
缢蛏早期耐高温家系建立及抗氧化能力测定
2022-03-03 13:18申奔龙薛宝宝孟德龙沈和定
浙江农业学报 2022年2期
申奔龙,薛宝宝,孟德龙,沈和定
(上海海洋大学 水产种质资源发掘与利用教育部重点实验室,水产科学国家级实验教学示范中心,海洋动物系统分类与进化上海高校重点实验室,上海 201306)
缢蛏()是中国四大海水养殖贝类之一,养殖历史悠久、分布广泛,因其肉多壳薄、味道鲜美、营养丰富、医用价值等优点深受沿海居民的喜爱。
因近年来全球气候变暖且未来还将持续,这对海洋无脊椎动物尤其是潮间带滩涂贝类的生长、发育、免疫、生存等带来不同程度的影响。如栉孔扇贝()在高温下导致生理代谢失常、抗逆力下降,其免疫系统更易受病原的侵袭,最终导致大规模患病和死亡。在急性温度刺激下,紫贻贝()与翡翠贻贝()的耗氧率和排氨率均随温度升高而升高,这也与缢蛏的表现相似。在高温条件下还会导致部分双壳贝类产生强烈的氧化应激反应,自由基活性氧(ROS)水平上升,机体过氧化损伤加剧,影响正常的新陈代谢及功能的维持,从而导致其体质活力下降及抗逆性降低。为使体内的ROS保持在正常水平,生物必须及时清除过量的ROS。还原性谷胱甘肽(GSH)和过氧化物酶(POD)就是ROS清除系统中的重要成员,具有清除氧自由基、维持细胞膜完整性及DNA合成等多种生物功能,是衡量机体抗氧化能力的重要因素。
缢蛏作为中国一种重要的海水滩涂养殖贝类,2019年产量869 251 t,在养殖过程中同样会受到夏季高温的制约。尤其是在夏季退潮后,沿海滩涂养殖区或河口存有少量海水的情况下,缢蛏会被暴晒后沿着出入水口进入蛏穴的海水烫伤或烫死。为提高缢蛏的抗逆性,迫切需要开展缢蛏耐高温性状新品系的选育工作。
家系选育是选择育种中一种重要手段,近年来,已有对贝类的抗逆性、生长等性状选育的报道,如在生长方面,Evams等通过家系选育对4种不同养殖环境下的长牡蛎的平均体重、存活率和产量等指标进行了筛选;闫喜武等通过建立家系对菲律宾蛤仔()的卵径、受精率、孵化率及生长、存活和变态的相关指标进行了分析,为其后续挑选生长优良品种提供了理论支持。在抗逆性方面,刘项峰等验证了虾夷扇贝()的高温耐受性可以通过表型进行选育;王庆志等发现,早期筛选的虾夷扇贝耐高温家系的成活率明显高于普通家系。
同时,利用家系选育已获得了不少贝类新品系,如中国水产科学研究院南海水产研究所培育的马氏珠母贝()南珍1号,中国海洋大学研究团队培育的长牡蛎()海大2号,辽宁省海洋水产研究所选育的虾夷扇贝明月贝等。本研究是在本实验室经5代连续选育得到的缢蛏快长新品种申浙1号的基础上,继续进行耐高温抗逆性状的家系选育,通过监测各家系生长数据、分析特定家系的耐热性及抗氧化能力等情况,为种质改良、培育出生长快、耐高温的缢蛏新品系奠定理论基础。
1 材料与方法
1.1 材料
从浙江省三门县水产养殖基地采集性腺成熟的一龄缢蛏新品种申浙 1 号,并在浙江省三门县东航水产育苗科技公司进行暂养。
半致死温度的测定、控温装置由若干500 W 石英加热管、1 000 W 电子继电器、40 L泡沫保温箱、水银触点温度计(0.1 ℃)、500 mL烧杯以及气管气石等组成,所有温度计均经过严格的校准后使用,整套装置控温精度0.1 ℃。
1.2 方法
1.2.1 亲贝的筛选与暂养
选择活动力强、性腺饱满、表面无损伤、大小相似的新品种缢蛏申浙1号300枚暂养在水泥池,池底铺设30 cm 左右泥沙,砂滤海水,海水比重在1.009~1.016,pH为7.8,海水温度在28~33 ℃。暂养期间,池壁气管持续微充气。为保证水中饵料充足,定时投喂牟氏角毛藻()、亚心型扁藻()等混合单细胞藻类和虾塘水。每2 d全换水一次。
1.2.2 家系建立
2019年9月,亲贝性腺基本成熟,从暂养池中挖出经简单清洗后,阴干刺激12 h再放入遮光曝气的水中3 h后,开始观察到亲贝出水管产卵排精。将正在排精(雾状)的雄性和正在产卵(颗粒状)的雌性挑出来,用干净的沙滤海水清洗后分别放入盛有新鲜海水的2 L烧杯中,海水比重1.009。最终选取雌雄各16枚,建立了16个全同胞家系。将排好的精卵倒入盛有沙滤海水的50 L白色塑料桶中均匀搅拌,充气孵化。精卵结合后进行卵裂,首先发育为担轮幼虫,约18 h后发育成“D”形幼虫。整个操作过程中各实验组严格隔离,不受外来精卵干扰。
1.2.3 室内幼虫培养
浮游幼虫培育。受精卵孵化18 h后,用300目尼龙网袋过滤出“D”形幼虫,目的是将死卵剔除。幼虫培育在50 L白桶中进行。2 d后全换水1次。在第一次换水过程中会对幼虫密度进行调整,以消除养殖密度对生长速度的影响,使每个家系保持一致,调整密度为8~10 mL。第2、3日投喂湛江等边金藻(),每天投喂3次,第4日开始投喂湛江等鞭金藻和牟氏角毛藻,投喂量视幼虫摄食情况而定。为防止换水过程中各家系间出现混杂,换水工具单独使用。浮游期海水比重初始为1.009,后几日比重由1.009逐渐降至1.006。pH为7.8,水温在28~30 ℃。
稚贝室内培育。幼虫在第8天培育后基本变态为可附着生长的匍匐幼虫。在附着前两日往桶中投放适量的50目筛网过滤的无菌泥土。每5 d全换水一次,主要投喂牟氏角毛藻和虾塘水,每天两次。海水比重1.006,pH为7.8,水温在26~28 ℃。其他操作同前。
1.2.4 半致死温度
幼虫培育到稚贝期后,从3个壳长最长的家系和3个壳长最短的家系中随机挑选180只稚贝,再随机分为6组,分别放入一个常温实验组(室温约26~28 ℃)和5个高温实验组中(30、32、34、36、37 ℃)。为尽量避免快速升温对稚贝造成的影响,各高温组先后以每2 h升高1 ℃的速率缓慢升温,使各高温组同时达到预定温度,定期观察并记录各组稚贝死亡数,死亡的判定标准为两壳张开,外套膜无收缩能力。至所有组合不再出现死亡时实验结束。饵料经加温后再投喂,每天换水1/2,其他同前。实验持续8 d。设3组重复。温度设计参考文献[18]。
1.2.5 抗氧化能力测定
耐高温实验在加温前以及加温第1、3、7天每个温度下的每个家系随机挑取6只活力良好的缢蛏放入1.5 mL冻存管并立即放入液氮中冷冻,放入-80 ℃冰箱长期保存。实验时,首先称量算出净重,按质量体积比1∶9加入0.9 %的生理盐水并用匀浆机冰浴研磨。将研磨好的组织液放入低温离心机中,2 500×离心10 min,取上清液用于总蛋白(total protein,TP)、总抗氧化能力(total antioxidant capacity,T-AOC)、过氧化物酶(peroxidase,POD)、谷胱甘肽(glutathione,GSH)的测定。测定使用南京建成公司试剂盒,测定原理、实验步骤和计算方法均参照试剂盒说明书。
1.2.6 指标测定与数据分析
测量各家系缢蛏幼虫壳长。浮游期幼虫(壳长<300 μm)在显微镜下用目微尺(100×)测量,300 μm≤壳长<3.00 mm 的稚贝测量在体视显微镜下用目微尺(25×)进行,壳长>3.00 mm的幼贝用游标卡尺测量。每个家系用相同方法重复测量3次,每组随机测定30只幼虫。
真正存活率()=实验结束时存活数/实验开始总数;相对存活率()=真正存活率/对照组存活率。
使用SPSS 26.0统计软件及Excel 2016对文章数据进行分析处理,采用单因素方差(ANOVA)分析不同家系间的数据,温度效应间的显著性差异采用Duncan多重法比较,差异显著性水平设置为<0.05。使用Logistic模型确定半致死温度。GraphPad Prism 8.02、Excel 2016作图。
2 结果与分析
2.1 浮游期和稚贝期生长
实验共建立了16组全同胞家系(编号1-16),每组分别选取雌雄各1枚,共32枚亲本。家系1、2、9因浮游期成活率太低被淘汰。
浮游期第二日龄的壳长除家系8最长外,其他组合无显著差异(<0.05)。浮游期结束时,家系1、2、9因成活率太低被剔除。其中家系6、8、10、12、14、15、16壳长和壳高显著大于其他家系,其中家系14最大。稚贝期结束时,家系3、4、5的壳长显著低于其他家系。家系8、14、15、16的壳长和壳高显著大于其他家系(<0.05)(表1)。

表1 缢蛏不同家系在浮游期和稚贝期的壳长和壳高
2.2 稚贝期存活率
家系6、14在稚贝期的成活率显著高于其他家系,其中家系14的成活率最高,达到83.17%±0.85%,家系7的成活率最低,仅54.00%±2.64%(图1)。

不同处理间没有相同字母表示差异显著(P<0.05)。
2.3 半致死温度
在6个实验家系中,家系16的半致死温度最高,达到了36.64 ℃。家系3、4、5、14、15的半致死温度分别为34.79、34.71、34.23、36.55、35.73 ℃,壳长较长的家系与壳长较短的家系半致死温度差异显著(<0.05)(图2)。

壳长较短的家系编号为3、4、5,壳长较长的家系编号为14、15、16。
2.4 抗氧化能力测定
2.4.1 POD活性测定
实验组温度为30、32、34 ℃时,各家系POD活性随温度增高而增高,但之后开始下降,37 ℃时活性反而低于常温状态下的活性;随着高温的持续刺激,各家系1、3、7天的活性有一个缓慢下降的趋势。从3组壳长家系(家系14、家系15、家系16)与壳短家系(家系3、家系4、家系5)来比较,壳长各家系在32、34、36 ℃下的平均酶活性显著高于壳短家系的(<0.05)(图3)。

横坐标代表每个家系分别在0、1、3、7天不同温度下取样。数据以蛋白质质量计。下同。
2.4.2 GSH含量测定
GSH含量显示,各家系GSH的含量随温度升高而逐渐升高,并在37 ℃时达到最高。从三组壳长家系与壳短家系来比较,36、37 ℃下壳长各家系的GSH平均值显著高于壳短家系(<0.05)(图4)。

图4 温度对缢蛏家系还原性谷胱甘肽含量的影响
2.4.3 T-AOC测定
T-AOC趋势与POD活性趋势相似,同样是在34 ℃时达到最高,并且各家系在1、3、7天的T-AOC值有一个缓慢下降的趋势。但在36、37 ℃时T-AOC急剧降低且低于常温对照组。三组壳长家系在34 ℃时的平均T-AOC显著高于壳短家系(<0.05)(图5)。
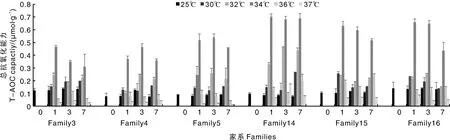
图5 温度对缢蛏家系总抗氧化酶的影响
3 讨论
3.1 各家系选育指标分析
家系选育是指以家系为单位进行的选择育种,其中水产生物通过对高表型值亲本的选择和选配可获得较好的选育效果,在鱼类、甲壳动物上均有报道。应用家系选育技术在贝类上的报道主要集中在提高贝类的生长速率、抗逆性等方面。本研究采用家系选育的方法来挑选耐高温家系后代。采用快长新品种申浙1号为基础群体,先期对亲贝筛选后,利用家系选育技术构建家系,测量各家系的生长数据。
在实验过程中家系1、2、9初期因死亡率较高已挑除外,家系6、14在稚贝期的成活率显著高于其他家系(<0.05)。稚贝室内培育期结束时,家系3、4、5壳长显著低于其他家系(<0.05);家系8、14、15、16壳长显著高于其他家系(<0.05),其中家系14最大,壳长和壳高分别为(7.04±0.72)mm,(2.83±0.29)mm。
选择壳长最长(家系14、家系15、家系16)和最短(家系5、家系3、家系4)的各3组家系进行半致死温度测定以确定壳长是否与温度耐受性相关。实验结果表明,三组壳长较长的家系半致死温度与三组较短的家系差异显著(<0.05),即壳长较长的个体对温度的耐受性要强于壳长较短的,由此推断个头较大的个体耐受性较好,这与刘志刚等对墨西哥湾扇贝()高温的耐受性的研究相似,不同规格贝对高温的耐受性为中贝=小贝>稚贝。同时,各家系对高温的耐受性可能不仅限于个体大小,同时还可能会受各家系的耐热基因型和遗传力的影响,如刘项峰等和王庆志等证实了虾夷扇贝在耐热性上遗传力的表现,同时还证明了早期筛选对养成阶段的重要性。该实验进程中保证了各家系培育条件相同:适宜的温度、新鲜且充足的饵料、氧气以及沙滤海水等,尽可能为各家系能充分展现其生长性能,避免外部环境等客观因素干扰提供了条件。
3.2 抗氧化能力分析
海洋贝类的研究表明,提高环境温度会显著影响海洋双壳贝类机体生理化过程,进而影响呼吸率、食物摄取、消化吸收速率以及排泄率等。Abele等对海洋贝类细胞水平研究表明,海水温度升高时会导致细胞代谢率上升,ROS水平上升,DNA、蛋白及脂质大分子的过氧化损伤加剧,细胞内稳态失衡。以上研究表明,环境温度的提升会导致机体氧化损伤加剧,影响其抗氧化能力,还会使机体代谢率上升。
在对缢蛏各家系的耐热实验中,在非致死高温(30~34 ℃)刺激下,缢蛏各家系的总抗氧化能力与POD活性上升,而当在更高的温度(36~37 ℃)持续刺激下,其抗氧化能力下降甚至达不到常温水平。这表明机体在耐受范围内可将体内ROS维持在正常水平,而一旦超过其耐受极限就会导致稳态失衡、抗性下降。这与Gueera等对大肚海扇贝()的研究结果相似,过高的温度反而会导致贝的代谢率由升高转为下降;当高温胁迫后的栉孔扇贝死亡率达到62%时,其外套膜、鳃和肾脏的抗氧化酶基因、的表达水平对比常温对照组显著下降,这也与本文的结果相似。GSH含量在各家系均呈现持续上升的趋势,作为一种清除ROS的重要底物,其通过还原活性氧来保护细胞,机体为努力维持过氧化物和自由基的平衡而不断的产生GSH。
横向比较来看,壳长较长组与较短组的差异明显,直观表现为各家系的存活情况也就是半致死温度的差异,内在表现为三种抗氧化指标的变化。在受到高温持续刺激且超过机体耐受范围情况下,抗氧化能力会受到显著影响,机体无法维持正常的新陈代谢,组织氧化损伤急剧上升,甚至造成个体死亡。纵向来看,各家系在同一温度持续刺激下,第1、3、7天的抗氧化能力基本呈现小幅下降趋势,推测缢蛏体内的抗氧化系统即使在非致死高温环境下也难以长时间维持最初的水平。
与正常温度相比,高温胁迫下各家系个体的抗氧化能力受到显著影响,且壳长较长的缢蛏个体的综合体质状态及抗逆性明显好于较短的。因此,本研究推测夏季高温期,缢蛏自身体质及抗性的下降是诱发其死亡的根本原因。在实际生产中,广东、福建等地由于缢蛏性腺成熟较早,8月份就会有工厂开始缢蛏育种,此时应注意将水温控制在幼虫适宜的范围内。同时幼虫在转移至土池中间暂养后也应注意大潮换水时存蓄的少量海水的温度,避免暴晒太久而使水温过高。
采用新品种申浙1号构建家系并评估子一代的壳长、存活率和耐热性等差异,在保留快长的基础上,利用家系选育和高温实验挑选具有抗逆性的耐高温后代,并通过抗氧化能力的测定在生理生化的角度证明了家系挑选的必要性。其中家系14生长最快、存活率高且耐热性较好,最终确定家系14是适合生产的最优家系,家系15、16可用作备选家系。同时为后续缢蛏抗逆性新品系的筛选提供了理论依据。
猜你喜欢
天津农业科学(2022年6期)2022-07-17
疯狂英语·新阅版(2021年8期)2021-09-10
时代英语·高二(2021年4期)2021-07-29
农民致富之友(2019年31期)2019-11-23
保健与生活(2019年13期)2019-08-27
现代交际(2016年20期)2017-02-15
时尚北京(2016年12期)2017-01-16
农村农业农民·B版(2016年7期)2016-10-21
杂文选刊(2007年16期)2007-05-14
