
光照强度和水分对花榈木幼苗结瘤及生理特性的影响
2022-03-02 03:53:16安常蓉韦小丽
中南林业科技大学学报 2022年2期
葛 萌,安常蓉,韦小丽
(1.贵州大学 林学院,贵州 贵阳 550025;2.贵州省生物研究所,贵州 贵阳 550009)
花榈木Ormosia henryi属豆目蝶形花科红豆属,又名花梨木,为国家二级保护植物,我国特有种。其材质优良,树形通直美观,是常见的名贵硬木,也是优良的园林绿化树种,同时其根、皮、茎、叶均可入药,因此具有较高的市场经济价值[1]。有研究表明,环境因素可对豆科树种结瘤、苗木生长和固氮效应产生强烈影响,使得固氮速率在时间和空间上差异很大[2]。因此,寻找影响花榈木幼苗结瘤和生长发育的重要环境因素,对花榈木的人工栽培驯化和发展扩大种群具有重要的意义。
花榈木苗期与根瘤菌共生,通过固氮作用为植物提供生长所需的氮素营养。根瘤菌是通过豆科植物根毛或其他部位侵入进入到根皮层,刺激宿主细胞分裂,形成根瘤[3]。而根瘤之所以能够有效固氮,是因为根瘤中固氮酶和豆血红蛋白共同作用的结果[4]。但是根瘤菌的固氮效应除受宿主树种的生长状况影响外,还受到外界环境的重要影响。光照强度对根瘤的固氮作用影响非常大,可以显著影响根部结瘤效应和植物体内生理特征的变化,因为根瘤菌生存所需的能源依赖于植物光合作用的产物,而光强是植物进行光合作用的前提。光强不足时,植物光合作用弱,不仅根瘤数量和质量减少,根瘤的固氮酶活力也显著下降[5]。当豆科植物处于黑暗条件下时,根瘤中蔗糖合成酶活性和转录组水平会显著降低。但长期在高光强下生长的植物或者弱光下生长的植物突遭受强光时,光合作用可能会受到抑制,进而导致光合速率下降[6],同时,高光强也会导致植物体内叶绿素和类胡萝卜素快速分解而降低[7],进而影响结瘤和幼苗生长发育。
根瘤能否固氮与植物的光合作用有很大关系,而光合作用不仅受到光照、CO2浓度等环境条件的影响,还受到水分的重要影响。水分不仅会影响豆科植物根瘤形态、数量、大小及生理特性[8],还会影响植物体的细胞结构[9]、根系分布[10]、叶绿体光化学及生化活性以及各种酶的活性[11]。随着干旱胁迫程度的加剧,根瘤菌侵染细胞数逐渐减少,不仅影响根瘤菌与寄主的识别过程,还会显著降低根瘤的形成和固氮能力[12]。
综上所述,光照强度和水分对豆科植物的结瘤和生理特性的影响非常重要,迫切需要开展试验以寻求花榈木幼苗生长发育所需的最佳光照强度和基质水分条件。本研究以1年生花榈木幼苗为研究对象,通过人工模拟不同梯度下的光照强度和基质含水量,探究其对花榈木幼苗结瘤效应和生理生化特性的影响,旨在寻找适合花榈木幼苗生长、结瘤及固氮效应的最佳光照和水分环境,为提高花榈木苗木结瘤率和固氮能力提供技术指导。
1 材料与方法
1.1 试验材料
1.1.1 花榈木种子
在贵州石阡县选择花榈木同一母株上规格一致、粒大饱满、无病虫害的健康花榈木种子。千粒重300 g,种子长11.32±0.77 mm、宽8.95±0.57 mm。
1.1.2 幼苗培养
于2月中旬对种子进行消毒、灭菌并进行催芽处理,待胚芽突出2~3 cm 时,转移至盛有灭菌基质的花盆中,基质配比为泥炭∶蛭石∶珍珠岩=2∶1∶1。每盆播种4 粒催芽种子并浇透水,覆盖保鲜膜。随后置于75%全光照、25℃恒温下的温室培养箱中培育,定期浇水,待幼苗全部出齐并长出2~3 片真叶后开始进行处理。
1.1.3 根瘤菌菌液制备
将采集的新鲜、个大饱满的根瘤用无菌水洗净,在无菌条件下用75%的乙醇溶液消毒30 s,接着用1‰的HgCl2溶液消毒5 min,用无菌水冲洗5 次以上。然后将消毒后的根瘤压碎,并接种在预先准备的YMA 培养基上,并用接种针进行划线分离,然后倒置于28℃恒温培养箱中培养3~5 d。待菌落出现后进行纯化,重复2~3 次,直至分离出单菌落为止,并进行根瘤菌的鉴定。将纯化后的菌株接种于YMB 液体培养基中,并置于摇床中28℃下震荡培养5 d,备用。
1.2 试验设计
1.2.1 不同光照梯度下的接种试验
于4月中、下旬在贵阳市花溪区贵州大学林学院生态试验场进行不同光照梯度下的接种试验。用不同密度的遮阳网控制光照,分别设置100%全光照(G1)、75% 全光照(G2)、50%全光照(G3)、25%全光照(G4)4 个光照处理组。光照相对强度由照度计测定的各处理光照强度与全光照的百分比换算而成。每个处理设置4 个重复,每个重复10 株苗。将长势一致的健康幼苗分别置于不同密度遮荫网下培养7 d 后分别接种4 mL 根瘤菌菌液,后期定期浇灌无菌水,持续处理5 个月。
1.2.2 不同水分梯度下的接种试验
于4月中、下旬在贵阳市花溪区贵州大学林学院试验苗圃塑料大棚中进行不同水分梯度下的接种试验。基质相对含水量(基质含水量占基质田间持水量的百分数)分别设置为80%基质田间持水量(S1)、60%基质田间持水量(S2)、40%基质田间持水量(S3)和20%基质田间持水量(S4)4 个处理组,处理前测得基质田间持水量为30.64%,即4 个处理分别为基质含水量的24.51%、18.38%、12.26%和6.13%。每个处理设置4 个重复,每个重复10 株苗,每株幼苗接种4 mL 根瘤菌菌液,然后置于塑料大棚中培养5 d,每2 d 用称重补水法进行1 次含水量的调节以保持含水量稳定。
1.3 指标测定
1.3.1 幼苗结瘤情况统计
于10月下旬开始收集不同处理下的幼苗根瘤,记录结瘤株数、单株根瘤数量及质量、单个根瘤质量,并计算结瘤率。
1.3.2 生理生化指标测定
于10月下旬开始收集不同处理下的植物叶片、根系以及根瘤,并立即进行生理生化指标的测定(参照张志良等[13]的方法):采用丙酮乙醇混合液(丙酮∶乙醇=1∶1)浸提法测定叶绿素含量;采用活体真空法测定硝酸还原酶含量;采用水杨酸法测定硝态氮含量;采用TTC 法测定根系活力。根瘤中豆血红蛋白含量采用比色法测定[14]。
1.4 数据处理
所有数据均采用Excel 2007 软件进行整理计算并制作图表,采用SPSS 25.0 统计软件进行不同处理指标的方差分析和主成分分析。
2 结果与分析
2.1 不同光照梯度处理对花榈木幼苗接种效应的影响
2.1.1 对花榈木幼苗结瘤数量及质量的影响
由图1~3 可以看出,不同光照处理下,花榈木幼苗接种根瘤菌均能形成根瘤。其中,G2 处理的花榈木幼苗结瘤各项指标与其他处理相比均较高,结瘤率达到了80%,结瘤个数达到105 个,瘤鲜质量为2.68 g,单瘤质量为25.54 mg,分别是G4 的4.5、3.5、7.4、2.1 倍。 其次是G1 处理,结瘤率为60%。最差为G4 处理,结瘤率只有17.5%,表明在75%光照处理下能显著促进花榈木幼苗根系的结瘤数量及质量。
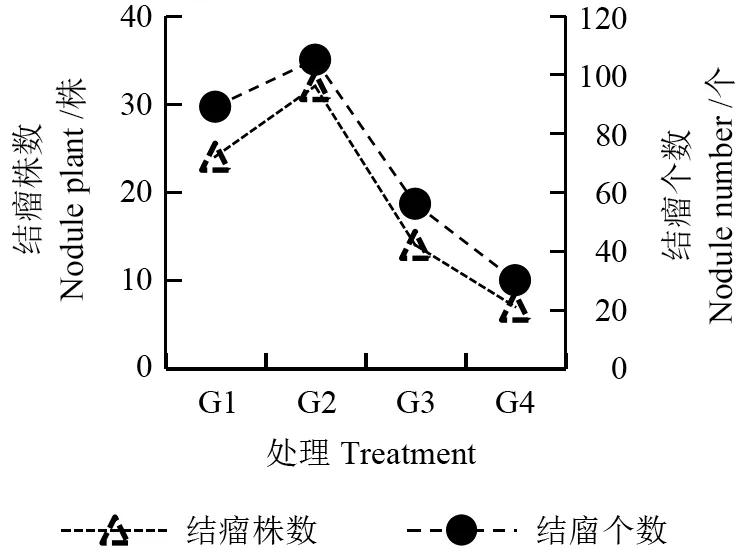
图1 不同光照处理对花榈木幼苗结瘤数量的影响Fig.1 Effects of different light intensity treatments on the number of nodules of Ormosia henryi seedlings

图2 不同光照处理对花榈木幼苗结瘤质量的影响Fig.2 Effects of different light intensity treatments on the quality of nodules of Ormosia henryiseedlings
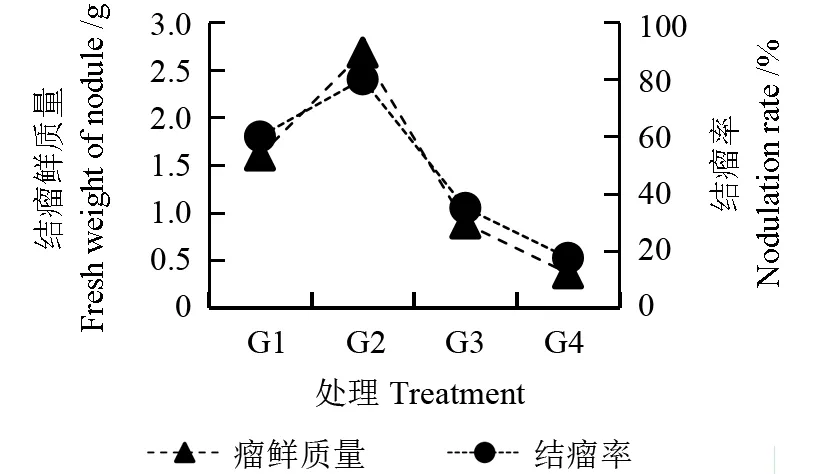
图3 不同光照处理对花榈木幼苗结瘤鲜质量及结瘤率的影响Fig.3 Effects of different light intensity treatments on the fresh weight of nodules and nodulation rates of Ormosia henryi seedlings
2.1.2 对花榈木结瘤幼苗及根瘤生理特性的影响
由表1 可知,不同光照处理下的花榈木结瘤幼苗叶绿素含量、硝态氮含量、硝酸还原酶活性、豆血红蛋白含量以及叶片全氮、全磷、全钾含量差异显著(P<0.05),而根系活力差异不显著(P>0.05)。其中,叶绿素含量、硝酸还原酶活性和全氮含量随着光照强度的降低而增加,以G4 处理下最高,分别为3.47 mg/g、115.94 μg/(g·h)、2.135%。而硝态氮含量、根系活力、豆血红蛋白含量以及全磷、全钾含量随光照强度的降低而呈现出先增高后降低的趋势,以G2 处理的含量最高,其次是G1 处理,最低是G4 处理。结合各项指标,最适宜花榈木幼苗和根瘤生长发育的光照强度是75%全光照。

表1 不同光照强度处理下花榈木结瘤幼苗生理生化特性、根瘤固氮效应及叶片营养元素含量的比较Table 1 Comparison of physiology and biochemical characteristics,nitrogen fixation of nodules and leaf nutrient contents of Ormosia henryiseedlings with nodules under different light intensity treatments
2.1.3 不同光照处理下各指标间相关性分析
由表2 可知,不同光照处理下叶绿素与叶片全氮呈极显著正相关(P<0.01);硝态氮与根系活力呈极显著正相关(P<0.01),与豆血红蛋白呈显著正相关(P<0.05),而其他因子相互之间不具有相关性。说明花榈木结瘤幼苗在不同光强下的植物生理特性、营养元素含量以及幼苗固氮效应等方面虽然存在差异,但它们的一些特性指标与幼苗的结瘤效应相关性不大。

表2 不同光照处理下花榈木结瘤幼苗指标间的相关性分析†Table 2 The correlation analysis of indexes of Ormosia henryi seedlings with nodules under different light intensity treatments
2.1.4 不同光照处理对花榈木幼苗结瘤效应的综合评价
对不同光照强度处理下的花榈木幼苗叶绿素含量、硝态氮含量、硝酸还原酶活性、根系活力、豆血红蛋白含量,以及叶片全氮、全磷、全钾8个指标进行主成分综合评价分析(表3),结果表明:综合得分最高为G2 处理,其次是G1 处理,最低为G4 处理。因此,适合花榈木幼苗结瘤效应的最佳光照强度处理是75%全光照。

表3 不同光照处理对花榈木幼苗结瘤效应的综合评价Table 3 Comprehensive comparison on nodule formation of O.Henryi seedlings with nodules under different light intensity treatments
2.2 不同水分梯度处理对花榈木幼苗接种效应的影响
2.2.1 对花榈木幼苗结瘤数量及质量的影响
由图4~6 可知,不同水分梯度下花榈木幼苗结瘤数量和质量不同。其中S2 的花榈木幼苗结瘤株数最多、每株结瘤个数最多、结瘤率最高,结瘤率达到了70%,结瘤个数达到103 个,瘤鲜质量为2.65 g,分别是S1 结瘤率的5.6 倍、结瘤个数的6.87 倍、瘤鲜质量的7.4 倍。其次是S3 处理,结瘤率为45%。最差为S1 处理,结瘤率只有12.5%。表明在S2 基质水分处理下,能显著促进花榈木幼苗根系的结瘤数量及质量。
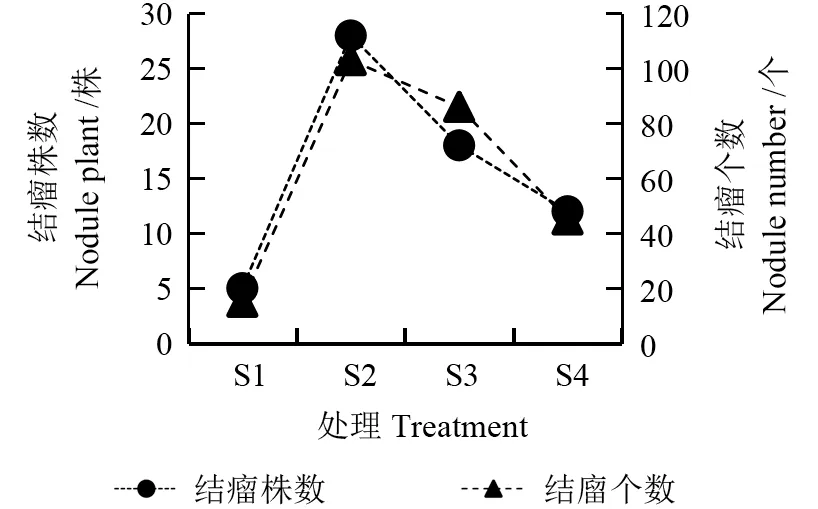
图4 不同水分处理对花榈木幼苗结瘤数量的影响Fig.4 Effects of different substrate water treatments on the number of nodules of Ormosia henryi seedlings
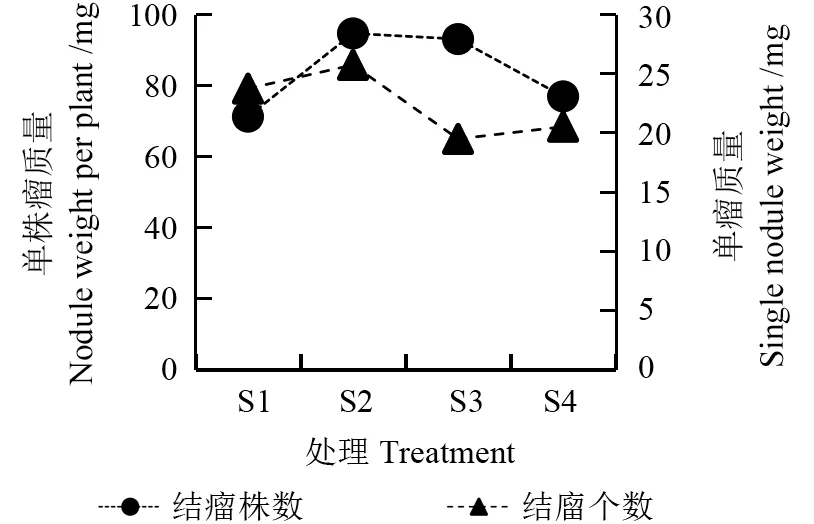
图5 不同水分处理对花榈木幼苗结瘤质量的影响Fig.5 Effects of different substrate water treatments on the quality of nodules of Ormosia henryi seedlings

图6 不同水分处理对花榈木幼苗结瘤鲜重及结瘤率的影响Fig.6 Effects of different substrate water treatments on the fresh weight of nodules and nodulation rates of O.henryi seedlings
2.2.2 对花榈木结瘤幼苗及根瘤生理特性的影响
从表4 可以看出,不同水分处理下的花榈木结瘤幼苗叶绿素含量、硝态氮含量、根系活力、豆血红蛋白含量以及叶片全氮、全磷、全钾含量差异显著(P<0.05),而硝酸还原酶活性差异不显著(P>0.05)。其中,叶绿素含量随着水分含量的降低而降低,S1 最高,为3.00 mg/g;全氮含量随着水分含量的降低而增加,S4 最高,为2.417 mg/g;硝态氮含量、根系活力、豆血红蛋白含量以及全磷、全钾含量随着水分含量的降低而呈现先升高后缓慢降低的趋势,以S2 最高,分别为12.70 mg/g、159.54 mg/(g·h)、126.64 mg/g、0.555%、0.613%;其次是S3,最差为S1。结合各指标而言,基质水分含量过高,对花榈木幼苗和根瘤生理特性的影响较大,不利于花榈木幼苗的生长和结瘤,最适宜花榈木幼苗和根瘤生长发育的基质水分含量是60%基质田间持水量。

表4 不同水分处理下花榈木结瘤幼苗生理生化特性及叶片营养元素含量的比较Table 4 Comparison of physiology and biochemical characteristics,leaf nutrient contents of Ormosia henryi seedlings with nodules under different moisture treatments
2.2.3 不同水分处理下各指标间相关性分析
由表5 可知,不同水分处理下叶绿素与叶片全氮呈显著负相关(P<0.05);硝态氮与豆血红蛋白呈极显著正相关(P<0.01),与根系活力、叶片全磷含量呈显著正相关(P<0.05);根系活力与豆血红蛋白含量、叶片全磷含量呈显著正相关(P<0.05);豆血红蛋白含量与叶片全磷含量呈显著正相关(P<0.05);其他因子之间不具有相关性,说明花榈木结瘤幼苗在不同水分处理下的叶片全磷含量与硝态氮含量、根系活力、豆血红蛋白含量均存在相关性,硝态氮含量与全磷含量、根系活力、豆血红蛋白含量均存在相关性,缺磷和硝态氮均会影响根系的生长和根瘤菌的固氮能力。
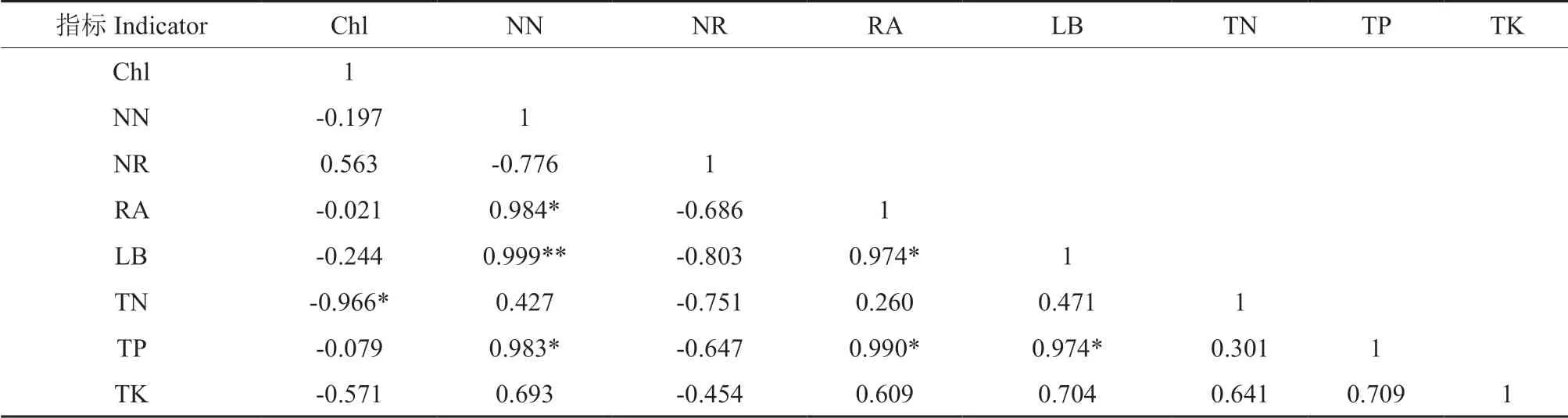
表5 不同水分处理下花榈木结瘤幼苗指标间的相关性分析Table 5 The correlation analysis of indexes of Ormosia henryi seedlings with nodules under different moisture treatments
2.2.4 不同水分处理对花榈木幼苗结瘤效应的综合评价
对不同水分处理下的花榈木幼苗叶绿素含量、硝态氮含量、硝酸还原酶活性、根系活力、豆血红蛋白含量,以及叶片全氮、全磷、全钾8 个指标进行主成分综合评价分析(表6),结果表明:综合得分最高为S2 处理,其次是S3 处理,综合得分最低为S1 处理。因此,适合花榈木幼苗结瘤效应的最佳水分处理是60%基质田间持水量。
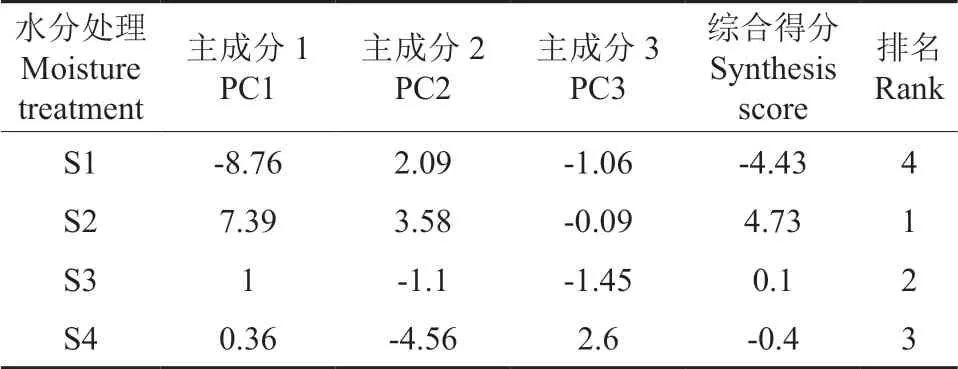
表6 不同水分处理对花榈木结瘤幼苗结瘤效应的综合评价Table 6 Comprehensive comparison of nodule formation of O.Henryi seedlings with nodules under different moisture treatments
3 讨论与结论
3.1 讨论
通过模拟不同光照强度和基质含水量,研究其对接种根瘤菌的花榈木幼苗结瘤效应和生理生化指标的影响。结果表明,不同的光照强度和水分对花榈木结瘤幼苗的结瘤数、瘤鲜质量、结瘤率、叶绿素含量、硝态氮含量、硝酸还原酶活性、根系活力、根瘤中豆血红蛋白含量以及叶片营养元素等指标均有不同程度的影响。
根瘤是生物固氮的主要场所,根瘤的数量和质量是评判根瘤固氮能力强弱的直观表现指标[15]。光对植物来说是主要的环境因子,也是光合作用的重要能量来源。不同的光照条件不仅影响植物的生长发育、内部物质的合成与代谢,甚至对于某些豆科植物来说,光照强度在幼苗的存活方面起着关键性的作用,如高山豆、唐古特岩黄耆Hedysarum tanguticum和天蓝苜蓿,光强过低可能会导致这3 个物种的幼苗大量死亡[16]。段如雁等[17]通过人工模拟不同光照强度对花榈木幼苗的生态适应性研究发现,75%光强下的花榈木幼苗生长发育最好,同时也说明无论接种根瘤菌与否,75%的全光照处理对花榈木幼苗的生态适应性指标和结瘤效应指标均最好。周相娟等[18]也验证了遮光会直接限制植株生长和根瘤发育,大大减少根瘤数目和质量,同时固氮酶活性也显著降低,但是随着遮光程度的加重,植物叶片中的叶绿素总量、叶片总氮量增加。研究结果表明,随着光照的减弱,植物内部硝态氮含量、根系活力、根瘤豆血红蛋白含量以及叶片全磷、全钾均呈现先升高后降低的趋势,而叶绿素含量、硝酸还原酶活性和全氮含量呈现升高的趋势。导致这个结果的原因可能是由于高光强和弱光胁迫下均易导致植物体内发生氧化损伤,细胞易受到伤害[19-20],导致根系活力下降,幼苗光合作用易受到抑制,进而影响光合作用下的能量产出,从而影响根瘤菌的固氮效果、根系活力以及结瘤效果[21]。且遮光后叶中N的再吸收速率一般比碳的同化速率要慢得多,可能会增加N 向植物叶组织的分配,从而造成叶片中硝酸还原酶活性和全氮含量升高。随着光照强度的降低,叶绿素含量逐渐增加,说明植物可以通过提高叶绿素的含量来增强对弱光的适应能力。一般而言,随着光强的减弱,植物为了能更好地利用环境光能以对生长进行补偿,通常叶绿素含量会增加[22]。在弱光环境下的光以短波为主,而叶绿素b 能有效吸收短波光,叶绿素b 的增加有利于LHCP(捕光色素复合体)含量的增加,使类囊体膜垛叠形成基粒并调节激发能在PSI 与PSII之间的均衡分配[23],从而使叶绿素含量增加,这与前人研究结果一致。
关于水分对豆科植物结瘤和固氮能力的影响研究,李志芳等[24]发现干旱胁迫会影响植株生物量的积累和根瘤菌的固氮能力,当根瘤形成之后,土壤干旱会导致N2生物固定量降低,当土壤湿度持续降低时,N2固定几乎停止。姜顺邦等[25]对花榈木容器苗水分需求规律的研究中发现,在80%田间持水量下,花榈木容器苗生理生化指标受水分胁迫的影响较小。本研究结果是以60%基质田间持水量为宜,过高和过低均不利于其生长。原因可能是前者未接种根瘤菌,而本试验主要是探究接种根瘤菌后的花榈木幼苗的结瘤效应,水分过高可能会使植物体内代谢紊乱,SOD 基因表达受阻,无法清除体内过多超氧自由基,从而导致植物氧化损伤程度加深并释放有毒物质[26]。还有可能是根系缺氧造成类菌体呼吸和固氮所需氧供应量不足,进而导致根瘤菌的固氮效应降低[27]。同时水分过高或过低可能会导致病害的发生,使其死亡率升高。随着水分含量的降低,叶绿素含量随之降低,说明缺水会影响光合作用的正常运行,而叶绿素a 和叶绿素b 是植物重要的光合色素,因此缺水会抑制光合色素的生物合成,从而造成叶绿素含量降低。这与董斌等[28]的研究结果一致。
豆血红蛋白(Leghemoglobin,Lb)是根瘤中最丰富的一类蛋白,约占成熟根瘤总蛋白含量的5%以上,可以调控根瘤高效固氮。有研究表明,豆血红蛋白缺失突变体会导致早期的根瘤快速老化,影响血红素和蛋白酶的合成,同时豆血红蛋白的缺失会导致根瘤超微结构和活性氧的严重改变,导致植物体内碳和氮代谢相关基因发生剧烈变化[29],因此豆血红蛋白对促进根瘤高效固氮具有重要作用。不同光照和不同水分处理下各指标间相关性分析结果表明:花榈木结瘤幼苗在不同光照处理下的硝态氮含量与根系活力呈极显著正相关(P<0.01),与豆血红蛋白呈显著正相关(P<0.05),说明在弱光下,硝态氮的供应可以促进根系的生长以及促进根瘤的固氮能力。花榈木结瘤幼苗在不同水分处理下的叶片全磷含量与硝态氮含量、根系活力、豆血红蛋白含量均存在正相关性,且硝态氮含量与全磷含量、根系活力、豆血红蛋白含量均存在正相关性,说明磷和硝态氮均会影响植物的生长发育和根瘤菌的固氮能力。因此建议在弱光环境和干旱胁迫下的花榈木结瘤幼苗,人为补充适量的有效磷或硝态氮,以应对不同环境胁迫下的危害[30-31],这在前人研究中已被证实。
从结瘤效应综合评价结果可以看出,G2 和S2处理下的多数指标均表现优异,初步认定G2 和S2 单一因素处理下比较适宜花榈木结瘤幼苗的生长。但对结瘤效应的综合评价还需要对花榈木幼苗各生长指标、形态指标以及光合指标等进行观测,同时还需对每个处理中的根瘤菌进行长期定位观测,观察并探究根瘤菌株定殖情况、分布情况、种群丰度以及耐受性等因素,全方位指标分析可为花榈木幼苗在人工栽培中提供更加全面的理论依据。
3.2 结论
1)75%全光照处理和60%基质田间持水量处理花榈木幼苗均可显著促进其结瘤,结瘤数量及质量均达到最大值,结瘤率分别达到了80%和70%。
2)75%全光照处理和60%基质田间持水量处理下的花榈木结瘤幼苗,其根瘤内部豆血红蛋白含量以及幼苗硝态氮含量、根系活力,叶片全磷、全钾含量均显著提高,其中叶绿素含量随着光照强度的减弱而增加,随着基质水分的降低而减少。
3)综合评价结果表明,不同光照和水分处理对花榈木幼苗结瘤效应的综合评价由大到小分别是:G2 >G1 >G3 >G4;S2 >S3 >S4 >S1,分别以75%全光照和60%基质田间持水量为最佳。
猜你喜欢
山东冶金(2022年1期)2022-04-19 13:40:14
中国南方果树(2022年1期)2022-01-28 07:39:16
农技服务(2021年7期)2021-09-24 04:13:02
上海金属(2021年4期)2021-07-28 12:33:22
园艺与种苗(2021年1期)2021-03-16 03:01:08
上海金属(2020年6期)2021-01-04 12:25:52
河北果树(2020年1期)2020-02-09 12:31:36
植物营养与肥料学报(2019年11期)2019-12-13 05:55:04
西藏农业科技(2019年3期)2019-11-04 00:35:08
河南科技(2014年8期)2014-02-27 14:07:58
