
紫叶紫薇花芽分化及雌雄配子体发育的形态学研究
2022-03-02 03:53:28罗雪梦王晓明曾慧杰李永欣乔中全
中南林业科技大学学报 2022年2期
罗雪梦,王晓明,曾慧杰,李永欣,乔中全,唐 丽
(1.中南林业科技大学,湖南 长沙 410004;2.湖南省林业科学院,湖南 长沙 410004)
紫薇Lagerstroemia indicaL.广泛分布于我国西南、华南、华北地区,目前国内有21 个城市将紫薇列为市花[1]。因其具有树型优美、花色艳丽、花期长等优点,在公园绿化、道路绿化、花境配置、盆景造型等方面应用颇多,具有极重要的园林价值和经济价值。紫叶紫薇Purple-leafLagerstroemia indica为千屈菜科Lythraceae 紫薇属Lagerstroemia小乔木或落叶灌木,枝干直立性强,花色深红色,嫩叶紫红色,成熟叶黑紫色。
国内外对紫薇的研究主要集中于引种、组培快繁、种质资源调查以及遗传性状研究等方面[2-7]。国内有少数学者进行了紫薇种子萌发以及扦插育苗方面的研究[8-9]。紫薇属植物在生殖生物学方面国内已有少数报道,郑绍宇等[10]对狭瓣紫薇Lagerstroemia stenopetalaChun 花粉活力进行了研究,并通过不同激素处理提高其柱头可授性;曾慧杰等[11]通过“不育紫薇”和“可育紫薇”分别自交与交叉杂交后检测其花粉活力和花粉萌发情况,表明“不育紫薇”的不育性状由雌、雄性不育共同所致;蒙真铖[12]对毛萼紫薇Lagerstroemia balansae传粉机制、繁育系统及种子萌发进行了探讨,为其深入的保育工作提供了研究背景;袁娟[13]首次对大花紫薇Lagerstroemia speciosa的不同水平开花物候、花粉活力、花部形态与功能、雌雄配子体发育过程进行了系统研究,充实了紫薇属植物生殖生物学研究的不足;郭玉敏[14]对日本矮紫薇Lagerstroemia indicacv.花芽形态分化、大小孢子发生和雌雄配子体形成过程的解剖结构进行研究,明确了各花器官分化顺序,掌握了花芽分化与营养生长的相关性;许欢[15]比较正常结实紫薇与不结实紫薇的花芽分化内部结构差异,从形态学方面阐明不结实紫薇的雌雄性不育主要表现为生理型花粉败育及胚囊败育。
紫叶紫薇以其寓意美好的深红色花而受到大众的喜爱,决定紫叶紫薇的成花时间、质量及数量等的关键基础是花芽分化。花芽分化是生殖生物学重要的组成内容,包含一系列复杂的形态结构变化、生理指标变化及开花基因变化等[16]。同种植物的花芽分化进程也会因不同品种和不同地区的栽培条件、气候条件等有所不同,因此要对紫叶紫薇花发育进行深入的试验研究,就要明确其在研究区域的花芽分化过程。目前,紫叶紫薇在生殖生物学方面还缺乏研究资料,首先应对紫叶紫薇的细胞学和形态学进行研究。本研究以选育良种‘丹红紫叶’Lagerstroemia indica‘Ebony Embers’为试验材料,对紫叶紫薇的花芽形态分化和雌雄配子体发育过程进行观察,掌握紫叶紫薇在中南地区的花芽分化时间及整体进程,明确各分化时期的内部结构特征,为紫叶紫薇花期调控和栽培管理提供科学参考,为进一步研究紫叶紫薇成花机理及物候因素奠定基础,为紫薇属植物生殖生物学的长期发展积累资料。
1 材料与方法
1.1 试验地概况
试验地位于湖南省长沙市雨花区湖南省林业科学院林木新品种创制中试基地,地处113°01′E,28°06′N,属低山丘陵地貌,地势较平缓,坡度15~20°。属亚热带季风性湿润气候,年均气温17℃,年均降水量1 400 mm,年均日照时数1 500 h。试验地土壤为第四纪酸性红壤,pH 值为4.0~5.5。
1.2 试验材料
试验材料选取5年生紫薇品种‘丹红紫叶’。平均树高约3 m,树势中等,长势良好。
1.3 观察及取样方法
2020年3月下旬进入抽枝展叶期,此时选择长势基本一致的5 株植株为试验对象,每株植株在东、南、西、北4 个方向各选择5 枝生长状况良好的新枝,每枝条挂上标签,4月每7 d 观察当年生枝顶芽并采集,5月上旬每2~3 d 观察当年生枝顶芽并采集,5月中旬至6月中旬每2~3 d观察当年生枝花序顶端花蕾并采集,经体视显微镜观察后放入FAA 固定液(50%酒精∶甲醛∶冰醋酸=18∶1∶1)中固定,4℃下保存,用于石蜡切片和显微观察。
1.4 石蜡切片制作方法
采用石蜡切片法制片,经固定、抽气后的材料按50%、70%、85%、95%、100% 酒精浓度梯度逐级脱水(据材料大小不同实际处理时长20~60 min),体积比1∶1 的无水酒精、二甲苯混合液透明1 次,纯二甲苯透明2 次(均处理1 h),放置材料于体积比约1∶1 的二甲苯和石蜡中37℃下过夜。于60℃下更换纯石蜡3 次,再进行包埋、切片、展片、烘片,切片厚度为8~10 μm。60℃下纯二甲苯脱蜡3 次,由高至低梯度浓度乙醇复水,硫酸高铁铵媒染(现用现配),纯净水冲洗,苏木精染色,乙醇再次脱水,二甲苯再次透明,树胶封片,Olympus 显微镜下观察。
2 结果与分析
2.1 花芽形态分化
紫叶紫薇花芽分化为当年分化当年开花型,其顺序大致分为未分化期、分化初期、花序分化期、萼片分化期、花瓣分化期、雄蕊分化期和雌蕊分化期。
2.1.1 未分化期
5月10日前,当年生花枝尚在营养生长期,花芽处于未分化期,此时生长点处于发生生理变化的花诱导阶段,诱导营养生长向生殖生长转变,积累营养物质,为花芽分化奠定基础。生长锥顶部较扁,表层细胞体积小而排列紧密。生长锥外由4 对片状结构的苞叶包裹,起保护作用(图1A)。
2.1.2 分化初期
5月10日前后,顶芽开始分化,顶端分生组织快速分裂,生长锥经伸长增宽生长后体积较未分化期略有增大,形状由平变圆,顶部隆起呈半球形(图1B)。
2.1.3 花序分化期
5月12日前后,生长锥一侧或两侧有新的突起,体积不断膨大,经伸长生长形成圆锥花序,每花序可形成1 朵或数朵小花(图1C)。
2.1.4 萼片分化期
5月中旬观察可见生长锥顶部由半球形逐渐变平坦,生长锥基部外围向上突起,小突起即为萼片原基,纵切面仅见2 枚萼片原基(图1D)。萼片原基向上伸长生长,逐渐向内弯曲直至靠拢形成闭合的弧形萼筒,萼片原基形成萼片的速度较慢(图1E)。
2.1.5 花瓣分化期
5月20日前后,观察可见花瓣原基于萼片内侧边缘偏上的位置突出(图1F),同时生长锥基部继续变宽。花瓣原基于发育后期向下伸长,形成弯曲褶皱的花瓣(图1H)。
2.1.6 雄蕊分化期
5月23日前后,生长点基部变宽,产生密集的间隔式小颗粒,此时周边位置为雄蕊原基,中心位置为雌蕊原基(图1G)。此后,雄蕊原基发育速度略快,发育时间较雌蕊原基更早,上部横向分裂旺盛,膨大形成花药。雄蕊原基体积增大过程中数目仍不断增加,最终若干个突起形成雄蕊群围绕着中部较大的雌蕊原基(图1H)。
2.1.7 雌蕊分化期
5月28日前后,中心雌蕊原基开始形成膨大的子房,多个心皮凹陷形成子房室,子房顶部逐渐愈合(图1I)。雌蕊顶端继续向上伸长形成花柱及柱头,每个子房室内开始分化胚珠原基。
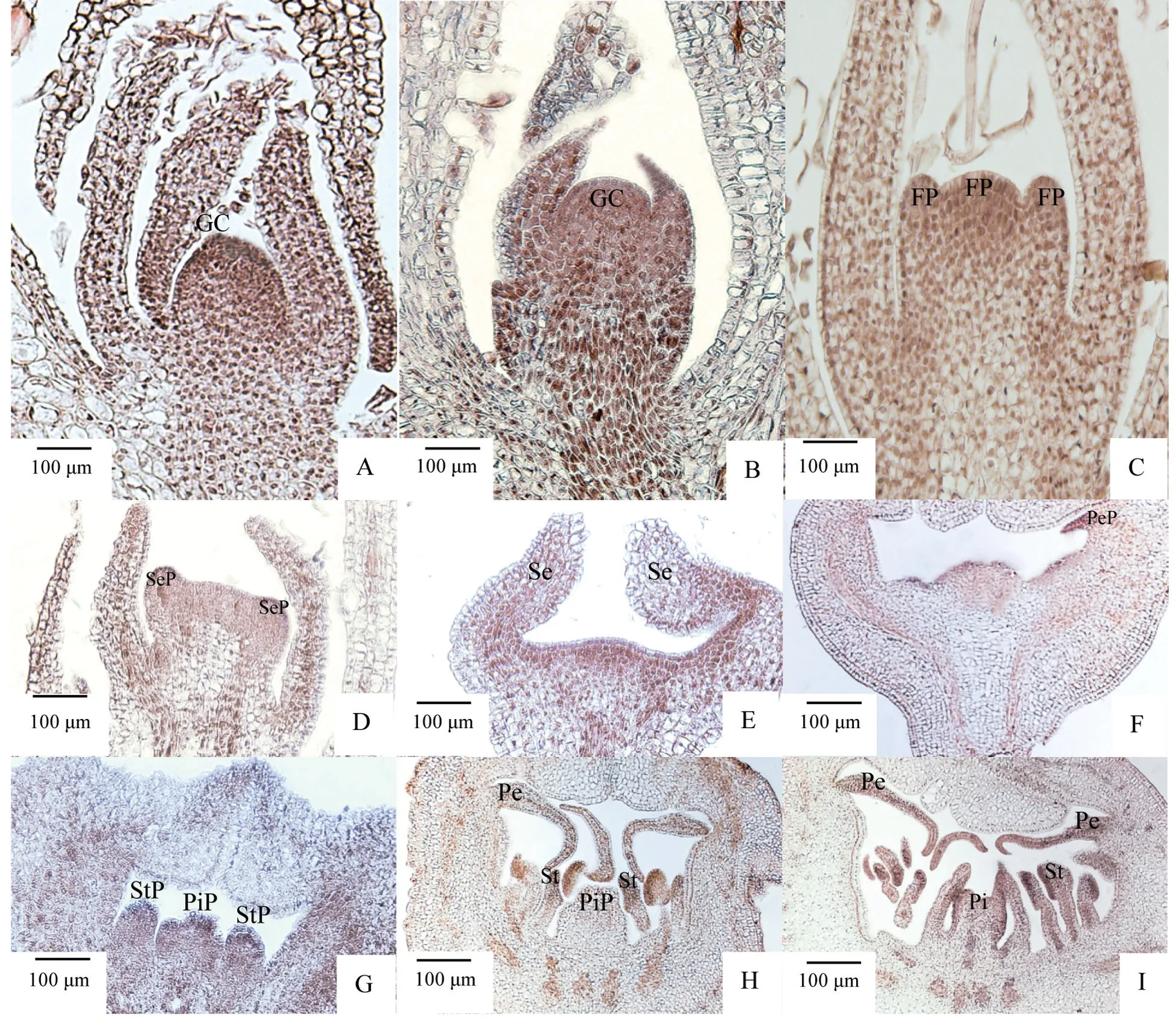
图1 紫叶紫薇花芽形态分化不同时期解剖结构Fig.1 The anatomical structure at different periods of flower bud morphological differentiation
从5月上旬至下旬基本完成花芽形态分化过程(图2),此过程中分化速度非常快,从花芽分化初期至最早雌雄蕊原基发生历经约十几天便进入各性器官的进一步发育阶段。
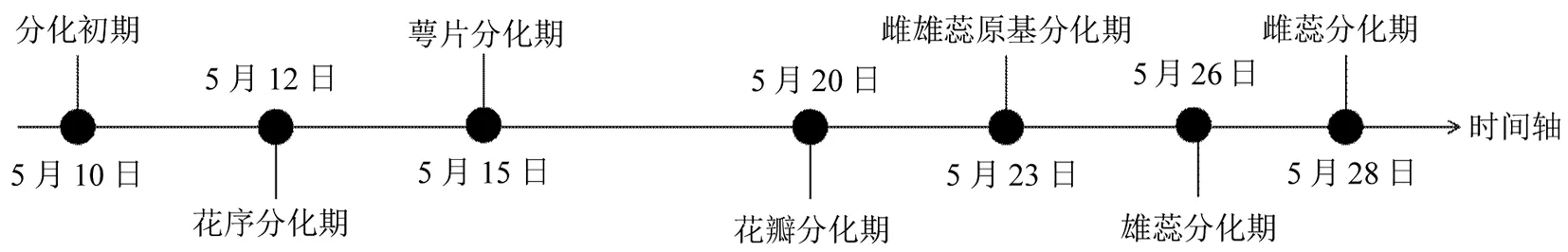
图2 紫叶紫薇花芽形态分化时间Fig.2 The morphological differentiation of flower buds of purple-leaf Lagerstroemia indica
2.2 雄蕊发育
2.2.1 小孢子发生
紫叶紫薇的雄蕊多数,呈紧密排列状,有长雄蕊和短雄蕊之分,长雄蕊花药形态大,黑褐色,一般不可育;短雄蕊花药形态小,黄色,一般可育,文中观察的主要为可育雄蕊。5月23日左右,若干个雄蕊原基环绕心皮原基(图3A),从横切面观察未分化花药轮廓似椭圆形,最外一层是表皮细胞,表皮内为逐渐分裂的活跃细胞(图3B)。5月26日前后,花药先于心皮开始分化,顶端分生组织逐渐活跃,细胞分裂较快,形成幼嫩花药(图3C)。幼小花药的4 个角隅细胞分裂更快,一般形成4 个花粉囊,从横切面看花药形似蝶状(图3D)。在四角处表皮细胞最邻近的细胞层分化形成孢原细胞(图3E),随后孢原细胞通过平周分裂使细胞层增多,初生造孢细胞发生于内层,比周围细胞体积更大,细胞质更浓,细胞核更明显(图3F)。5月28日前后,各细胞间出现细微间隔且细胞形状呈多边形,这是由初生造孢细胞经有丝分裂形成的次生造孢细胞(图3G)。次生造孢细胞分化小孢子母细胞的过程中胼胝质不断沉积,为小孢子母细胞增大体积提供某些营养物质(图3H)。5月30日前后,胼胝质逐渐溶解使小孢子母细胞逐渐分离,单个小孢子母细胞体积大、细胞核明显,形状由多边形渐变圆形,液泡不明显(图3I)。
小孢子母细胞进入第一次减数分裂,染色体高度螺旋化,核仁开始消失(图3J),减数分裂末期小孢子母细胞形成2 个子细胞共处1 个胼胝质壁内,此时并未形成细胞壁与细胞板(图3K)。第二次减数分裂末期每个子细胞再次分裂,小孢子母细胞形成4 个子核,但大部分在同一平面上只看到3 个核,核仁明显,子细胞之间的小孢子母细胞表面开始形成新的细胞壁(图3L)。两次减数分裂完成后,4 个小孢子由胼胝质壁分离完全,排列形成四面体型四分体(图3M),依据紫叶紫薇减数分裂及四分体的情况,其胞质分裂方式为同时型。四分体后期,胼胝质壁正常降解,四面体型四分体解体,小孢子离散至花粉囊中成为单个的小孢子,早期的单核小孢子体积较小,细胞壁薄且略收缩(图3N)。
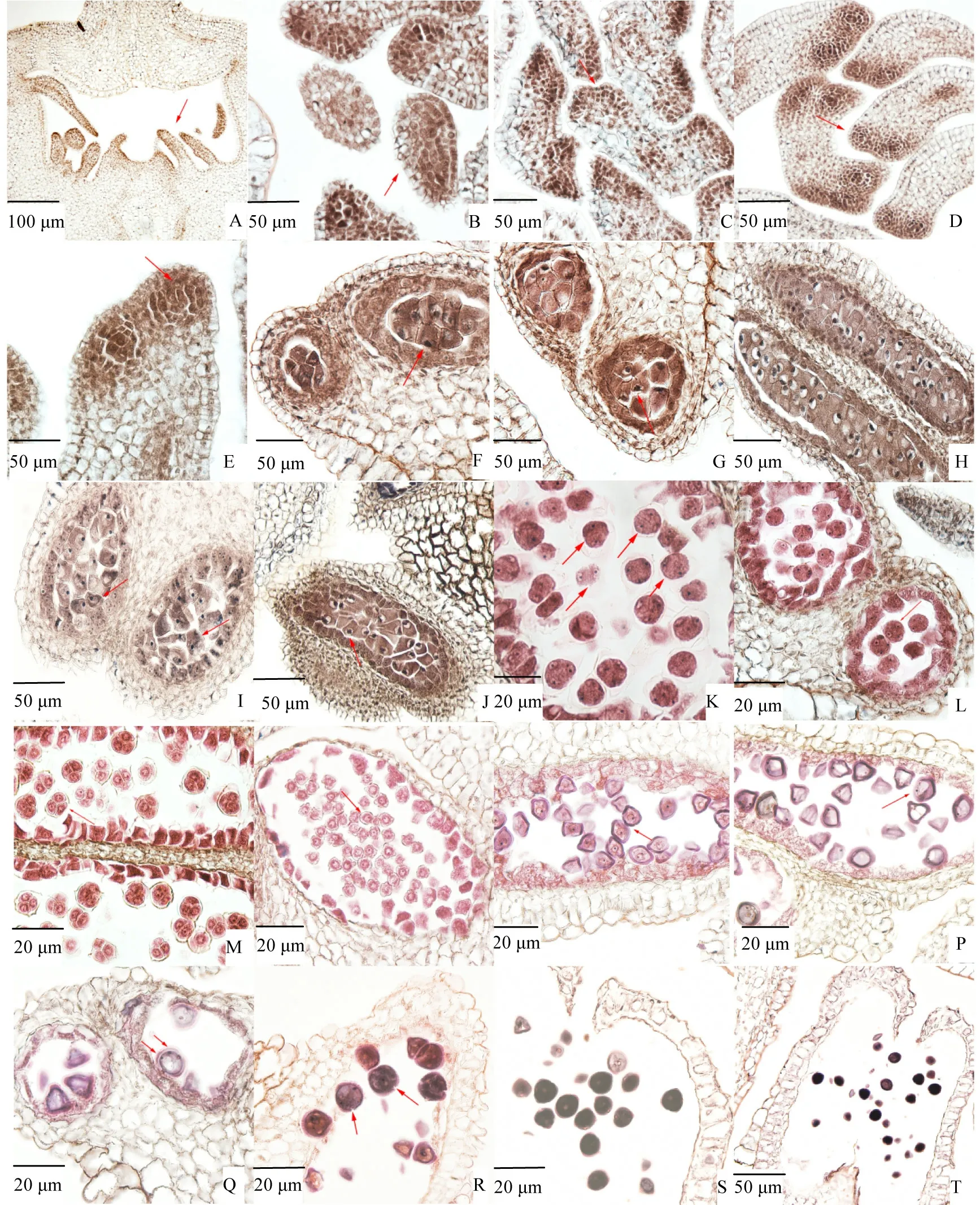
图3 紫叶紫薇小孢子发生和雄配子体发育Fig.3 The microsporogenesis and development of male gametophyte in purple-leaf Lagerstroemia indica
2.2.2 雄配子体发育
6月6日前后,单核小孢子进入雄配子体发育时期,小孢子体积显著增大,细胞壁增厚,细胞核位于小孢子中央,形成单核中央期的小孢子(图3O)。细胞核逐渐向细胞壁移动形成单核靠边期的小孢子,这是由于胞质液化使液泡体积增大,液泡占据小孢子的主要空间,将细胞核推向细胞壁(图3P)。细胞核于靠近细胞壁位置发生有丝分裂,形成营养核与生殖核,形成二核花粉粒(图3Q)。紫叶紫薇花粉粒表面具三萌发孔沟,可从花药壁吸收养分(图3R),6月11日前后,花粉逐渐发育成熟,细胞质浓厚,细胞内含有大量淀粉等营养物质(图3S)。成熟花粉粒为二细胞型,于开花前相邻花粉囊破裂连通(图3T)。
紫叶紫薇完成花芽形态分化后,其雄蕊发育过程即小孢子发生和雄配子体发育从5月下旬至6月中旬基本完成(图4)。
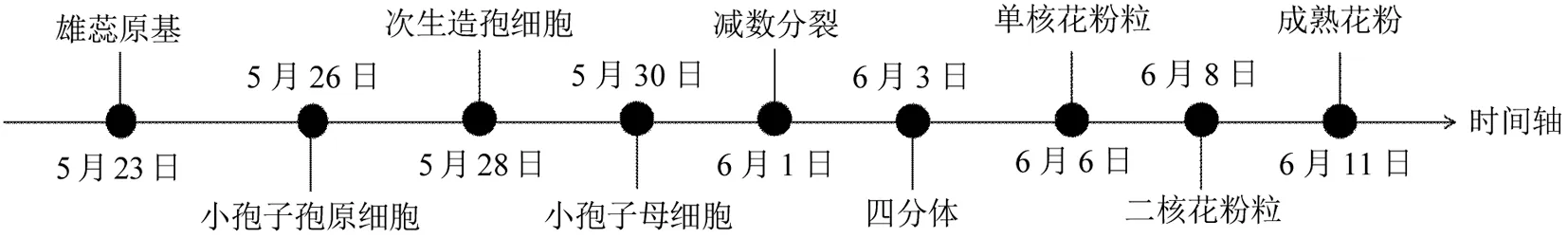
图4 紫叶紫薇雄蕊发育时间Fig.4 Stamen differentiation of purple-leaf Lagerstroemia indica
2.3 雌蕊发育
2.3.1 大孢子发生
5月下旬,紫叶紫薇雌蕊原基继续膨大发育,各心皮上部闭合形成花柱和柱头,下部多心皮表皮细胞分化闭合,构成中轴胎座和六室子房(图5A)。5月末,子房壁内壁细胞快速分裂,胚珠原基着生于中轴胎座(图5B),胚珠原基进一步发育为倒生型胚珠,胚珠前端为一团薄壁组织,即珠心(图5C),紧靠珠心外围先后突出内外双层珠被。珠心表皮下有一个细胞相较于周围细胞其体积显著增大,分裂更为快速,形成大孢子孢原细胞,此时可见二分之一范围的珠心组织由双层珠被包裹(图5D)。6月初观察显示大孢子孢原细胞快速分化,大孢子母细胞形成,此时珠心组织已由双层珠被全包裹(图5E)。大孢子母细胞经第一次减数分裂形成两个显著的子核,新细胞壁出现后大孢子母细胞发育为大孢子二分体(图5F)。二分体再经第二次减数分裂,大孢子四分体形成,4 个大孢子体积大小不完全相同,4 个大孢子中的2 个珠孔端大孢子及1 个合点端大孢子的细胞核凝缩,出现退化迹象(图5G)。大孢子四分体形成后快速进入大孢子解体状态,2 个珠孔端大孢子与1 个合点端大孢子退化后的残留细胞体积明显缩小,形状不规则,胞质浓厚,无法识别细胞核,另一个靠近合点端的大孢子吸取了充足的养分,体积继续膨大,细胞核明显,形成功能大孢子,胞质逐渐液泡化,分化形成多数小液泡(图5H)。
2.3.2 雌配子体发育
功能大孢子继续分化,多数小液泡聚集形成几个大液泡位于细胞核周围,单核胚囊形成,其细胞核大、核仁明显(图5I)。6月9日前后观察显示单核胚囊经有丝分裂后形成2 个细胞核,细胞中央位置由大液泡占据,2 个细胞核分别向胚囊的珠孔端与合点端反向移动,二核胚囊分化形成(图5J)。二核胚囊经过第二次有丝分裂,珠孔端与合点端分别有2 个子核,6月11日左右四核胚囊形成,胚囊形似椭圆形或纺锤形(图5K)。第三次有丝分裂后八核胚囊形成,珠孔端与合点端各有4 个细胞核,八核胚囊发育迅速,难以观察到胚囊两极分别有4 个细胞核共处同一平面。珠孔端与合点端各有1 个游离细胞核向胚囊中央位置移动,互相靠近(图5L),紧密结合的两个极核互相融合,中央细胞形成2 个核仁共存的细胞核(图5M),当两个极核完全融合时形成一大一小核仁的次生核。胚囊两极分别保留3 个细胞核,珠孔端的3 个细胞核分化形成1 个卵细胞及2个助细胞,构成卵器,呈三角形排列(图5N)。刚组合成卵器时卵细胞与助细胞体积较小,逐渐分化成熟后形态显著增宽伸长,成熟的卵细胞及助细胞核仁明显,细胞内具大液泡(图5O)。合点端的3 个细胞核分化形成3 个较小的反足细胞,反足细胞保持游离核的状态,排列不完全规则(图5P)。紫叶紫薇成熟胚囊即雌配子体由3 个反足细胞、2 个极核、2 个助细胞和1 个卵细胞形成,2 个极核在受精前融合后与大量周围胞质形成1 个大型中央细胞,这种7 细胞8 核型胚囊被称为蓼型胚囊。
紫叶紫薇雌蕊发育过程即大孢子发生和雌配子体发育从5月下旬至6月中旬基本完成(图6),整个花芽分化过程初始至成熟历经大约1 个月,此后花蕾含苞待放,几天后到达初花时间。
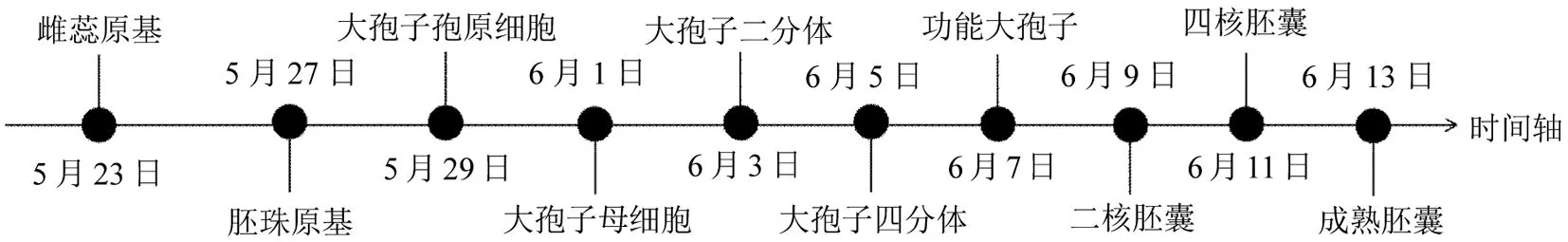
图6 紫叶紫薇雌蕊发育时间Fig.6 Pistil differentiation of purple-leaf Lagerstroemia indica
3 结论与讨论
花芽分化包括生理分化、形态分化和性细胞形成3 个阶段[17]。植株经过必要的营养生长后,当外界环境满足茎尖生长点从营养生长转变为生殖生长的条件时,芽分生组织开始发生生理变化和一系列形态变化,逐渐产生各花器官原基并最终发育为成熟花器官[18]。本研究观察紫叶紫薇整个花芽分化过程34 d 左右,顶芽于5月上旬开始分化进入花序分化期,5月中旬进入萼片分化期和花瓣分化期,5月下旬出现雌雄蕊原基,5月底至6月初,大、小孢子发生及雌雄配子体发育,6月中旬形成成熟花粉和成熟胚囊。因此将紫叶紫薇花芽分化大致分为未分化期、分化初期、花序分化期、萼片分化期、花瓣分化期、雄蕊分化期和雌蕊分化期7 个时期,划分标准是紫叶紫薇花芽内部相对应的原基开始分化。本研究结果表明紫叶紫薇的雌雄蕊原基几乎同步发生,随后雄蕊的分化时间稍早于雌蕊且形态发育速度有差异,前期花药分化速度比子房稍快,后期胚囊分化速度加快,最终成熟花粉和成熟胚囊进程基本相当。紫叶紫薇大、小孢子发生及雌雄配子体发育过程均正常,与大多数双子叶植物发育过程相似,其雌雄配子体的解剖结构特征也符合紫薇属植物的基本结构特征。
目前对花芽分化没有确切的阶段划分标准,但通常花芽分化划分的依据是由外至内各花器官形成的顺序,首先萼片,接着花瓣,然后是雄蕊和雌蕊[19]。本研究将紫叶紫薇花芽分化分为7个时期,与前人对日本矮紫薇[14]、结实紫薇和不结实紫薇[15]的划分结果相似。不同植物种类具有自身特异性,因此在植物花芽分化的形态学研究中,研究者对时期的划分也存在差异。如李心等[20]将‘木门’百合Liliumoriental×trumpet hybrid‘Conca D’Or’花芽分化进程分为花芽分化初期、小花原基分化期及花器官分化期3 个时期。汪晓谦等[21]将郁金香Tulipa gesnerianaL.花芽分化过程分为未分化期、花瓣分化期(包括外轮花瓣分化期与内轮花瓣分化期)、雄蕊分化期、雌蕊分化期5 个时期。胡梅等[22]将竹叶花椒Zanthoxylum armatumDC.花芽分化分为未分化期、花芽分化始期、花序轴分化期、花蕾分化期、萼片分化期、子房分化期和花柱分化期7 个时期。赵云芳等[23]将蓝花丹Plumbago auriculata花芽分化分为营养生长期、花芽分化前期、小花原基分化期、萼片原基分化期、花瓣原基分化期、雄蕊原基分化期、雌蕊原基分化期、花粉形成期及子房膨大期9 个时期。
紫叶紫薇花药具四花粉囊,小孢子母细胞体积大、细胞核显著、细胞游离,胞质分裂为同时型,小孢子四分体为四面体型,小孢子具三萌发孔沟,成熟花粉粒为二细胞型。在前人对紫薇花粉母细胞减数分裂的研究中,有学者认为日本矮紫薇胞质的分裂方式为连续型,而本研究发现紫叶紫薇胞质的分裂方式为同时型,这与袁娟[13]对大花紫薇、许欢[15]对结实紫薇与不结实紫薇的研究结果一致。大花紫薇的小孢子四分体排列形式以四面体型为主、偶有左右对称型存在,而本研究发现紫叶紫薇的小孢子四分体均为正四面体型,与日本矮紫薇的小孢子四分体排列形式相同。除此之外,前人观察发现日本矮紫薇少数花药中具6个花粉囊,且成熟花粉粒为三细胞型。四分体、药室、成熟花粉等形态学特征不仅可作为紫薇属内分类的依据,还可判断与其他属的亲缘关系。本研究显示紫叶紫薇不同花药间、同一花药不同花粉囊间以及同一花粉囊均存在小孢子母细胞发育时期略有差异的情况,处于减Ⅰ的小孢子母细胞与减Ⅱ的小孢子母细胞同时存在,或单核小孢子与二核花粉粒同时存在,此类不同步现象也在不少自然界其他种类植物的雄蕊发育研究中出现,如袁德义等[24]研究油茶‘湘林1 号’Camellia oleifera‘Xianglin No.1’、石艳兰等[25]研究二倍体芒diploidMiscanthus sinensis、董立格等[26]研究‘晚银桂’Osmanthus fragrans‘Wanyingui’。推测出现这种现象的原因可能是胼胝壁向内扩张分离小孢子母细胞时,细胞质营养分配不均衡,或是一些特殊的减数分裂基因调控异常等遗传原因,或是前期的孢原细胞、造孢细胞分化先后时间有差异,或与环境因素有关,或与花粉囊空间位置差异有关。紫叶紫薇小孢子母细胞减数分裂不同步的原因仍有待进一步研究。有学者认为减数分裂不同步导致四分体小孢子的有丝分裂异常,引起花粉败育[27];也有学者认为不同步现象是该植物长期适应自然环境的一种传粉机制[28]。本研究认为花药散粉时间有差异,有利于延长传粉受精时间,促进繁殖进化,降低恶劣气候条件的影响。在紫叶紫薇的开花物候方面,开花早的植株可能出现花果共存现象,其单株开花不同步性可能与小孢子发育的不同步相关,在一定程度上有利于延长花期持续时间。
紫叶紫薇子房六室,中轴胎座,倒生胚珠,双层珠被;大孢子孢原细胞形成时,珠心由珠被包裹二分之一;大孢子母细胞形成时,珠心由珠被完全包裹;减数分裂后4 个大孢子中仅余合点端1 个大孢子逐步分化,最终形成7 细胞8 核胚囊。紫叶紫薇的大孢子发生及雌配子体发育过程与前人对紫薇其他种的研究结果基本一致,日本矮紫薇子房除六室以外具四室、五室,可能是日本矮紫薇属于矮化型紫薇,其花部特性相较于紫叶紫薇更小。通过对四分体形态多次切片发现4 个大孢子难以完整切在同一平面上,因此大孢子四分体并非呈完全的直线型排列,而是“S”型排列方式。紫薇属植物在有性生殖中均是单个大孢子形成雌配子体,而另外3 个大孢子退化消失,这种细胞退化的机理和遗传基础有待深入研究。
紫叶紫薇经历30 d 左右完成花芽分化,与菊花、蝴蝶兰、郁金香、西红花等草本花卉及一些灌木花卉如野生茶树、金花茶、蓝花丹、木兰、勒杜鹃等的花芽分化速度接近。大多数木本植物都要经历较长的花芽分化时间,如荔枝需要4~5个月,苹果需要约5 个月,八仙花需要约4 个月,钝叶柃需要约4 个月,竹叶花椒需要约7 个月。紫叶紫薇花芽分化的时间短、速度快,属于一年多次分化型,不需要经过低温诱导即可在短时间内开花,除了遗传因素起决定作用外,还可能与成花过程的温度与光照等环境因素有密切关系,紫叶紫薇花芽分化与气候因子等的关系有待进一步研究。紫叶紫薇的快速发育可能会极大地消耗植物体内的营养物质,刺激内源激素的动态平衡,如袁小军等[29]认为矿质营养对油茶花芽生长及分化起到关键作用,周芳萍等[30]认为板栗花芽分化受到内源激素种类及比值的共同影响。本研究缺乏营养成分及内源激素在紫叶紫薇花芽分化过程中的调控作用研究,同时还需要了解各生理生化指标的相关性。通过了解紫叶紫薇整个花芽分化的时间进程,为下一步花期调控实践奠定基础,在紫叶紫薇的花芽分化期采取一定措施,为提早或延迟花期提供有利条件。掌握紫叶紫薇花芽分化的形态特征,使其既可作为授粉树种,也可作为优良的杂交母本,为后续的杂交育种工作提供参考资料。
猜你喜欢
果树学报(2023年1期)2023-02-28 19:59:52
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02 01:56:08
北京园林(2020年2期)2020-01-18 03:30:56
计算机应用(2018年10期)2018-11-22 09:37:54
现代园艺(2017年13期)2018-01-19 02:28:02
华南师范大学学报(自然科学版)(2017年4期)2017-09-11 09:16:23
浙江大学学报(农业与生命科学版)(2017年2期)2017-05-19 07:41:15
花卉(2016年1期)2016-02-17 07:10:18
上海质量(2015年2期)2015-12-02 02:26:44
中国民族民间医药·下半月(2015年4期)2015-05-22 07:12:43
