
Hippo/YAP通路在大鼠脑动脉瘤形成过程中的作用及机制研究
2022-03-04 07:25王宏国石海亮
微循环学杂志 2022年1期
王宏国 熊 刚 石海亮
脑动脉瘤(Cerebral Aneurysm, CA)是一种致命的神经血管疾病,由多种因素引起,包括慢性炎症、血流动力学应激和血管壁重塑[1]。内皮损伤可由血流动力学应激的变化引起,导致促炎细胞因子分泌上调,随后募集各种炎性细胞。炎性细胞被吸引到内皮损伤部位,这种募集和随后的浸润诱导平滑肌细胞增殖、凋亡和动脉壁的重塑,促进动脉瘤的形成和破裂[2,3]。白细胞,特别是巨噬细胞,渗入血管壁被认为是CA形成和破裂的关键步骤;巨噬细胞已被确定为在CA中协调炎症反应的关键效应细胞。近年来,已提出失衡的巨噬细胞M1/M2极化参与CA的进展[4,5]。在CA形成中促炎M1/M2比率随时间延长而增加[6];此外,在破裂的CA中M1细胞显著高于M2细胞,针对巨噬细胞激活或防止M1/M2失衡的疗法可阻止CA的形成和破裂[7]。
Hippo通路在炎症和脑血管疾病中具有重要作用。Yes相关蛋白(Yes-associated Protein, YAP)是Hippo信号通路下游的关键效应因子,在中枢神经系统的生理和病理发展中发挥重要作用,尤其是可以调节细胞增殖、分化、迁移和凋亡。研究已经证实YAP的异常上调或核定位发生在许多人类恶性肿瘤中,并促进它们的形成、进展和转移[8,9]。然而,Hippo/YAP通路在中枢神经系统疾病中的作用和潜在机制仍不明确。据报道,Hippo信号通路关键蛋白YAP可调控巨噬细胞极化,在炎症性肠病中,YAP的过度表达会损害白细胞介素(Interleukin,IL)-4/IL-13诱导的M2巨噬细胞极化,同时促进脂多糖(LPS)/干扰素-γ(IFN-γ)触发的M1巨噬细胞激活以产生IL-6;敲低其表达可促进M2巨噬细胞极化[10]。在机械通气诱导的肺损伤和肺部炎症中,巨噬细胞YAP的表达上调,YAP缺乏会增强M2极化,同时抑制M1极化[11]。而Hippo/YAP通路是否参与CA的形成和进展还未可知。因此,本研究旨在探究Hippo/YAP通路在CA形成中的作用及其可能的分子机制,为CA的防治提供参考。
1 材料与方法
1.1 实验动物
SPF级雄性SD大鼠,体重(200±20)g,购自北京维通利华实验动物技术有限公司,许可证为SCXK(京)2019-0009。饲养在温度(20±2)℃、湿度(45-55)%、12h光照/12h黑暗周期的条件下,可自由饮食和摄水。实验经本院伦理委员会批准,并按实验动物使用的3R原则进行。
1.2 试剂及仪器
维替泊芬(Verteporfin,YAP抑制剂,可破坏YAP-TEAD相互作用,HY-B0146)购自美国MedChemExpress(MCE)公司;HE染色试剂(C0105)、RIPA裂解液(P0013B)、BCA试剂盒(P0010)均购自碧云天生物科技公司;大鼠IL-6(ml102828)、肿瘤坏死因子-α(Tumor Necrosis Factor-α,TNF-α)(ml002859)、IL-10(ml002813)、血管内皮生长因子(Vascular Endothelial Growth Factor,VEGF)(ml002862)、内皮素-1(Endothelin-1,ET-1)(ml002890)、单核细胞趋化蛋白-1(Monocyte Chemotactic Protein-1, MCP-1)(ml002960)ELISA检测试剂盒均购自上海酶联生物科技有限公司;小鼠抗大鼠F4/80抗体(sc-377009)购自Santa Cruz Biotechnology;兔抗大鼠诱导型一氧化氮合酶(Inducible Nitric Oxide Synthase,iNOS)(ab178945)、兔抗大鼠精氨酸酶-1(Argininase 1,Arg-1)(ab96183)、山羊抗小鼠Alexa Fluor 594(ab150116)、山羊抗兔Alexa Fluor®488(ab150077)、兔抗大鼠Histone H3(ab1791)均购自英国abcam公司;CelLyticTMNuCLEARTM提取试剂盒(NXTRACT)购自Sigma-Aldrich公司;兔源一抗哺乳动物Ste-20样激酶(Mammalian Sterile 20-like Kinase, MST)1/2(PA5-36100)、Phospho-MST1/2(p-MST1/2,PA5-104616)、YAP(PA5-17609)、大型肿瘤抑制因子(Large Tumor Suppressor, LATS)1/2(PA5-115498)、Phospho-LATS1/2(p-LATS1/2,PA5-105895)、β-actin(MA5-32479)、HRP标记的山羊抗兔IgG(H+L)(65-6120)均购自Thermo Fisher Scientific公司。iMark680多功能酶标仪、蛋白转膜装置(美国Bio-Rad公司);IX71倒置荧光显微镜(日本Olympus公司);EZ4HD高清数码解剖显微镜(德国Leica公司)。
1.3 实验方法
1.3.1 模型制备及分组:大鼠随机分为对照组(n=10)、模型组(n=30)和YAP抑制剂组(n=10),模型组根据样本采集时间分为A、B、C共3个亚组,每个亚组10只。模型组和YAP抑制剂组大鼠采用结扎左颈总动脉和双侧肾动脉联合高盐饮食建立CA模型[12,13]:用3%戊巴比妥钠(30mg/kg)腹腔注射麻醉大鼠,脱去背部毛发,经背双侧肋缘下行纵切口,用丝线结扎双侧肾动脉后支。7天后,在手术显微镜下用丝线结扎左颈总动脉,并用1%盐水替代饮用水,建立大鼠肾性高血压模型。YAP抑制剂组大鼠每2天腹腔注射100mg/kg的Verteporfin[14]。对照组仅分离左颈总动脉和双肾动脉,不做结扎,其余操作同上,标准喂养。使用鼠尾袖套法测定大鼠在清醒状态下的血压值。分别于造模后1周、造模后1个月、造模后3个月三个时间点对A、B、C亚组大鼠进行取材和指标检测,对照组和YAP抑制剂组大鼠于造模后3个月进行取材。
1.3.2 ELISA检测血清炎性因子水平及血管内皮损伤标志物水平:大鼠麻醉后,腹主动脉取血3ml,静置后1 500r/min离心10min,分离血清,按照ELISA试剂盒说明书检测血清MCP-1、TNF-α、IL-6、IL-10和血管内皮损伤标志物VEGF、ET-1水平。
1.3.3 CA形态学评估:取血完成后,通过左心室灌注4%多聚甲醛处死大鼠。然后立即通过左心室灌注溶解在10%(w/v)明胶/PBS溶液中的溴酚蓝。收集大脑,光学显微镜检查Willis环是否有血管异常[6]。0级,Willis环血管正常;1级,血管迂曲、扩张,形态不规则,但没有可识别的动脉瘤;2级,单个囊状动脉瘤;3级,多个囊状动脉瘤;4级,破裂的囊状动脉瘤。
1.3.4 免疫荧光染色检测动脉瘤组织中巨噬细胞极化和浸润:收集有或没有动脉瘤的大鼠Willis环,用4%多聚甲醛固定24h后,将样品转移到18%蔗糖溶液中过夜,然后冷冻在OCT化合物中。用低温恒温器将所有样品切片(30μm),放在载玻片上,并在室温下干燥过夜。抗原热修复后,0.01M PBS静置水化20min,漂洗后用10%正常山羊血清封闭1h,小鼠抗大鼠F4/80、兔抗大鼠iNOS、兔抗大鼠Arg-1(按1∶100稀释)一抗孵育。再使用山羊抗小鼠Alexa Fluor 594、山羊抗兔Alexa Fluor®488二抗(按1∶200稀释)检测一抗。DAPI染核,固定。荧光显微镜观察巨噬细胞浸润动脉壁的情况。图像分析由不知情的观察者进行。M1巨噬细胞定义为F4/80+iNOS+,M2巨噬细胞定义为F4/80+Arg-1+细胞。
1.3.5 Western blotting检测动脉瘤血管组织Hippo/YAP通路相关蛋白表达:取有或没有动脉瘤的大鼠Willis环血管组织,剪碎后,使用RIPA裂解液提取组织中的蛋白质(用于检测MST1/2、p-MST1/2、LATS1/2、p-LATS1/2蛋白表达),并采用细胞核蛋白提取试剂盒提取细胞核中的蛋白质(用于检测YAP蛋白表达)。BCA法测定总蛋白浓度,上样,SDS-PAGE电泳分离,并转移到PVDF膜上。用5%脱脂牛奶在室温下封闭2h后,对应加入一抗(MST1/2、p-MST1/2、LATS1/2、p-LATS1/2、YAP、β-actin、Histone H3,按1∶1 000稀释)4℃孵育过夜。洗去一抗,加二抗(1∶2 000)室温下孵育1h,ECL显色,凝胶成像仪观察蛋白条带并拍照。以β-actin或Histone H3为内参蛋白,以目的蛋白与内参蛋白条带灰度值的比值表示目的蛋白的相对表达量。
1.4 统计学处理

2 结 果
2.1 各组大鼠血清血管内皮损伤标志物VEGF、ET-1水平
各组大鼠血清VEGF、ET-1水平差异有统计学意义(P<0.05)。与对照组相比,A、B、C组大鼠血清VEGF和EF-1水平均显著升高(t均>6.029,P<0.05);与A组相比,B组和C组大鼠血清VEGF、ET-1水平显著升高(t均>4.761,P<0.05);与B组相比,C组大鼠血清VEGF、ET-1水平显著升高(t均>13.607,P<0.05);与C组相比,YAP抑制剂组大鼠血清VEGF、ET-1水平显著降低(t=7.167、11.175,P<0.05)。见表1。

表1 各组大鼠血清VEGF、ET-1水平
2.2 各组大鼠血清MCP-1、TNF-α、IL-6、IL-10水平
各组大鼠血清MCP-1、TNF-α、IL-6、IL-10水平差异有统计学意义(P<0.05)。与对照组相比,A、B、C组大鼠血清MCP-1(t=5.634、11.999、14.132,P<0.05)、TNF-α(t=13.229、19.131、25.906,P<0.05)、IL-6(t=14.098、19.736、27.559,P<0.05)水平均显著升高,IL-10水平(t=6.888、13.125、19.607,P<0.05)显著降低;与A组相比,B组和C组大鼠血清MCP-1(t=7.119、9.567,P<0.05)、TNF-α(t=7.209、15.184,P<0.05)、IL-6(t=17.781、7.404,P<0.05)水平显著升高,IL-10水平(t=6.460、12.708,P<0.05)显著降低;与B组相比,C组大鼠血清MCP-1(t=2.635,P<0.05)、TNF-α(t=8.184,P<0.05)、IL-6(t=11.087,P<0.05)水平均显著升高,IL-10(t=5.553,P<0.05)水平显著降低;与C组相比,YAP抑制剂组大鼠血清MCP-1、TNF-α、IL-6水平均显著降低,IL-10水平显著升高(t=2.178、6.427、8.067、5.278,P<0.05)。见表2。
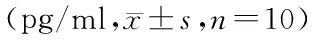
表2 各组大鼠血清MCP-1、TNF-α、IL-6、IL-10水平
2.3 各组大鼠CA形态学变化
对照组大鼠均无CA形成,A、B、C组大鼠可见动脉瘤形成,且动脉瘤级别依次增加;与C组相比,YAP抑制剂组大鼠动脉瘤形成级别显著降低。见图1和表3。

图1 各组大鼠CA形态学变化

表3 各组大鼠CA级别分布情况(%,n均=10)
2.4 各组大鼠CA中巨噬细胞极化和浸润情况
对照组大鼠的Willis环中未发现F4/80阳性细胞,无巨噬细胞浸润;与对照组相比,A、B、C组大鼠Willis环中出现大量F4/80阳性细胞,M1型巨噬细胞比例、M2型巨噬细胞比例显著升高(P<0.05);与A组相比,B组和C组M1型巨噬细胞比例(t=6.922、14.109,P<0.05)、M1/M2型巨噬细胞比值(t=12.017、28.185,P<0.05)显著增加,M2型巨噬细胞比例(t=5.057、10.110,P<0.05)显著降低;与B组相比,C组M1型巨噬细胞比例(t=8.210,P<0.05)、M1/M2型巨噬细胞比值(t=17.594,P<0.05)显著增加,M2型巨噬细胞比例(t=5.413,P<0.05)显著降低;与C组相比,YAP抑制剂组大鼠Willis环中M1型巨噬细胞比例、M1/M2型巨噬细胞比值显著降低(P<0.05),M2型巨噬细胞比例显著升高(t=6.590、14.353、3.963,P<0.05)。见图2和表4。

注:红色荧光,iNOS+(Arg-1+);绿色荧光,F4/80+;橙黄色荧光,双阳性

表4 各组大鼠CA中M1型、M2型巨噬细胞比例
2.5 各组大鼠动脉瘤血管组织Hippo/YAP通路相关蛋白表达
与对照组相比,A、B、C组大鼠动脉瘤血管组织p-MST1/2/MST1/2比值(t=4.462、8.535、12.738,P<0.05)、p-LATS1/2/LATS1/2比值(t=4.044、8.629、14.118,P<0.05)显著降低,胞核YAP的表达显著增加(t=4.445、7.894、14.033,P<0.05);与A组相比,B组和C组p-MST1/2/MST1/2比值(t=4.116、8.455,P<0.05)、p-LATS1/2/LATS1/2比值(t=5.433、12.474,P<0.05)显著降低,胞核YAP的表达显著升高(t=3.644、9.312,P<0.05);与B组相比,C组p-MST1/2/MST1/2比值(t=4.454,P<0.05)、p-LATS1/2/LATS1/2比值(t=7.589,P<0.05)显著降低,胞核YAP的表达(t=5.217,P<0.05)显著升高;与C组相比,YAP抑制剂组大鼠动脉瘤血管组织胞核YAP的表达显著降低(t=4.859,P<0.05),p-MST1/2/MST1/2和p-LATS1/2/LATS1/2比值差异无统计学意义(P>0.05)。见图3和表5。

图3 各组大鼠动脉瘤血管组织Hippo/YAP通路相关蛋白的表达(Western blotting)

表5 各组大鼠动脉瘤血管组织Hippo/YAP通路相关蛋白的表达
3 讨 论
巨噬细胞通常分为两个亚群,分别称为经典活化巨噬细胞(M1型)和交替活化巨噬细胞(M2型)。静息状态的巨噬细胞可识别LPS、TNF-α、IL-1β等向M1型巨噬细胞转化,高表达iNOS、CD11b和促炎细胞因子,M2巨噬细胞可以被IL-4、IL-10、TGF-β等诱导分化,高表达Arg-1、CD206和IL-10等[15]。通常,M1细胞表现出促炎作用,而M2细胞促进炎症消退并促进组织修复,它们的失衡被认为与各种疾病有关。CA发展的特征是向M1巨噬细胞表型的极化增加;源自M1细胞的炎性细胞因子引发动脉瘤壁的病理变化,尤其是TNF-α和IL-6。TNF-α是CA发病机制中的一种重要细胞因子,在未破裂和破裂的CA中升高,在CA形成和破裂中起关键作用;TNF-α抑制剂可有助于预防CA进展和破裂[16]。IL-6在人CA中表达,在雌激素缺乏小鼠血清中升高,可通过增强Willis环处的巨噬细胞浸润促进小鼠雌激素缺乏相关的CA破裂[4]。Nowicki等[6]发现在小鼠CA形成中M1/M2比率随时间的增加而逐渐升高,M1巨噬细胞是CA形成所必需的。血流动力学的改变、肾性血压升高与CA的形成关系密切。在本研究中,我们通过结扎大鼠左颈总动脉改变其颅内局部血流动力学,结扎双侧肾动脉后支以造成肾性高血压的方法来诱导建立CA大鼠模型。结果显示,随着时间的增加,大鼠颅底Willis环上可观察到从血管迂曲到有明显动脉瘤形成;并且在此过程中,血清TNF-α、IL-6、MCP-1和VEGF、ET-1水平升高,IL-10水平降低;通过免疫荧光双标法检测了动脉瘤中M1型(F4/80+iNOS+)和M2型(F4/80+Arg-1+)巨噬细胞的比例,结果显示,在CA形成和发展过程中,M1/M2型巨噬细胞比值显著增加,与以上研究结果一致。MCP-1可以诱导巨噬细胞浸润和粘附在动脉瘤的动脉壁上,促进CA的形成和发展[17]。以上结果说明巨噬细胞极化参与CA的形成和进展。Hippo信号通路最初是在果蝇中发现的,是一种保守的进化信号通路,参与组织发育和再生,通过调节细胞增殖和凋亡来控制器官大小[18]。近年来,因其在调节器官大小方面的显著作用以及与组织再生和癌症的明显相关性而迅速引起了人们的广泛关注。Hippo信号通路由核心激酶级联组成,其中上游激酶MST1/2磷酸化下游激酶LATS1/2,进而磷酸化和灭活促增殖转录共激活因子YAP和TAZ(具有PDZ结合基序的转录共激活因子),抑制YAP入核,通过与转录增强缔合域(TEAD)家族结合控制基因表达[19]。在生理上,Hippo通路充当肿瘤抑制因子,YAP是一种重要的转录共激活因子,受到Hippo信号通路的负调控,研究发现YAP/TAZ在人类癌症中被过度激活,参与细胞增殖、间充质转化、侵袭、转移形成,以及癌症干细胞维持和化学抗性[20,21];并且内皮YAP/TAZ过度激活可诱导过度的丝状伪足和增生的血管生长,参与病理性血管生成[22]。
最近的研究表明Hippo/YAP信号通路在中枢神经系统发育和脑肿瘤(包括神经胶质瘤)中起着重要作用[23,24]。有研究者对正常成人大脑以及264个脑肿瘤中核YAP的表达进行检测发现,在人脑肿瘤的细胞核中也很容易检测到YAP蛋白,高水平的YAP表达与神经胶质瘤的侵袭性分子亚群以及患者总生存期和无进展生存期相关[25,26]。Chen等[27]发现在颈动脉内膜损伤大鼠中,YAP及其许多下游靶基因的表达增加,可增强血管平滑肌细胞的增殖和迁移;通过激活Hippo信号通路,增强LATs的磷酸化,可抑制YAP入核,抑制血管损伤后的内膜增生;Xu等[28]研究也发现激活Hippo-YAP/TEAD1信号通路可防止氧化LDL诱导的内皮细胞功能障碍和炎症反应。更重要的是,研究发现巨噬细胞的极化受Hippo信号通路关键蛋白YAP的调控,抑制YAP可以降低巨噬细胞中iNOS、TNF-α、IL-6、IL-1β水平,减轻LPS刺激的巨噬细胞中的炎症反应[29]。Zhou等[10]发现YAP的过度表达会促进LPS/IFN-γ诱导的M1巨噬细胞激活以产生IL-6,同时会损害IL-4/IL-13诱导的M2巨噬细胞极化,加重炎症性肠病;Luo等[11]研究也发现特异性敲除巨噬细胞中的YAP,会增强M2极化,同时抑制M1极化,减轻机械通气后的肺损伤。本研究结果显示,在CA形成过程中,动脉瘤血管组织磷酸化MST1/2和磷酸化LATS1/2/的表达显著降低,胞核YAP的表达显著增加,说明Hippo信号通路被抑制;且胞核YAP的表达变化趋势与M1/M2型巨噬细胞比值趋势相反。使用YAP抑制剂Verteporfin干预可明显降低胞核YAP的表达,并降低M1型巨噬细胞比例及M1/M2型巨噬细胞比值,升高M2型巨噬细胞比例;提示,抑制YAP的活化可抑制巨噬细胞向M1型极化,同时促进其向M2极化。
综上所述,Hippo/YAP信号通路可能通过介导巨噬细胞极化参与CA的形成和进展。本研究仅从体内动物水平上初步探讨了Hippo/YAP在CA形成和进展中的作用机制,后续将考虑进行细胞实验,深入验证此研究结果。
◀
猜你喜欢
现代财经-天津财经大学学报(2022年5期)2022-06-01
航天电子对抗(2022年2期)2022-05-24
北京航空航天大学学报(2021年9期)2021-11-02
天津医科大学学报(2021年4期)2021-08-21
现代临床医学(2021年2期)2021-03-29
航天电子对抗(2019年4期)2019-06-02
广东教育·高中(2018年1期)2018-01-31
中学生数理化·教与学(2017年4期)2017-04-22
安徽医科大学学报(2015年9期)2015-12-16
新高考·高一物理(2015年4期)2015-08-20
