦的重要产区,灌浆期高温是该区域小麦生产中常见的自然灾害。随着全球环境的恶化和气候变暖,小麦灌浆期遭遇高温胁迫的风险也将逐渐增大。有研究预测未来全球平均气温每升高1 ℃,全球平均小麦产量将下降6%。光合作用是作物产量形成的最终能量来源,灌浆期小麦叶片的光合作用直接影响小麦千粒重,是决定小麦产量的重要因素。因此,研究高温对小麦灌浆期旗叶光合功能的影响特别重要。
光合作用包括光反应和暗反应两个过程,在光反应过程中,植物利用吸收的光能驱动水的分解,释放出电子,并产生氧气,电子再经过光系统Ⅱ(PSⅡ)、质体醌(PQ)、细胞色素B6f 复合体(Cyt)、质体蓝素(PC)、光系统Ⅰ(PSⅠ)等一系列电子传递体传递给NADP+生成NADPH,同时产生跨膜质子梯度驱动ATP 的生成,NADPH 和ATP 再用于暗反应中的卡尔文循环,完成CO的固定。高温可以通过多种途径抑制植物叶片的光合作用。多项研究表明高温胁迫下核酮糖-1,5-二磷酸羧化酶(Rubisco)的活性显著下降,因此,Rubisco 被认为是高温胁迫下光合作用的主要抑制位点,也有研究表明高温下Rubisco 的活性下降主要是由于Rubisco 活化酶的活性受到了抑制。此外,高温也会破坏光系统Ⅱ的活性,而且会导致放氧复合体锰离子的脱落。类囊体膜的稳定性对于维持光系统Ⅱ的活性至关重要,研究表明类囊体膜不饱和脂肪酸比例的增加有利于维持PSⅡ高温下的活性。PSⅡ对高温的敏感程度已经成为衡量小麦以及其他农作物光合机构高温抗性的重要指标。前人对于高温下小麦光合作用的研究多集中于PSⅡ的整体活性,而对影响PSⅡ活性的具体过程关注较少,譬如PSⅡ对光能的吸收和传递、放氧复合体活性以及PSⅠ电子传递活性对高温的响应情况等。而且以往对于小麦叶片高温伤害机制的研究大多使用较少的小麦品种,所获得的结论缺乏广泛的代表性。
叶绿素荧光参数/表示PSⅡ的最大光化学效率,是反映PSⅡ活性的最常用的荧光参数。但是该指标的应用也存在一定的局限性,譬如在轻度胁迫下,或者在敌草隆(DCMU)、2,5-二溴-6-异丙基-3-甲基-1,4-苯醌(DBMIB)等特定位点电子传递抑制剂的处理下,/并没有显著的改变。因此,多种研究手段的协同使用对于研究高温胁迫下小麦光合电子传递链的状态和功能十分必要。
快速叶绿素荧光一般指暗适应后的叶片在饱和光的照射下从最小荧光值()上升到最大荧光()的过程,该过程与不同光合电子传递阶段的活性和状态密切相关,已经被广泛用于光合逆境生理的研究。光系统Ⅰ(PSⅠ)反应中心色素P700 处于氧化态时,可以特定地吸收820 nm 波长的光,通过动态监测叶片对820 nm 光的反射信号可以研究PSⅠ反应中心的氧化还原状态,且能同时推测PSⅡ向PSⅠ的电子传递状况。光合电子传递过程发生的氧化还原反应都是可逆的,当电子发生逆向传递时就会导致PSⅡ反应中心色素P680 与供体侧或者受体侧的电子传递体发生电荷重组,进一步引起P680 再激发形成激发态的P680*,P680*可以通过天线色素将能量以荧光的形式释放出来,称为延迟荧光。延迟荧光也可以用于光合电子传递过程的研究。近年来,多功能植物效率仪(M-PEA)的出现,使快速叶绿素荧光、延迟荧光、820 nm 光反射(MR)同步测定技术越来越多地用于逆境下光合作用的研究。
本研究拟使用快速叶绿素荧光、延迟荧光、820 nm光反射同步测定技术研究高温对35 个小麦品种离体旗叶光合电子传递过程的影响及作用机制,为我国抗高温小麦品种的选育和栽培管理技术的创新提供理论支撑。
1 材料与方法
1.1 试验材料
试验材料选用1941-2014年通过审定且在河南省大面积种植的35 个冬小麦品种(表1),种植于河南省平原新区河南省农业科学院实验基地,小区面积6 m,每个品种3 个重复。试验田肥力均匀,按照生产实际整地,2017年10月10日播种,行距20 cm,株距2 cm,采用人工播种。整个生育期采用合理统一的田间管理,适时施肥,浇水。灌浆前期(2018年5月9日),取各品种完整均匀一致的旗叶,用湿纱布包裹后带回实验室。参考杨程等和Liu 等的方法,将各小麦品种旗叶剪掉基部和叶尖部位,保留中间部位,然后将中间部位叶片剪成长约1 cm 的小段,在直径6 cm 培养皿中加入15 mL 蒸馏水,每个培养皿放入10 个均匀一致的旗叶片段,并使叶片漂浮在水面。每个品种均按照该方法设置3 个盛有叶片段的培养皿,分别置于设置不同温度22 ℃(CK)、30 ℃(T1)、38 ℃(T2)的光照培养箱进行处理,培养箱光照强度均为250 μmol·m·s,处理2 h 后,进行叶绿素快速荧光诱导动力学曲线、820 nm 光反射和延迟荧光的同步测定,每个品种测定6 个重复。
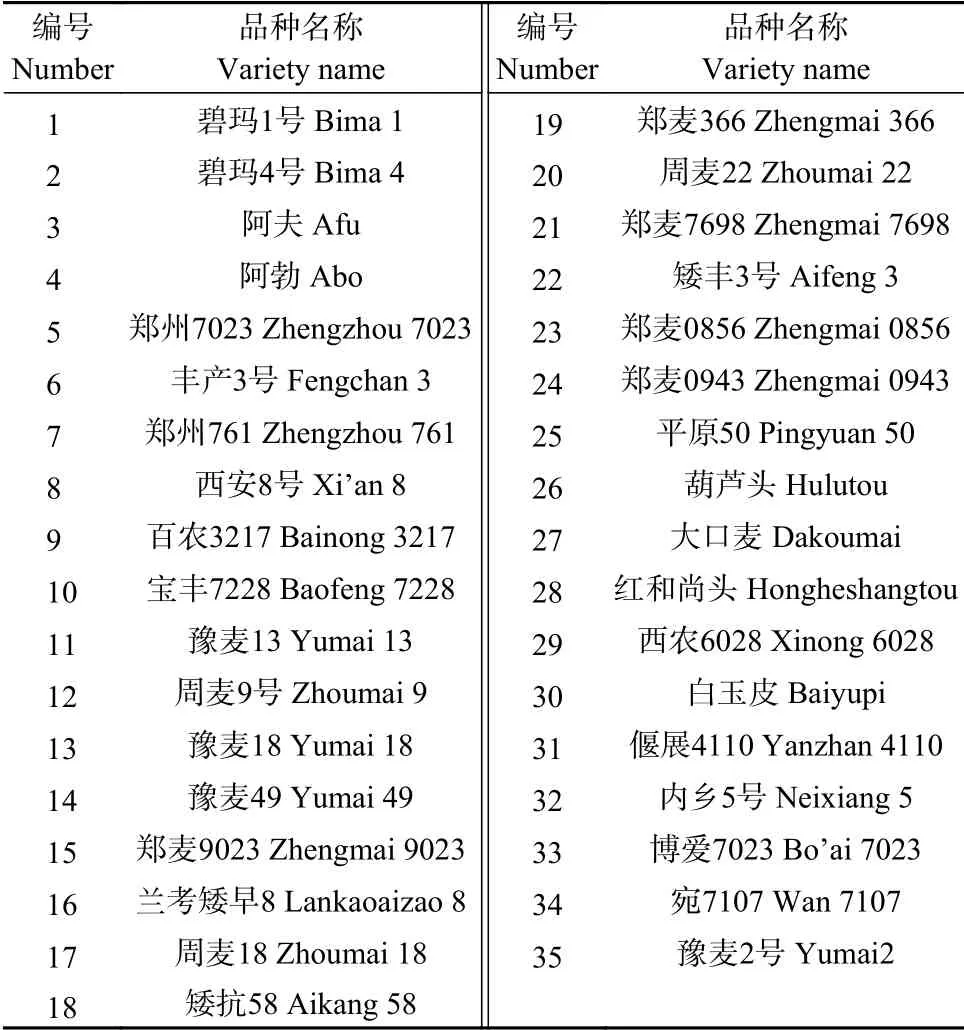
表1 选用的小麦品种Table 1 Wheat cultivars used in this study
1.2 叶绿素快速荧光诱导动力学曲线、820 nm 光反射和延迟荧光测定
叶绿素快速荧光诱导动力学曲线(OJIP)、820 nm光反射(MR)和延迟荧光(DF)使用多功能植物效率仪(M-PEA,Hansatech Instruments,UK)进行测定,参考Gao 等的方法,稍作修改:叶片测定前均在室温条件下采用仪器配套的暗适应夹进行30 min 的暗适应,然后用2 s 5000 μmol·m·s的红光同步测定快速荧光、延迟荧光和820 nm 的光反射,红光波长为(627±10)nm。同步测定时,从曝光300 μs 开始进行光暗转换,光下记录快速荧光和820 nm 的光反射信号,黑暗时记录延迟荧光信号,直至完成2 s 的测定过程。
OJIP 参数计算参考Strasser 等、杨程等的方法,具体见表2。

表2 叶绿素快速荧光诱导动力学曲线参数及公式Table 2 Parameters and formulas of rapid chlorophyll fluorescence inducing kinetic curve
叶片在暗适应后,在饱和红光照射下MR 信号呈先下降后上升的趋势,为了排除叶片之间的基础信号的差异,MR 的变化用MR/MR表示(MR表示820 nm 光反射信号的初始值),MR 的最大下降速率()通过MR/MR的1.1~2.0 ms 阶段的下降斜率计算,上升段的最大速率(-)通过30~100 ms 的上升斜率计算。
延迟荧光通过软件M-PEA data-analyzer V.4.4.3进行原始数据的分析和导出。
1.3 统计分析
数据显著性检验采用DPS v18.10 软件分析。
2 结果与分析
2.1 不同小麦品种叶片最大光化学效率高温响应规律及分类
最大光化学效率(/)是应用最广泛的荧光参数之一,通常用来反映植物逆境下的光抑制程度。从表3 可知,高温处理后各小麦品种的/均有所下降。通过K-均值聚类法对38 ℃高温处理(T2)下小麦旗叶的/下降幅度进行快速聚类,将35 个小麦品种分成了高温不敏感(G1)和高温敏感(G2)两组,其中高温不敏感组包含13 个小麦品种,高温处理条件下/下降幅度范围为1.4%~39.5%;高温敏感组包含22 个小麦品种,/下降幅度范围为46.7%~82.9%。

表3 不同类型小麦品种不同温度处理下的旗叶FV/FM 及其38 ℃处理下的下降幅度和分类结果Table 3 FV/FM of flag leaves at different temperatures and its decrease extents at 38 ℃,and classification results of different wheat cultivars
2.2 高温对小麦旗叶快速叶绿素诱导曲线的影响
充分暗适应后的小麦叶片在饱和光照射下PSⅡ从完全开放状态逐渐到完全关闭,荧光信号整个过程呈现典型的O-J-I-P 变化。通过对35 个品种的冬小麦旗叶进行高温处理(T1 为30 ℃,T2 为38 ℃),以22 ℃处理为对照,按照表1 中的分类结果分别进行平均。如图1所示,随着处理温度的升高,可以明显看到3 ms 荧光(J 点)和30 ms 荧光(I 点)的升高,高温敏感的小麦品种J 点和I 点荧光的升高幅度大于高温不敏感的小麦。

图1 高温对高温不敏感型(G1)和高温敏感型(G2)小麦品种旗叶叶绿素荧光诱导曲线的影响(曲线已标准化)Fig.1 Effect of high temperature on chlorophyll fluorescence induction curves of wheat flag leaves of high-temperature insensitive(G1)and sensitive(G2)cultivars(the curve has been standardized)
为了获得更多的PSⅡ的信息,将快速荧光诱导曲线的O-J 点进行标准化,从而可以看到K 点(0.3 ms)荧光的变化(图2A、2B),依照同样的方法对O-K 点标准化(图2C、2D),可以看到L 点(约150 μs)的变化。结果显示,G1 和G2 两组小麦品种在高温胁迫下K 点均下降;G1 品种L 点没有明显变化,而G2 组小麦品种高温胁迫下L 点显著上升。

图2 高温对高温不敏感型(G1)和高温敏感型(G2)小麦品种旗叶叶绿素荧光诱导曲线O-J 段和O-K 段荧光强度的影响Fig.2 Effect of high temperature on the fluorescence intensities of O-J segment and O-K segment of chlorophyll fluorescence induction curves of wheat flag leaves of high-temperature insensitive(G1)and sensitive(G2)cultivars
荧光参数、和是反映光合电子传递不同阶段效率的重要指标,表示PSⅡ捕获光能的效率,表示传递到Q- 下游的光能占吸收光能的比率,表示传递到PSⅠ末端的光能占PSⅡ吸收光能的比率,PI表示PSⅡ的综合性能指数。如图3所示,两组小麦品种、、和PI在高温胁迫下均呈下降趋势,但是高温敏感小麦(G2)在T2(38 ℃)处理下降幅度显著大于高温不敏感的小麦品种(G1)。o 表示PSⅡ耗散光能的比率,与变化趋势相反,两类小麦品种随着处理温度的升高,均呈上升趋势,其中高温敏感的小麦品种在T2 处理下的上升幅度显著大于高温不敏感的小麦品种。RC/CS表示单位面积PSⅡ有活性反应中心的数量,RC/CS呈现与相同的趋势,仅在T2 处理下与CK 呈现显著差异。表示传递到PSⅠ受体侧的电子占传递到Q-下游的电子的比率,在T2 处理下,显著上升,且高温敏感的小麦品种在T2 处理下上升幅度大于高温不敏感的小麦品种。
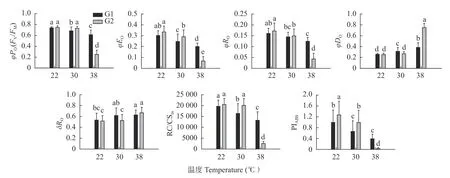
图3 高温对高温不敏感型(G1)和高温敏感型(G2)小麦品种旗叶叶绿素荧光参数φPo、φEo、φRo、φDo、RC/CSm、δRo、PIABS 的影响Fig.3 Effect of high temperature on chlorophyll fluorescence parameters φPo, φEo, φRo, φDo,RC/CSm, δRo and PIABS of wheat flag leaves of high-temperature insensitive(G1)and sensitive(G2)cultivars
2.3 高温对小麦旗叶820 nm 光反射的影响
PSⅠ的反应中心色素P700 在吸收光能后会释放电子转变为P700,P700能特异地吸收820 nm 的光,所以通过监测光下叶片对820 nm 光的反射变化可以获得PSⅠ反应中心的氧化还原状态的变化。在完全暗适应后,PSⅠ反应中心处于还原状态,在照光的初始阶段,由于PSⅡ的电子还未传到PSⅠ,因此P700处于快速积累状态,随着照光时间的延长和PSⅡ电子的到来,P700 的状态从氧化大于还原逐渐到氧化和还原的平衡,再到还原速率大于氧化速率,因此,MR 信号整体呈现先下降后上升的形态,在MR 信号的下降阶段,两个类型小麦品种在不同温度下均无显著差异,但是在上升阶段G2 组在高温下上升速率显著变慢且上升段的最大值出现明显下降(图4A、4B)。分别计算下降和上升的最大斜率,可以得到和两个参数,从图4C 和4D中可以看出,高温处理下,G1 和G2 组小麦品种叶片高温处理后与CK 处理相比均无显著差异,而G2 组小麦品种在T2 处理下下降幅度显著大于G1 组小麦品种(<0.05,图4D)。
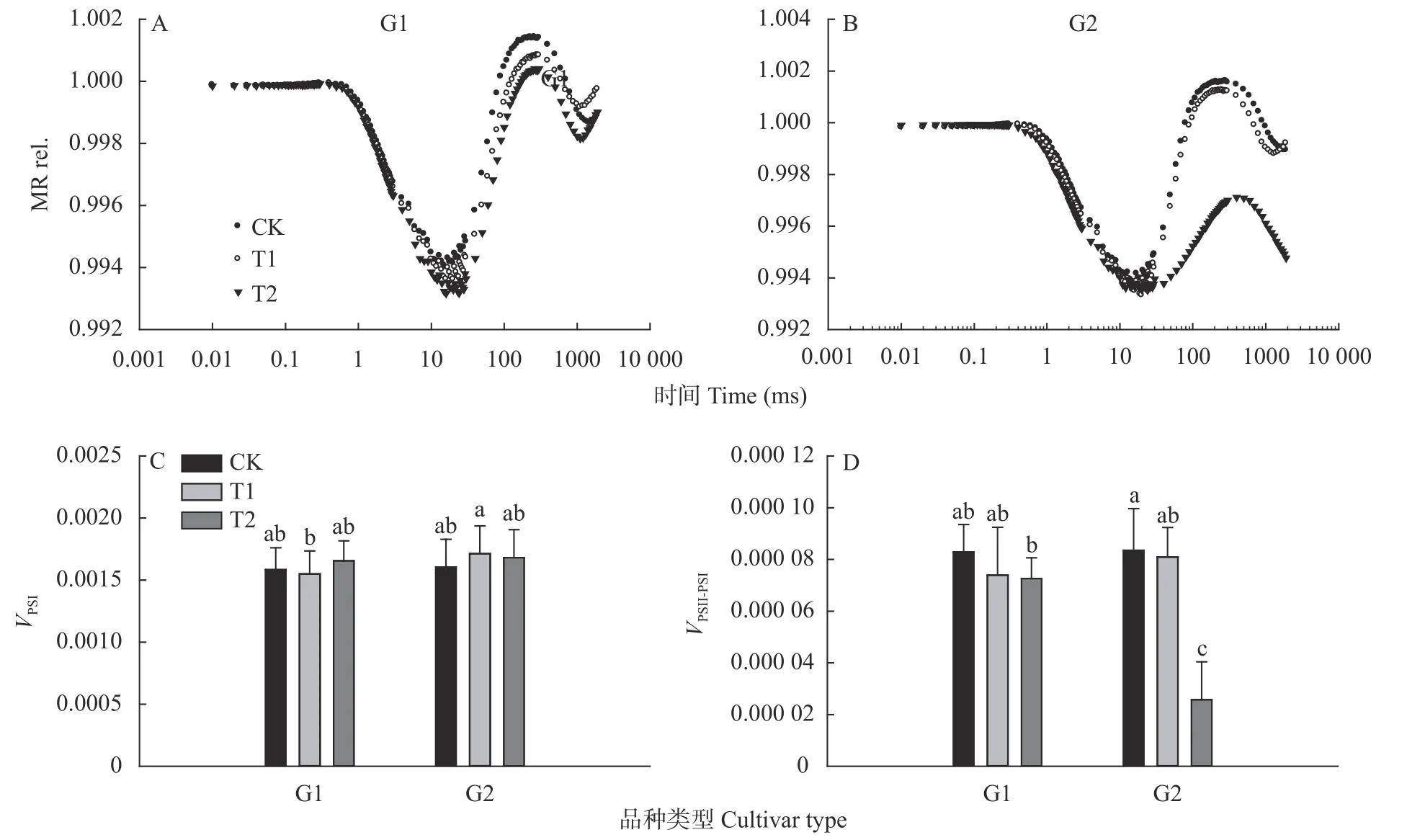
图4 高温对高温不敏感型(G1)和高温敏感型(G2)小麦品种旗叶820 nm 光反射(MR rel.)及其最大下降速率(VPSⅠ)和上升速率(VPSⅡ-PSⅠ)的影响Fig.4 Effect of high temperature on the light reflection at MR820nm(MR rel.)and its maximum decrease rate(VPSⅠ)and rise rate(VPSⅡ-PSⅠ)of wheat flag leaves of high-temperature insensitive(G1)and sensitive(G2)cultivars
2.4 高温对小麦旗叶延迟荧光的影响
延迟荧光与快速荧光一样蕴含着大量的光合电子传递的信息,在近年来逐渐用于各种逆境下的植物光合作用的研究。如图5所示,在PSⅡ从开放到逐渐关闭的过程中,延迟荧光信号(0.01~0.02 ms)呈现先上升后下降的趋势,随着处理温度的升高,I和I点的延迟荧光信号均逐渐下降,但是I点延迟荧光的下降幅度显著大于I,从而导致/的值随着处理温度的升高逐渐上升。高温敏感的小麦品种/在T2 处理下的上升幅度显著大于高温不敏感的小麦品种。

图5 高温对高温不敏感型(G1)和高温敏感型(G2)小麦品种旗叶延迟荧光诱导曲线(0.01~0.02 ms)(A 和B)和延迟荧光参数I2/I1(C)的影响Fig.5 Effect of high temperature on the delayed fluorescence induction curve(0.01-0.02 ms)(A and B)and delayed fluorescence parameter I2/I1(C)of wheat flag leaves of high-temperature insensitive(G1)and sensitive(G2)cultivars
3 讨论
我国拥有丰富的小麦品种资源,不同小麦品种对高温胁迫的敏感程度也存在差异。如表1所示,本试验中所选用的小麦品种在高温下/均呈下降趋势,表明高温胁迫下所有小麦品种PSⅡ活性均下降,但是下降幅度显著不同。为了进一步研究高温对光合机构的影响机理,根据K-均值聚类将35 个小麦品种分为高温不敏感和高温敏感两种类型,对比分析两种类型小麦品种高温胁迫下光合电子传递过程的变化规律。
供体侧和受体侧活性是影响植物光系统Ⅱ活性的重要因素,OJIP 曲线的J 点荧光的变化可以反映P680 受体侧Q到Q的电子传递活性。在本试验中,高温处理下J 点荧光上升说明Q和Q之间的电子传递受到了抑制,且抑制程度随着处理温度的升高而增大。I 点的出现主要是由于还原状态的PQ(PQH)再氧化成为限制因素所导致的。在本试验中,高温胁迫导致的I 点荧光升高表明PQH的再氧化过程受到了抑制。高温敏感型小麦品种Q和Q之间电子传递活性以及PQ 库的再氧化过程在高温下的受抑制程度均显著大于高温不敏感小麦品种,表明Q和Q之间电子传递活性以及PQ 库的再氧化活性是决定小麦品种高温抗性的重要因素。
K 点荧光的变化通常用来反映PSⅡ供体侧的活性,虽然有研究表明高温、水淹等多种逆境胁迫均会导致K 点荧光的上升,但是近年来对于K 点反映供体侧伤害程度的可靠性也出现了不同的观点,K 点变化反映的是K 点的相对变化,因此可能会受到PSⅡ受体侧活性的影响,当PSⅡ受体侧活性伤害程度大于供体侧活性时则会掩盖供体侧活性的变化。金立桥等通过黑暗高温抑制供体测活性的同时使用D1 蛋白抑制剂、不同时间强光处理等获得不同PSⅡ受体侧伤害程度的叶片,发现受体侧的伤害程度严重影响K 点荧光的相对变化,表明仅在PSⅡ供体侧伤害程度大于受体侧伤害程度时才能观测到K 点相对荧光的显著上升。在本试验中,无论高温敏感型还是高温不敏感型的小麦品种,随着处理温度的上升,K 点荧光均没有上升,我们推测这种变化可能也是PSⅡ受体侧活性逐渐加重所导致的。Li 等在研究大豆()叶片对高温的响应时也观测到在短时间高温处理条件下K 点荧光下降的现象,印证了我们的推测。有研究表明小麦叶片放氧复合体对高温有较强的适应能力。在本试验中最高处理温度38 ℃也在冬小麦灌浆期的气温变化区间内,因此,38 ℃的高温可能不足以对大多数小麦品种旗叶PSⅡ的供体侧造成伤害。此外,光照是影响放氧复合体活性的重要因素,前人的试验往往在强光下进行,而本试验在弱光下进行,弱光环境可能也是导致K 点没有显著上升的原因之一。L 点的荧光反映了PSⅡ单元之间的能量偶联程度,在T2处理下,L 点荧光显著上升,且高温敏感型小麦品种L 点荧光上升幅度更大,表明了高温敏感型小麦品种高温胁迫下PSⅡ单元之间偶联程度下降程度更大,从而可能进一步影响PSⅡ对光能的利用效率。本试验中,和在高温胁迫下均显著下降,表明高温胁迫下,吸收的光能被PSⅡ反应中心捕获的效率、用来推动电子传递到Q下游电子载体以及用来推动电子传递到PSⅠ受体侧的效率均下降,且高温敏感型小麦品种更为敏感。表示传递到PSⅠ受体侧的电子占传递到Q下游的电子的比率,在本试验中随着处理温度的升高逐渐增大,表明高温胁迫下传递到PSⅠ受体侧的电子占传递到Q下游的电子的比率上升,由于在高温下是下降的,因此,可以推断的下降主要是由于电子传递到Q下游的比率减少所导致的,而不是PSⅠ的电子传递效率下降引起的。PI是反映PSⅡ活性的综合性能指数,逆境对光能捕获和传递的影响均能体现在PI的变化上。本试验中PI显著下降,也再次印证了高温对PSⅡ光能吸收和传递的抑制作用。
PSⅠ吸收光能后通过P700 不断氧化和再还原将供体侧的电子传递到受体侧,从而维持光系统Ⅱ的电子传递和整个光合电子传递链的运转。通过对叶片820 nm 光反射的监测,可以方便地研究PSⅠ的氧化和再还原的过程,进而分析PSⅠ以及PSⅡ电子传递能力的变化。无论高温敏感还是不敏感的小麦类型,在高温处理下,MR信号的最大氧化速率在各个处理之间没有显著差异,表明PSⅠ不是高温胁迫的主要伤害位点,Sun 等的研究也得到了类似的结论,印证了我们的观点。随着照光时间的延长,从PSⅡ传来的大量的电子超过了PSⅠ向下游传递电子的能力,从而导致了PSⅠ的再还原,再还原过程的最大斜率显著下降,且高温敏感型小麦下降幅度更大,表明PSⅡ向PSⅠ的电子传递受到高温的显著抑制,也印证了快速叶绿素荧光的结果。
延迟荧光是电子传递过程中的可逆反应产生的,当PSⅡ的供体侧或受体侧的电子传递体与P680发生电荷重组时,就会导致P680 的再激发和延迟荧光的产生,因此,延迟荧光主要反映了电子传递链P680 与其供体侧和受体侧的电荷状态。I点延迟荧光主要与SZP680Q-基团的相对浓度有关,主要受有活性P680 的数量以及供体和受体侧电子传递能力的影响。高温处理下的下降表明PSⅡ反应中心活性、供体侧活性和受体侧活性可能均受到抑制,而结合瞬时荧光K 点、和RC/CS的变化则进一步印证了高温对反应中心和受体侧电子传递活性的抑制。I的出现对应于快速叶绿素荧光曲线的I-P 段,I的出现是由于PQH向PSⅠ受体侧的电子传递导致了Q-的再氧化和PSⅡ反应中心的重新开放,因此,/可以用来反映PSⅠ供体侧电子传递到受体侧的效率。在本试验中,38 ℃高温处理下/显著提高,表明了电子传递到PSⅠ末端的效率提高,且高温敏感型小麦上升幅度更大,该结果与快速荧光的变化相一致。
4 结论
本研究根据不同小麦品种旗叶在高温下/的下降程度将35 个小麦品种分为高温不敏感和高温敏感的两种类型,然后采用快速叶绿素荧光诱导动力学曲线、820 nm 光反射和延迟荧光同步测定的方法研究了两种类型小麦品种的旗叶光合机构对高温胁迫响应特征。结果显示高温胁迫下,高温敏感型小麦品种PSⅡ光能捕获和传递到Q下游的能力与高温不敏感的小麦品种相比下降更加明显,并进一步导致光合作用综合性能的快速下降,Q受体侧电子传递到PSⅠ末端的效率和放氧复合体活性并没有显著下降,通过820 nm 光反射和延迟荧光结果的进一步印证,最终表明PSⅡ反应中心活性、PSⅡ光能的捕获和Q向下游传递电子的能力的差异是导致两种类型小麦品种光合电子传递活性差异的主要原因,PSⅡ供体侧和PSⅠ活性对小麦PSⅡ高温抗性没有直接影响。
猜你喜欢
作物学报(2022年9期)2022-07-18 00:58:42
干旱地区农业研究(2021年6期)2021-12-08 07:52:22
今日农业(2021年17期)2021-11-26 23:38:44
阅读(科学探秘)(2020年8期)2020-11-06 06:22:48
浙江农业科学(2019年11期)2019-11-09 01:37:48
中国果业信息(2019年1期)2019-01-05 17:41:42
生物学教学(2017年9期)2017-08-20 13:22:32
当代化工研究(2016年7期)2016-03-20 16:21:50
实用器官移植电子杂志(2015年1期)2015-04-02 14:58:26
郑州大学学报(医学版)(2015年2期)2015-02-27 14:50:44
