
藏北两种牧草添加氮磷后叶片N∶P计量学特征及重吸收率
2022-03-05 06:46洪江涛王小丹
草业科学 2022年1期
徐 昕,洪江涛,王小丹
(1.中国科学院成都山地灾害与环境研究所,四川 成都 610044;2.申扎高寒草原与湿地生态系统观测试验站,西藏 那曲 853100;3.中国科学院大学,北京 100049)
生态化学计量学是探究生态系统多种化学元素平衡的一门学科[1]。该科学将分子-细胞-有机体-种群-生态系统等多尺度生态学理论有机统一起来,在厘清植物养分利用效率、判定限制性养分和探究生物地球化学循环等多个方面发挥着至关重要的作用[2]。氮(nitrogen,N)、磷(phosphorus,P)是构成植物细胞和器官的基本元素,也是影响植物生长、发育和繁殖的关键养分[3]。
植物叶片N∶P 计量学特征是衡量N、P 资源限制类型的重要指标之一,因其简便性被广泛应用到各类生态系统植物受限元素判定研究中[4-5],甚至可以代替传统的野外养分添加试验[6]。Koerselman 和Meuleman[7]通过对欧洲湿地施肥试验的汇总分析,认为植被叶片N∶P 小于14 时该生态系统主要物种受N 限制,N∶P 大于16 时该生态系统主要物种受P 限制,而N∶P 介于14 与16 之间被N 和P 共同限制或者都不受限。此外,N、P 养分重吸收是指养分从衰落组织转移到其他新组织(新叶、茎、根系等)的过程,是植物养分保存的重要环节[8-9]。该指标可以反映物种养分利用效率,是研究植物生态化学计量学与外界环境因子变化和生态系统稳定性相互关系的重要桥梁[10]。在某种特定养分受限环境中,植物往往会提高该元素重吸收效率以适应贫瘠养分环境。例如:热带地区土壤淋溶作用较强,P 更易损失,植物具备较高的P 重吸收效率以应对土壤P 的匮乏[10]。此外,植物在应对养分限制时,还会激发出特殊的生理生态机制以适应胁迫环境。低N∶P(N 限制)的环境下植物会减缓叶片生长,提高植物根系生物量分配以捕获N;而高N∶P (P 限制)环境下植物会刺激细根周转、增加根际分泌物以提高P 吸收效率[11-14]。
青稞(Hordeum vulgare)是青藏高原主要的粮食作物,主要分布在高原河谷农业地带[15]。藏北羌塘高原平均海拔大于 4 500 m,积温低,青稞不易结穗,种植用途主要为青贮饲料。垂穗披碱草(Elymus nutans)具有营养丰富、适口性好和产量高等特点,是青藏高原上重要的栽培牧草之一[1]。青稞和垂穗披碱草生态适应性广,具有耐寒和抗旱等优点,在保护生态环境和植被恢复方面具有重要意义。同时也是西藏乡土禾草种质资源重点开发利用的对象,可以为农牧区牲畜提供优良饲草。藏北由于气候严寒、降水稀少以及土壤养分贫瘠,研究土壤N、P 添加后牧草叶片营养特征的响应对理解极端自然环境下牧草生理生态适应机制和养分利用效率具有重要意义。目前,关于垂穗披碱草和青稞抗寒[16-17]、抗旱[18-20]及耐盐碱[21-22]等方面特性的研究较多,而对土壤N、P 添加响应的报道较为少见。
本研究以藏北典型牧草青稞和垂穗披碱草为研究对象,通过养分配比、温室培养、室内分析等试验方法,分析两种牧草绿叶和枯叶N 浓度、P 浓度、N∶P比值以及重吸收效率的差异性,探究栽培草地牧草叶片N∶P 计量学特征对土壤N∶P 养分输入水平和梯度的响应,为明确典型牧草对梯度养分的利用策略提供科学依据。
1 材料与方法
1.1 试验方法
试验于2017年7月 -10月在藏北申扎高寒草原与湿地生态系统观测试验站开展(30°57′N,88°42′E,海拔4 700 m)。取当地典型高寒草原土壤(0 -30 cm)过2 mm 筛,拌入适量腐熟有机肥,然后装入塑料花盆中(高15 cm、底径11 cm、口径15 cm)用于种子萌发。每盆放入1.5 kg 高寒草原土并播种100 粒成熟饱满青稞或垂穗披碱草种子。每个物种幼苗在3 叶龄时进行移栽。移栽时确保每个幼苗高度、茎粗和叶片数量基本一致。移栽花盆土壤也取自申扎典型高寒草原表层0 -30 cm 土壤(表层土壤中砂粒、粉粒、粘粒含量分别为91%、7%和2%,容重为1.72 g·cm-3,其中细土(<2 mm)容重为1.52 g·cm-3有机碳含量为8.80 g·kg-1,总氮为1 g·kg-1,总磷为0.6 g·kg-1)[23],过筛后填充约1.5 kg 进入花盆。每个花盆移植一株青稞或垂穗披碱草幼苗于花盆中央,由于本试验中设计有18 个不同养分梯度及N∶P 输入水平的交互处理,每个处理均有3 次重复,因此在试验前期移栽过程中每个处理的实际移栽盆数为4~5 盆,每盆移栽一株,以确保每盆为独立生长,以达到独立重复试验要求,且保证有3 株植物存活供试验采样使用。移植后每天浇水保证幼苗成活和快速定植。幼苗成活后,为避免水分胁迫,每隔1~2 d 浇水一次。N 和P 添加试验在幼苗移植成活半个月后进行。N 添加采用NH4NO3,P 添加为KH2PO4。试验设置6 个N∶P 输入水平,分别为0.5、1.5、4.5、13.5、40.5、121.5。每个N∶P 输入水平均设置高、中、低3 个养分梯度,每种试剂元素折合等量纯N 或P 质量计算(表1)。N∶P 配比由供应近似平衡状态(13.5)到N 供应的极度失衡(0.5)及P 供应的极度失衡(121.5),由此来说明青稞及垂穗披碱草N∶P 供应由平衡到极度失衡状态下叶片N、P 含量及其N∶P 化学计量学特征和重吸收效率的变化特征,而绝对N、P 浓度设置为中梯度是低梯度的两倍,高梯度是中梯度的两倍,这样能使N∶P 供应效应的评价独立于绝对养分供应差异的影响,依据植物养分需求和生长周期,分3 次添加N、P 营养,第1 次添加总量的1/6(7月15日),第2 次添加总量的1/3 (7月31日),第3 次添加总量的1/2 (8月15日)[13]。每次添加时,试剂均溶于50 mL 水,再施入花盆。为防止其他元素缺乏,每个月添加一次K (400 mg)、Ca (133 mg)、Mg(40 mg)、Fe (14 mg) 和Zn (0.3 mg)等微量元素。植物均在自然光照下进行生长。绿叶和枯叶样品采集分别在植物生长旺季(9月10日)和生长季末(10月15日)进行,共计216 盆植物样品(6 个N∶P 输入水平×3 个养分梯度×2 个采样时间×2 个物种×3 重复)。

表1 N∶P 输入水平和养分梯度对牧草叶片养分特征的影响Table 1 Effect of N∶P supply ration and nutrient gradient on the nutrient characteristics of forage leaves
1.2 指标测定与方法
植物叶片在采集后用蒸馏水清洗干净,置于信封内用烘箱烘干。65 ℃烘72 h 至恒重,然后研磨成粉末进行分析测试。植物叶片N 浓度采用凯氏定氮法测定,P 浓度采用钼锑抗比色法测定[24]。
1.3 数据处理与分析
养分重吸收效率计算公式为:NuRE=(1 -Nu枯叶/Nu绿叶)×100%。式中:Nu枯叶和Nu绿叶表示枯叶和绿叶的养分浓度[25]。N∶P 输入水平和养分梯度对垂穗披碱草和青稞叶片的N 和P 浓度、N∶P、N 和P 重吸收效率以及P重吸收效率∶N重吸收效率利用方差分析(ANOVA)进行分析。两个物种的N 和P 重吸收效率相互关系利用线性回归进行分析。数据在SPSS 16.0 中进行分析,统计图绘制在Sigmaplot 11.0中进行。
2 结果与分析
2.1 绿叶N∶P 计量学特征
青稞绿叶N 浓度随N∶P 输入水平提高而极显著增加(P<0.01) (图1a 和表2),其中N∶P 输入水平从0.5 提高到121.5,绿叶N 浓度平均值从18.34 mg·g-1增加到28.56 mg·g-1(图1a)。中高梯度下,青稞绿叶P 浓度随N∶P 输入水平提高而下降,但在N∶P输入水平为121.5 时P 浓度有所提升(图1b)。低梯度下青稞绿叶P 浓度随N∶P 输入水平先升高后降低再升高的趋势(图1b)。高梯度下,青稞绿叶N∶P在N∶P 输入水平为40.5 时达到最高值,低梯度下绿叶N∶P 随N∶P 输入水平先降低后升高再降低的趋势(图1c)。养分梯度及其与N∶P 输入水平双因子交互作用对青稞绿叶P 浓度和N∶P 有极显著影响(P<0.01),而对N浓度无显著影响(P>0.05)(图1 和表2)。高、中、低养分梯度下青稞绿叶P 浓度平均值为1.46、1.50 和1.16 mg·g-1,而N∶P 平均值分别为18.91、17.96 和22.57 (图1)。

表2 N∶P 输入水平和养分梯度对牧草叶片养分特征的影响(F 值)Table 2 Effect of supply ration and nutrient gradient on the nutrient characteristics of forage leaves (F value)

图1 N∶P 输入水平和梯度对两种牧草绿叶N∶P 计量学特征的影响Figure 1 Influence of N∶P supply ration and gradient on the N∶P metrological characteristics of two forage green leaves
垂穗披碱草绿叶N 浓度随N∶P 输入水平提高有极显著提升(P<0.01) (图1d),其中N∶P 输入水平从0.5 提高到121.5,绿叶N 浓度平均值从24.63 mg·g-1增加到31.69 mg·g-1(图1d)。高梯度下,垂穗披碱草绿叶P 浓度随N∶P 输入水平呈现先下降后上升的趋势,在N∶P 输入水平为40.5 时最低(图1e)。垂穗披碱草绿叶N∶P 随N∶P 输入水平提高总体呈现上升趋势(图1f)。养分梯度及其与N∶P 输入水平双因子交互作用对垂穗披碱草绿叶P 浓度、N∶P 有极显著影响(P<0.01) (图1 和表2)。高、中、低养分梯度下垂穗披碱草绿叶P 浓度平均值为2.17、1.84 和1.64 mg·g-1,而N∶P 比平均值分别为13.4、15.78 和17.18。
2.2 枯叶N∶P 计量学特征
青稞枯叶N 浓度随N∶P 输入水平提高总体呈现波动式上升趋势,而P 浓度呈现先下降后上升趋势,在N∶P 输入水平为40.5 时最低(均值为0.44 mg·g-1)(图2a,b)。青稞枯叶N∶P 随N∶P 输入水平提高总体呈上升趋势(图2c)。养分梯度对青稞枯叶N 浓度、P 浓度和N∶P 均无显著影响(P>0.05) (表2)。养分梯度与N∶P 输入水平交互作用对青稞枯叶N 浓度、P 浓度和N∶P 均有显著影响(P<0.05) (表2和 图2a,b,c)。

图2 N∶P 输入水平和梯度对两种牧草枯叶N∶P 计量学特征的影响Figure 2 Influence of N∶P supply ration and gradient on the N∶P metrological characteristics of two forage dead leaves
垂穗披碱草枯叶N 浓度随N∶P 输入水平提高变化不显著(图2d),在高梯度下P 浓度总体呈现先下降后上升的趋势,低、中梯度下表现为现下降后上升最后再下降的趋势(图2e)。垂穗披碱草枯叶N∶P随N∶P 输入水平提高低、中梯度呈现先上升后下降的趋势,高梯度则表现为波动上升趋势(图2f)。养分梯度仅对垂穗披碱草枯叶N 浓度有极显著影响(P<0.01) (表2),高、中、低梯度平均值分别为15.2、16.69 和16.79 mg·g-1。养分梯度与N∶P 输入水平交互作用对垂穗披碱草枯叶N 浓度、P 浓度和N∶P 均有显著影响(P<0.05) (表2 和 图2d,e,f)。
2.3 N、P 元素重吸收效率
青稞N 重吸收效率随N∶P 输入水平提高呈现先升高后下降的趋势(图3a),在N∶P 输入水平为13.5 时达 到最大 值(均 值 为46.01 mg g-1)。而N∶P输入水平对P 重吸收效率无显著影响(P>0.05) (表2和图3b)。青稞P 重吸收效率∶N 重吸收效率随N∶P输入水平提高呈现先下降后上升的趋势(图3c)。养分梯度对青稞N 重吸收效率无显著影响(P>0.05) (表2),但对P 重吸收效率以及P 重吸收效率∶N 重吸收效率有极显著影响(P<0.01) (表2)。高、中、低养分梯度P 重吸收效率分别为43%、56%和57%。青稞P 重吸收效率∶N 重吸收效率比均值为1.93,表现为P 重吸收效率高于N 重吸收效率。养分梯度与N∶P 输入水平交互作用对青稞N 重吸收效率、P 重吸收效率及P 重吸收效率∶N 重吸收效率均有显著影响(P<0.05) (表2 和图2a,b,c)。
垂穗披碱草N 重吸收效率平均值由N∶P 输入水平为0.5 时的29.88%提高到121.5 时的51.40%(图3d)。垂穗披碱草P 重吸收效率总体上呈现先上升后下降的趋势(图3e)。高梯度下垂穗披碱草P 重吸收效率∶N 重吸收效率在4.5 时最高(1.38) (图3f)。养分梯度及其与N∶P 输入水平双因子交互作用对N 重吸收效率、P 重吸收效率及P 重吸收效率∶N 重吸收效率均有极显著影响(P<0.01) (表2 和图3)。垂穗披碱草P 重吸收效率∶N 重吸收效率比均值为0.61,表现为P 重吸收效率低于N 重吸收效率。
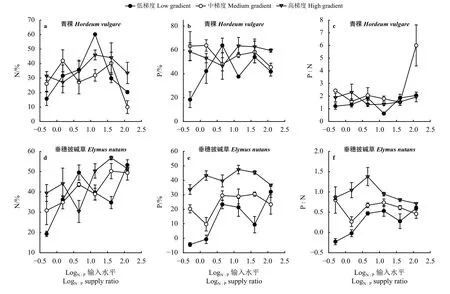
图3 N :P 输入水平和梯度对两种牧草绿叶N、P 重吸收效率的影响Figure 3 Effect of N :P supply ration and gradient on the reabsorption efficiency of N and P of two forage green leaves
3 讨论
青稞和垂穗披碱草绿叶N 浓度随N∶P 输入水平的提高均显著增加,可能与高N∶P 输入水平显著提升土壤活性N 含量,增加土壤N 的可利用性有关,促使植物从土壤中吸收更多的N[3]。N∶P 输入水平的提高总体会降低植物绿叶P 浓度,表现出一定的P 限制性。主要原因可能是大量的N 输入导致生物量显著提升,而P 吸收增幅有限产生“稀释效应”[26]。该结果与研究者[27-29]在高寒草甸、中国东部沿海杨树人工林、中国北方草原开展的研究结果一致。总体而言,养分梯度及其与N∶P 输入水平双因子交互作用对青稞和垂穗披碱草N 浓度影响不明显,而对P 浓度影响较大。本研究中,高养分梯度下N∶P 输入水平为121.5 时青稞和垂穗披碱草绿叶P 浓度均有一定提升,原因可能是植物为了避免N 和P 元素失衡,触发P 限制下的特殊生理调控与适应机制。Marklein 和Houlton[30]对多个陆地生态系统进行Meta 分析发现,随着土壤N 水平的提高,植物能够活化微生物(例如菌根活动)[31]或根际P 酸酶[30,32]等的活性,从而提高土壤中速效P 和植物叶片中P 含量,缓解N 水平过高而引起生态系统可能面临受P 限制的状态。自然状态下,植物叶片中N 与 P 浓度是相互耦合的,但土壤养分变化会改变植物组织N∶P 比以及体内N、P 元素平衡。根据 Koerselman 和Meuleman[7]养分限制理论与界限判定值,青稞在N∶P 输入水平为0.5,中高梯度养分输入时,N 限制作用较强(绿叶N∶P<14)。而随着N∶P 输入水平的提高,N 限制逐渐缓解,N∶P 逐步提高,植物趋向于P 限制(绿叶N∶P>16)。垂穗披碱草在N∶P 输入水平为0.5~4.5,高梯度养分输入时,N 限制作用最强(绿叶N∶P<14)。这与呼伦贝尔草地研究结果基本一致,随着N 输入水平的提高,除披针叶黄华(Thermopsislanceolata)外,其他物种叶片N∶P 均大于16,说明N 添加能够缓解呼伦贝尔草地优势物种的N 限制[33]。
枯叶中养分含量是表征植物养分重吸收程度的指标之一,不受绿叶养分含量的影响,能更为客观地反映养分再利用状况,枯叶中养分含量与植物重吸收程度呈反比关系,即养分含量越高,则植物对此种养分的重吸收程度越低[34]。在温带草原生态系统的研究[35]发现,N 添加提高了植物枯叶的N 浓度和N∶P,但对P 浓度没有显著影响。对一个半干旱温带草原的N 施加试验[36]发现,N 水平提高对黄囊苔草(Carex korshinskyi)枯叶的N 浓度和P 浓度影响不显著,而N 水平提高会增加羽茅(Achnatherum sibiricum)枯叶N 浓度,以及降低其枯叶P 浓度。本研究发现青稞枯叶N 浓度随着N∶P 输入水平的提高而增加,说明青稞在N∶P 输入水平较高时N 再利用程度较低。而垂穗披碱草N 浓度变化随着N∶P 输入水平的提高不显著。青稞和垂穗披碱草枯叶N 浓度对N∶P 输入水平响应的差异,表明养分再利用状况存在种间差异,主要取决于N 供应和植物需求平衡的结果[37]。青稞和垂穗披碱草P 浓度随N∶P 输入水平提高均呈现先下降后上升趋势。两个物种枯叶P 浓度随N∶P 输入水平呈现非线性响应,表明N∶P 输入水平为13.5 和40.5 时P 再利用程度最高。
营养元素的养分重吸收过程是植物保存养分的重要机制,可以使 N、P 元素从衰老的植物器官转移到新的器官,提高养分在植物体内存留时间,为后续生长繁殖提供能量和支持[8-9]。除此之外,养分重吸收过程还可以降低由生境地养分供给不足或者存在波动所带来的风险,有效缓解其因生长、繁殖和代谢需要大量养分所带来的压力,降低植物对外界环境的依赖性。对全球尺度内的植物养分重吸收进行了整合分析[10]发现,N 重吸收效率均值为46.9%,而P 重吸收效率均值为53.5%。本研究中,青稞和垂穗披碱草的N 和P 重吸收效率分别为37%和39%,略低于全球均值,这可能是由于本研究人为添加了N、P 元素,植物可以从土壤中直接获取大量养分,从而削弱了植物养分重吸收效率,印证了植物在高养分供给时存在“奢侈吸收”策略[38],该结果也在苔原[39]、沼泽[40]、森林[41]生态系统中得到验证。本研究还发现高N∶P 输入水平,青稞N 重吸收效率均呈现出下降趋势,但垂穗披碱草的N 重吸收效率均呈升高趋势。这种差异化响应主要由物种属性决定,反映了植物多元化的养分利用和分配策略。N 输入水平提高可以有效缓解N 对青稞的限制作用,但垂穗披碱草N 利用策略更为保守,即便是在高N 输入环境下,该物种仍会逐步提高N 重吸收速率以满足其对N 的需求,预示着该物种在该地区受N 限制作用更强。青稞P 重吸收效率显著高于垂穗披碱草,表明青稞P 的利用策略更为保守,在藏北更易受到P 限制
本研究中N∶P 输入水平和养分梯度跨度较大,但叶片N、P 重吸收效率显示出很强的耦合性,这与内蒙古温带草原的研究一致,N 输入水平提高情况下,优势物种羊草(Leymus chinensis)和大针茅(Stipa grandis) N 和P 重吸收速率均表现出一致的变化特征[42]。结果表明,虽然土壤养分是调控植物N、P 动态的重要因子,但是有机体内N、P 元素平衡是高等植物普遍规律,体现了植物叶片属性间经济谱策略[43]。未来气候变化背景下,环境介质养分输入配比可能存在失衡(例如N 沉降加剧),但高寒生态系统中典型牧草N 和P 的吸收、转化和利用显示出动态耦合的现象,表现出较强的养分适应性。
4 结论
N、P 元素输入后青稞和垂穗披碱草叶片的养分特性变化可以反映其生长和养分限制情况。两个物种绿叶和枯叶对N∶P 输入水平和梯度变化呈现不同的响应趋势,这种差异化变化可能由物种属性决定,反映了植物多元化的养分利用和分配策略。青稞和垂穗披碱草重吸收效率P∶N 均值分别为1.93和0.61,表明青稞的P 素利用策略更为保守,而垂穗披碱草的N 素利用策略更为保守。植物体内的N、P 计量学特征受土壤养分供给和物种养分利用策略的共同影响,但本研究并未测定植物生物量和土壤养分含量,在指导栽培草地建植方面还存在一定的局限性,下一步需要继续探究土壤养分调控与牧草生产力之间的关系,以期为建立栽培草地高效养分管理和营养调控技术提供科学支撑。
猜你喜欢
九江学院学报(自然科学版)(2022年2期)2022-07-02
星星·诗歌原创(2022年1期)2022-04-16
华人时刊(2021年17期)2021-12-02
西藏人文地理(2020年4期)2020-11-19
创新作文(5-6年级)(2019年10期)2019-12-23
翠苑(2019年4期)2019-11-11
华东师范大学学报(自然科学版)(2019年3期)2019-06-24
福建基础教育研究(2019年3期)2019-05-28
西部资源(2018年1期)2018-11-01
娃娃画报(2018年3期)2018-08-15
