
副干酪乳杆菌LPC-F通过促进Cajal间质细胞增殖缓解便秘
2022-02-25 08:24王琳琳杨树荣王嘉良王刚张灏赵建新陈卫
食品与发酵工业 2022年4期
王琳琳,杨树荣,王嘉良,王刚,张灏,赵建新,陈卫
(江南大学 食品学院,江苏 无锡,214122)
便秘是一种复杂的胃肠道功能障碍综合征。主要症状为自发排便次数减少、排便困难、大便干硬、排便量少。便秘的发生率相对较高,据统计,全世界儿童便秘的发生率为0.5%~29.6%,成人便秘的发生率为2.6%~31%,老年人便秘的发生率为4.4%~ 44.5%[1]。研究表明便秘与肠道微生物群之间存在很强的相关性。与野生型小鼠相比,无菌小鼠肠蠕动明显弱于野生型小鼠,将SPF级小鼠粪便移植给无菌鼠后,肠蠕动能力显著提高[2],这说明肠道菌群与肠蠕动有非常密切的关系。将便秘人群的粪便移植给无菌鼠后,无菌鼠结肠的平滑肌变弱,肠蠕动能力减弱[3]。上述结果进一步阐明了肠道菌群异常与便秘之间存在密切联系。便秘患者中厚壁菌门和拟杆菌门的相对丰度与正常人有较大差异[4]。
副干酪乳杆菌(Lactobacillusparacasei)是一种广泛存在于人肠道以及奶酪、泡菜等发酵食品中的益生菌,是维持人类微生态平衡的重要成员。前人的研究[5]发现副干酪乳杆菌LC-01,NTU 101及LC19均能不同程度的缓解由洛哌丁胺诱导的小鼠便秘症状。同时热灭活的副干酪乳杆菌LC19也具有良好的缓解便秘的效果。上述的研究都仅限于副干酪乳杆菌是否具有缓解便秘作用这种疗效指标上,并未对其缓解便秘的机制进行解析。基于此,本文选取不同来源的对便秘具有不同缓解效果的2株副干酪乳杆菌(健康成人粪便和泡菜),对其进行缓解便秘的可能机制的解析,主要研究内容包括:考察不同来源的2株副干酪乳杆菌对小鼠便秘症状的缓解情况;从c-kit基因、短链脂肪酸(short-chain fatty acid,SCFAs)、神经递质[5-羟色胺(5-hydroxytryptamine,5-HT)、P物质]、肠道病理损伤、炎症因子以及肠道菌群等检测指标,进一步探究副干酪乳杆菌改善便秘的相关作用途径;应用相关性分析的方法,建立肠道菌群和短链脂肪酸间的相关性,最终确定副干酪乳杆菌缓解便秘的可能机制。
1 材料与方法
1.1 实验材料
1.1.1 实验试剂
免疫组化试剂盒,生物工程(上海)股份有限公司;小鼠c-kit蛋白抗体,北京博奥森生物技术有限公司;小鼠白细胞介素-1β(IL-1β)、小鼠肿瘤坏死因子-α(TNF-α)ELISA试剂盒,武汉酶免生物科技有限公司;盐酸洛哌丁胺(2 mg),西安杨森制药有限公司;小鼠5-HT、小鼠P物质(SP)ELISA试剂盒,上海酶联生物科技有限公司;乙酸、丙酸、丁酸、戊酸、硫酸、无水乙醇、无水乙醚、无水硫酸钠、甲醇、30%过氧化氢,国药集团化学试剂公司。
活性炭溶液的配制:将阿拉伯胶100 g与水800 mL用玻璃棒混合均匀,放置于红外炉上煮沸至溶液透明,加入活性炭50 g,煮沸后取下,待混合液稍冷却再继续煮沸,重复3次,制成活性炭悬浊液。溶液冷却后,用水稀释至1 000 mL,贮存于4 ℃,用前摇匀。
1.1.2 实验菌株
将保存于-80 ℃冰箱中的菌株取出,以2%的接种量接种至MRS液体培养基中,37 ℃厌氧培养箱中培养24 h。连续活化2代后,以同样接种量接种至900 mL MRS液体培养基中进行扩培,同样条件培养后,将菌液以8 000×g离心15 min,弃上清液,得副干酪乳杆菌菌泥。用预冷生理盐水清洗菌泥,以7 000×g离心15 min,弃上清液,重复清洗3次,获得干净的菌泥。配制30%(体积分数)甘油,并按照菌泥∶30%甘油=1∶1(g∶mL)将甘油与菌泥混合均匀后进行分装,贮存于-80 ℃冰箱中。每隔7 d对菌液进行活菌计数,结果表明冻存1个月不会引起副干酪乳杆菌活菌数量级的变化,因此动物实验中每月制备1次菌液。
在灌胃当天,取出冻存的菌液,以6 000 r/min离心5 min,弃上清液,获得菌泥,用生理盐水清洗,以6 000 r/min离心5 min,弃上清液,重复清洗3次,获得干净的菌泥。向菌泥中添加一定体积的3%蔗糖溶液,使菌液浓度达到109CFU/mL,混合均匀后置于冰上备用。实验中所用的2株副干酪乳杆菌均来自江南大学食品生物技术中心菌种保藏中心,见表1。

表1 实验用菌株Table 1 Experimental strains
1.1.3 动物实验
动物实验选用40只6周龄BALB/c雄性小鼠,购自南京模式动物研究所,饲养于江南大学实验动物中心(许可证编号:SYXK2016-0045)的屏障环境中[室内温度(25±2)℃;湿度(50±5)%;昼夜循环12 h]。实验过程严格遵守江南大学实验动物中心相关管理规定,小鼠在饲养期间饲喂标准饲料,自由饮水。将所有小鼠随机分为空白组、模型组、副干酪乳杆菌LPC-F组和LPC-V干预组,每10只随机分为1组,共计4组。动物实验主要包括适应期、干预期与造模期,共计45 d。第1~7天为适应期,小鼠进驻动物房后适应性饲养1周。从第8~28天,每天8:00正常对照组、便秘模型组灌胃0.2 mL 3%蔗糖溶液、菌处理组灌胃相应的3%蔗糖溶液重悬的菌悬液(5×109CFU/mL)0.2 mL。从第29~37天,每天上午,模型组、菌处理组灌胃0.2 mL盐酸洛哌丁胺悬液(10 mg/kg BW),正常组灌胃0.2 mL蒸馏水,1 h后,正常组、模型组灌胃0.2 mL 3%蔗糖溶液,不同菌处理组灌胃0.2 mL 相应的3%蔗糖溶液重悬的菌悬液(5×109CFU/mL)。实验期间各组小鼠均饲喂标准维持饲料,从第0天开始,每周收集1次小鼠粪便,并于最短时间内保存于-80 ℃冰箱。实验结束后(第45天),按0.05~0.1 mL/10 kg,采用1%戊巴比妥钠溶液对小鼠进行腹腔注射,用镊子进行眼部静脉丛采血,采血完毕后脱颈椎处死。收集到的血液在室温条件下静置2 h,3 000×g条件下离心10 min,将血清进行分装,-80 ℃冻存备用,实验分组见表2。
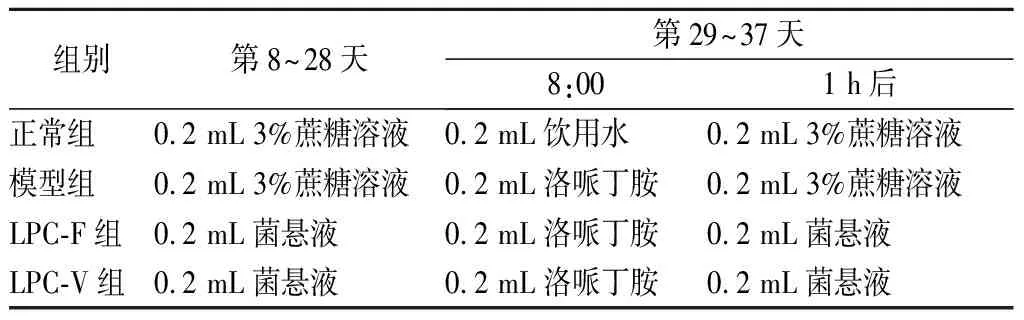
表2 动物实验分组Table 2 The group of animal experiment
1.1.3.1 排首粒黑便时间检测
实验第37 天,进行小鼠排首粒黑便时间测定。模型组小鼠和菌处理组小鼠灌胃0.2 mL盐酸洛哌丁胺悬液,正常组小鼠灌胃0.2 mL蒸馏水,1 h后,各组每只小鼠灌胃0.2 mL上述的活性炭溶液,记录每只小鼠灌胃结束后的时间,并将其放入干净的笼盒中,等其排出首粒黑便,然后记录排出黑便时所对应的时间。如上2个时间段的间隔时间即为排首粒黑便时间。
1.1.3.2 小肠推进率检测
实验最后1 d,对小鼠小肠推进率进行测定。第37天19:00禁食不禁水过夜,第38天8:00,便秘模型组和菌处理组小鼠灌胃0.2 mL盐酸洛哌丁胺悬液,正常组灌胃0.2 mL蒸馏水,1 h后,按照相应的顺序对每组小鼠灌胃活性炭溶液。30 min后用戊巴比妥钠溶液进行麻醉,对小鼠进行眼眶采血后,断颈牺牲。轻轻剪开小鼠腹腔,取出从幽门至盲肠段,用镊子在测量尺上拉直,记录小肠全长和活性炭前沿至幽门的距离。小肠推进率计算如公式(1)所示:

(1)
1.1.3.3 粪便含水量检测
每周上午8:00收集粪便,方法如下:先给每只小鼠灌胃盐酸洛哌丁胺,1 h后将小鼠分别单独放置在事先垫有滤纸的干净笼盒中,从放入笼盒的时刻开始计时,6 h后将小鼠轻轻放回饲养笼盒,期间收集每只小鼠的粪便。将收集到的粪便装入事先灭菌的1.5 mL EP管中,然后将EP管置于冰盒中保存。收集到的粪便经冷冻干燥后,记录粪便的干重,粪便的含水量计算如公式(2)所示:

(2)
1.1.3.4 血清中炎症相关指标及结肠组织中便秘相关胃肠调节肽的检测
小鼠血清用于检测TNF-α、IL-1β的浓度。检测方法参照相应ELISA试剂盒说明书进行。结肠组织用于5-HT、SP的检测。使用预冷的生理盐水轻轻冲洗结肠组织,去除组织中残留的杂质和周围的脂肪,剪取适当质量的结肠组织,准确称重,按照组织∶生理盐水=1∶9(g∶mL)的比例加入预冷的生理盐水,使用高通量组织研磨器,以70 Hz/s的频率研磨30 s,共振荡4次,直至组织不存在肉眼可见的大颗粒物质。12 000×g、4 ℃离心15 min,分装上清液,按照5-HT、SP的ELISA试剂盒说明书进行测定。
1.1.3.5 结肠组织中c-kit蛋白免疫组化
通过免疫组化染色评估结肠组织中c-kit蛋白,按照免疫组化试剂盒进行实验。具体操作步骤如下:(1)为防止组织脱片,烤片时间缩短为1 h;(2)不同浓度的乙醇进行梯度水化时,将水化时间由3 min提高至5 min。封片后,经自然风干,用无水乙醇或二甲苯擦去多余树胶,用切片扫描仪进行观察拍照。免疫组化结果使用Image J软件结合IHC profile插件对免疫组化切片阳性表达面积进行统计。每个切片随机选取6个区域进行统计。
采用实时荧光定量聚合酶链反应(real-time fluorescent polymerase chain reaction,RT-qPCR)来测定c-kit基因转录水平。取超低温冰箱冻存新鲜结肠组织,按说明书用Trizol法提取总RNA。按照反转录试剂盒说明书方法合成cDNA。c-kit基因、GAPDH基因引物序列如表3所示。RT-qPCR反应体系共10 μL。反应条件为:95 ℃,2 min;95 ℃,30 s;59 ℃,30 s;72 ℃,20 s;共35个循环。以GAPDH基因为内参基因,CFX96Manager软件分析结果。

表3 c-kit基因、GAPDH基因引物序列Table 3 Primer sequence of c-kit gene and GAPDH gene
1.1.3.6 粪便中SCFAs检测
参考毛丙永[6]的方法采用GC-MS测定小鼠粪便中SCFAs含量,将收集到的粪便经冻干、浸泡、酸化、乙醚萃取并脱水后利用Thermo气质联用仪测定SCFAs的含量。
1.1.3.7 粪便菌群检测
粪便DNA提取是按照Fast DNA SPIN Kit for Feces试剂盒标准流程操作。具体操作步骤如下:
(1)提取粪便样品中总DNA
应用MP粪便样品基因组提取试剂盒Fast DNA SPIN Kit for Feces,提取总DNA。
(2)16S rDNA可变区(V3~V4区)扩增
以粪便基因组为模板,以正向引物341F(5′- CCT AYG GGR BGC ASC AG-3′)、反向引物802R (5′- GGA CTA CNN GGG TAT CTA AT-3′)为引物,扩增16S rDNA V3~V4区片段,不同样品用7个碱基组成的barcode进行区分,并加在上游引物341F的5′端,其中目的片段长度约为465 bp左右。
(3)PCR产物切胶回收
按照 QIAquick Gel Extraction Kit 胶回收试剂盒说明书进行目的条带胶回收。
(4)混样、构建文库以及Illumina Miseq平台测序
参照TruSeq Nano DNA LT Sample Prep Kit建库试剂盒的说明书构建文库,完成末端修复、3′端加A、接头连接以及PCR扩增等一系列步骤。文库质控采用Agilent Bioanalyzer 2100生化分析仪测定DNA片段的分布情况,qPCR精确定量文库中DNA的质量浓度(ng/μL)。最后参照测序试剂盒MiSeq Reagent Kit v3的说明书将文库加至Reagent Kit Cartridge中,放入MiSeq测序仪中开始上机测序。
(5)下机数据质量控制
质控方法参考文献[7]。测序完成后,根据引物Barcode特异性拆分文库中对应样品的原始序列,过滤掉barcode和引物序列、低质量碱基序列(质量得分< 30),序列小于200 bp的序列和错配序列。过滤后的序列在QIIME软件进行正反向序列拼接,要求最小重叠区域为10 bp、重叠区域的最大错配比率为0.2、去除嵌合体序列,最终得到高质量拼接序列。采用VSEARCH软件对得到的优质拼接序列进行uclust聚类分析,根据序列相似性生成大量的操作分类单元(operational taxonomic unit,OTU),相似度 ≥ 97%的可认为是1个OTU。RDP classifier软件对OTUs代表序列进行分类学分析,与Silva参考数据库比对注释,得到微生物在不同水平上的分类学信息,进一步分析样本的α多样性和β多样性。Alpha多样性指数包括Chao 1、Simpson指数。FastTree软件构建OTU代表序列的系统发育树,进行层次聚类,计算样品间的UniFrac距离分析样品间相似性和差异性,体现不同样本微生物组成和群落结构差异(β多样性)。结合OTU的分类和丰度,进一步计算weighted Unifrac和unweighted Unifrac距离。
1.1.4 统计分析
本论文中,数据统计学方法均用“平均值±标准差”(mean±SD),使用GraphPad Prism 9.0版进行绘图。采用SIMCA、OmicShare tool在线绘图网站、Past3进行主成分分析(principal component analysis,PCA)绘图。使用SPSS(22版)进行One-Way ANOVA单因素方差分析及相关性分析。采用Duncan进行事后检验。对各组数据的差异进行分析,以不同字母标注表示有显著差异(P<0.05)。通过PCA,中心对数比(clr)变换的属水平计数表以及方差的排列多变量分析(perMANOVA)对β多样性进行了评估。通过线性判别分析(linear discriminant analysis,LDA)效应大小(LDA effect size,LEfSe)区分微生物生物标志物(lgLDA>2.0作为阈值)。采用TSS方式对数据进行标准化,并在MicrobiomeAnalyst中分析α多样性和LEfSe(https://www.microbiomeanalyst.ca/faces/home.xhtml)。
2 结果与分析
2.1 副干酪乳杆菌LPC-F 具有缓解便秘的作用
由图1可知,小鼠灌胃洛哌丁胺后,粪便含水量和小肠推进率显著下降,排首粒黑便时间显著增加,表明便秘模型造模成功。应用不同的副干酪乳杆菌干预后,LPC-F组显著改变了便秘相关的指标,而LPC-V组仅对排首粒黑便时间有效,结合《保健食品评价技术规范》中有关缓解便秘阳性结果的评价标准,提示L.paracaseiLPC-F具有缓解便秘的作用。以往的研究发现青春双歧杆菌在缓解便秘方面具有种间差异,其产生差异的原因可能与菌株的黏附特性及菌株的生长速率相关[8]。因此下一步可以从菌株的生理特性方面去分析这2株副干酪乳杆菌在缓解便秘方面具有差异的可能原因。
a-粪便含水量;b-小肠推进率;c-首粒黑便时间图1 副干酪乳杆菌对小鼠便秘相关指标的影响Fig.1 The effect of L. paracasei on constipation-related indicators in mice 注:#, 模型组与正常组之间具有显著性差异, 其中#表示P<0.05,###P<0.005, ####P<0.001; *,菌干预组之间或菌干预组与模型组之间具有显著性差异,其中*表示P<0.05,**P<0.01,***P<0.005,****P<0.001; ns, 两组间无显著差异(下同)
2.2 副干酪乳杆菌LPC-F可降低小鼠的炎症水平
IL-1β可以引导炎症细胞进入病变部位,促进血管白细胞黏附分子的表达,引起炎症反应[9]。而TNF-α主要在炎症初期诱导多种细胞产生其他炎症因子,产生重要的协同作用。在本研究中(图2),模型组小鼠血清中的TNF-α和IL-1β的水平显著升高(P<0.05),证明便秘同时也伴有低水平的炎症,这与MACKOS等[10]的发现相一致。相较于模型组,2株副干酪乳杆菌均能显著下调促炎因子TNF-α和IL-1β的水平(P<0.05),改善了便秘所引起的机体的炎症水平。乳杆菌在缓解炎症,调节机体TNF-α水平方面的研究很多,且相关菌株调节免疫系统的功能也得到验证。例如罗伊乳杆菌BM36301可降低老年鼠体内TNF-α水平[11],也有研究指出副干酪乳杆菌能降低宿主TNF-α的水平[11],这与本研究中副干酪乳杆菌可缓解便秘所引起的机体炎症水平的结果相一致。结合2株副干酪乳杆菌对便秘相关指标的影响情况,可知L.paracaseiLPC-F在缓解便秘的同时,下调了机体的炎症水平,而L.paracaseiLPC-V虽能下调机体的炎症水平,但对便秘无缓解作用。因此,下调炎症因子水平可能不是2株副干酪乳杆菌具有缓解便秘差异的原因。

a-SP;b-TNF-a图2 副干酪乳杆菌对小鼠血清中炎症因子水平的影响Fig.2 The effect of L.paracasei on levels of inflammatory factors in mice
2.3 副干酪乳杆菌LPC-F可提高结肠组织中胃肠调节肽5-HT和SP的水平
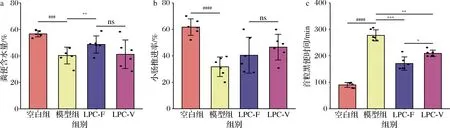
有关5-HT对肠动力影响的研究已超过50余年[12],临床上已有5-HT4受体激动剂(商品名:西沙比利)作为促动力药用于治疗便秘。因此本文对结肠内5-HT的水平进行了检测(图3),模型组5-HT的水平显著下调,这与先前的研究结果一致[13]。LPC-F组小鼠结肠中5-HT的水平显著高于模型组,提示L.paracaseiLPC-F进入宿主体内后,可以通过其自身的代谢产物或者通过改善肠道菌群的组成提高宿主体内5-HT的水平,从而缓解结肠动力减弱的症状。然而也有研究发现,不同类型的便秘患者5-HT的变化趋势截然不同[14],这可能是因为5-HT需要通过激活相应的5-HT受体才能发挥作用。同时,受血清素转运蛋白(serotonin transporter,SERT)的影响,5-HT可被转运灭活,从而终止反应[14]。这就提示我们仅测量组织中5-HT的水平,还不够准确,还需要测定肠道中5-HT受体及SERT表达情况,才能更加准确的解析L.paracaseiLPC-F对5-HT信号分子相关通路的影响情况。
SP是胃肠道一种重要的兴奋性神经递质,研究表明,肠道菌群会影响宿主体内SP的水平[15]。模型组小鼠结肠SP水平显著低于正常组小鼠。LPC-F组小鼠结肠中SP的水平显著高于模型组,提示L.paracaseiLPC-F缓解便秘可能与其上调宿主体内SP水平有关。
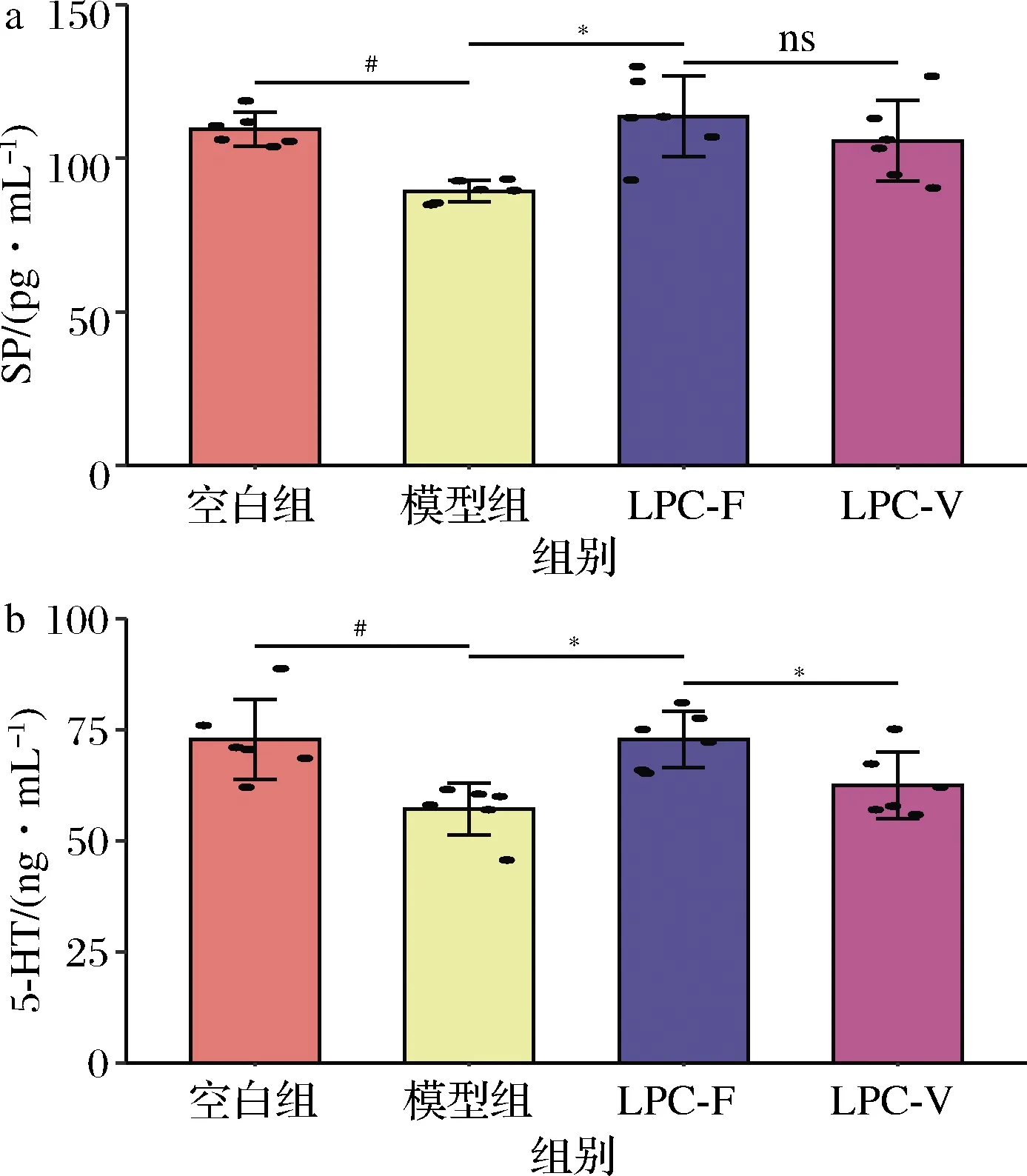
a-SP;b-5-HT图3 副干酪乳杆菌对小鼠血清中胃肠动力调节激素水平的影响Fig.3 The effect of L.paracasei on levels of gastrointestinal motility hormone in mice serum
2.4 副干酪乳杆菌LPC-F显著提高了结肠Cajal间质细胞(interstitial cells of Cajal, ICC)的数量
已有大量文献证明,ICC数量的减少以及形态变化是导致胃肠道功能障碍的原因之一。酪氨酸激酶受体蛋白c-kit蛋白与ICC的功能密切相关,敲除c-kit将导致ICC失去功能,因此,可以将c-kit作为ICC的标记物[16]。免疫组化染色结果如图4所示,模型组小鼠c-kit蛋白阳性信号强度弱于正常组小鼠,提示模型组小鼠ICC的数量显著低于正常组小鼠。LPC-F组小鼠结肠中c-kit蛋白的阳性信号较强,呈较深的黄棕色,提示LPC-F组小鼠ICC数量高于模型组。由于免疫组化染色只能进行样品的半定量,于是本研究进一步对c-kit基因转录水平进行测定,结果如图4-b所示,模型组小鼠结肠c-kit基因的相对表达量下调,LPC-F组小鼠结肠中c-kit基因转录水平显著高于模型组,提示L.paracaseiLPC-F 缓解便秘的同时促进了ICC数量的恢复。
2.5 副干酪乳杆菌LPC-F提高粪便中乙酸、丙酸水平
SCFAs是肠道细菌发酵多糖产生的重要代谢产物。本研究主要针对3种肠道中含量较为丰富的SCFAs展开测定,结果如图5所示。模型组小鼠粪便中的乙酸、丙酸、丁酸的含量显著下降(P<0.05)。临床研究也发现,便秘患者粪便中乙酸、丙酸、丁酸含量较正常人也有显著下调的现象[17],这与我们的研究结果相一致。L.paracaseiLPC-F干预后,小鼠粪便乙酸、丙酸的含量显著高于模型组,提示L.paracaseiLPC-F缓解便秘可能与其粪便中乙酸、丙酸的含量相关。有文献指出,乙酸可以刺激结肠释放5-HT,使得结肠蠕动增加1倍[18]。结合L.paracaseiLPC-F缓解便秘及对5-HT的影响,推测L.paracaseiLPC-F可能通过提高结肠中乙酸、丙酸的含量,从而刺激肠嗜铬细胞分泌5-HT促进排便。

a-免疫组化染色;b-c-kit相比表达量图4 副干酪乳杆菌对ICC数量的影响Fig.4 The effect of L.paracasei on the number of ICC

a-乙酸;b-丙酸;c-丁酸图5 副干酪乳杆菌对小鼠粪便短链脂肪酸的影响Fig.5 The effect of L.paracasei on short-chain fatty acids in mice feces
2.6 副干酪乳杆菌LPC-F对便秘小鼠肠道菌群的影响具有其独特性
肠道菌群通过黏附在肠黏膜上,形成了又一类肠道屏障——生物屏障[19]。一方面,生物屏障与宿主存在共生关系,两者之间能够进行营养物质、激素以及信号分子的传递,参与调控肠道免疫系统。另一方面,这些肠道菌群能够抵抗致病菌的侵袭与伤害,维持菌群生态平衡。根据已有研究,便秘的发生与肠道菌群的紊乱相关,同时,恢复肠道菌群稳态有益于缓解便秘[19]。因此,探究便秘患者肠道菌群的变化与双歧杆菌对肠道菌群的影响是十分必要的。
2.6.1 对肠道菌群多样性的影响
本研究通过Chao1指数及Simpson指数衡量各组小鼠粪便菌群的α-多样性。Chao 1指数如图6-a、图6-b所示, 便秘小鼠粪便菌群的Chao1指数显著下降而Simpson指数显著上调(P<0.05),这表明洛哌丁胺作用后,小鼠粪便中微生物的丰富度减小,粪便中可观测到的微生物种类变少。2株副干酪乳杆菌干预后小鼠粪便菌群Chao1指数显著高于模型组小鼠,提示2株副干酪乳杆菌可以显著提高粪便中物种的丰富度。
研究证实,便秘人群肠道菌群β-多样性显著区别于正常人群[4]。本研究也同样考察了小鼠肠道菌群β-多样性,根据Bray-Curtis距离进行主坐标分析(principal coordinates analysis,PCoA),组间比较采用多元方差分析,结果如图6-c所示,PCoA图显示模型组小鼠的数据点和正常组小鼠的数据点出现了较大的偏移,说明洛哌丁胺对小鼠粪便菌群结构已产生了极为显著的影响。LPC-F组的β-多样性与模型组有偏差,而LPC-V的β-多样性与模型组相似。
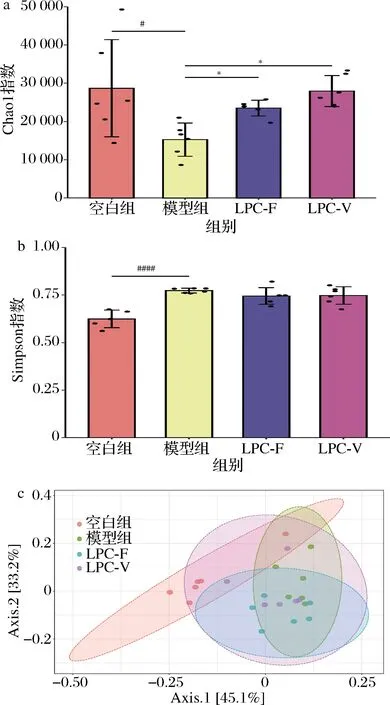
a-副干酪乳杆菌对小鼠粪便菌群Chao 1指数的影响; b-副干酪乳杆菌对小鼠粪便菌群Simpson指数的影响; c-副干酪乳杆菌对小鼠粪便菌群β-多样性的影响图6 副干酪乳杆菌对小鼠粪便菌群多样性的影响Fig.6 The effect of L.paracasei on the diversity of intestinal microbiota in mice
2.6.2 对肠道菌群在门水平上的影响
门水平上(图7-a),主要由Firmicutes、Bacteroidetes、Proteobacteria和Actinobacteria 4个门组成。洛哌丁胺作用后,小鼠粪便中Bacteroidetes相对丰度下降12%左右,而Firmicutes的相对丰度则相对应提高12.8%左右。模型组小鼠肠道菌群中Verrucomicrobia的相对丰度也显著提高,这与BOUDREAU等[20]在长期灌胃芦荟大鼠中观察到的结果一致。模型组小鼠粪便中Deferribacteres相对丰度是正常组小鼠的3倍之多。在LI等[21]的研究中也发现便秘小鼠粪便中Deferribacteres相对丰度有上调的现象。副干酪乳杆菌干预后逆转了便秘所引起的Bacteroidetes与Firmicutes相对丰度的改变,而肠道微生物群组成改变的一大主轴是沿着拟杆菌门-厚壁菌门轴进行的,拟杆菌门/厚壁菌门的结果与α-多样性可能存在一定相关性[22]。
2.6.3 对肠道菌群在属水平上的影响
进一步对小鼠粪便菌群数据展开LEfSe分析,得到了在小鼠肠道中具有统计学差异的生物标识(biomarker)(LDA值>2)。结果如图7-b所示,模型组肠道菌群的特征属为Ruminococcus和Allobaculum。LPC-F组的特征属为Lactobacillus、Bacteroides及Akkermansia。而LPC-V组的特征属为Anaeroplasma。
根据LEfSe分析结果,对相关属的细菌相对丰度进行汇总统计,结果如图8-a所示,便秘小鼠粪便菌群中Lactobacillus和Ruminococcus的相对丰度显著上升,而Staphylococcus和Adlercreutzia的相对丰度则显著下调。LPC-F干预后,图8-a中的8个属的相对丰度都有不同程度的上升或下降,从而导致其肠道菌群代谢产物SCFAs的水平发生变化,尤其是乙酸和丙酸的含量发生变化。基于此,建立便秘是否缓解的指标(粪便含水量、小肠推进率、排首粒黑便时间)与炎症因子、便秘相关胃肠调节肽(5-HT,SP)、SCFAs、c-kit之间相关性分析,同时建立菌群与SCFAs之间的相关性分析,结果分别如图8-b,图8-c所示,乙酸、丙酸和丁酸的含量与粪便含水量、小肠推进率呈正相关,而与排首粒黑便时间呈负相关;IL-1β则与粪便含水量、小肠推进率呈负相关,而与排首粒黑便时间呈正相关。5-HT与粪便含水量呈正相关,而与排首粒黑便时间呈负相关。c-kit及SP仅与排首粒黑便时间呈负相关。乙酸与Staphylococcus和Adlercreutzia的相对丰度呈正相关,而与Oscillospira,Anaerostipes,Ruminococcus及Akkermansia的相对丰度呈负相关。丙酸与Staphylococcus和Adlercreutzia的相对丰度呈正相关,而与Anaerostipes,Ruminococcus及Akkermansia的相对丰度呈负相关。这就说明肠道菌群结构的改变,导致代谢产物乙酸、丙酸含量发生变化。而我们前期研究发现乙酸具有缓解便秘的作用[23]。结合本文的结果可知L.paracaseiLPC-F缓解便秘的可能机制是:L.paracaseiLPC-F通过改变肠道菌群的结构,提高肠道内乙酸的水平,乙酸通过刺激肠嗜铬细胞释放5-HT,释放入肠腔的5-HT通过与其受体相结合,促进ICC的增殖,从而最终增强肠蠕动缓解便秘。
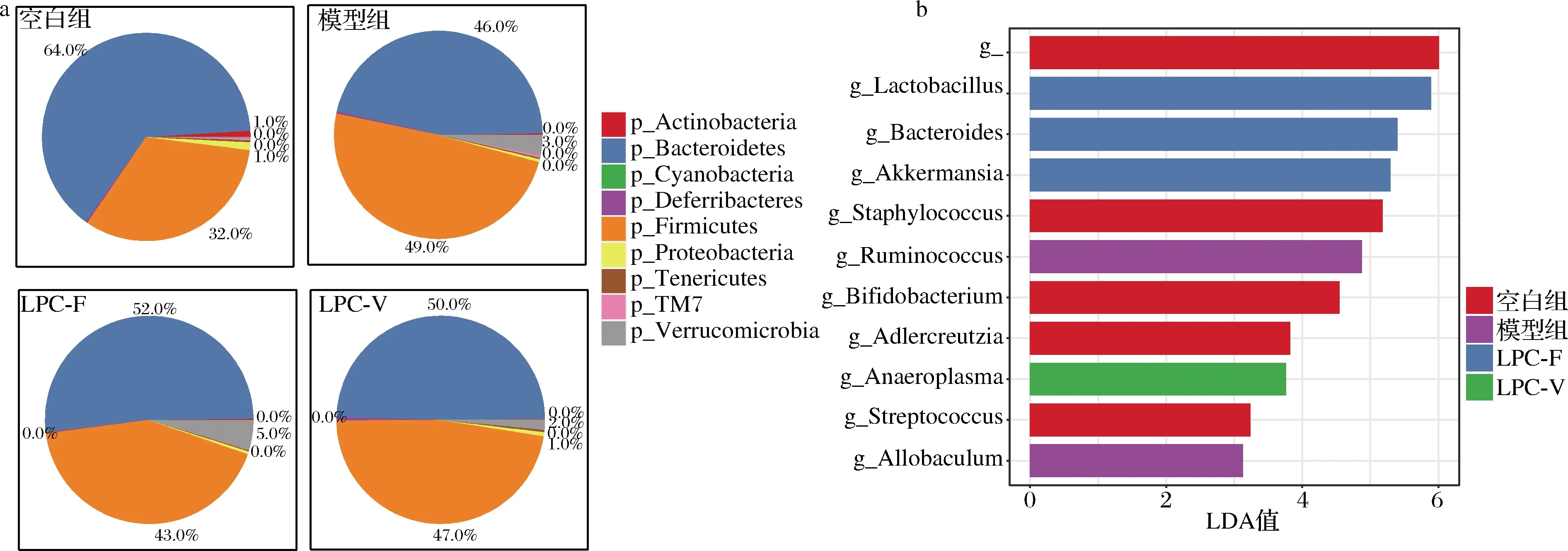
a-门水平上的影响;b-LEfSe分析图7 副干酪乳杆菌对小鼠粪便菌群的影响Fig.7 The effect of L.paracasei on relative abundance of bacteria
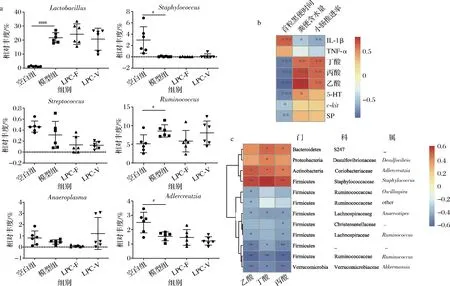
a-副干酪乳杆菌对肠道菌群属水平相对丰度的影响;b-便秘缓解指标与检测指标之间相关性分析;c-SCFA与肠道菌群之间相关性分析图8 干酪乳杆菌对肠道菌群的影响及指标间相关性分析Fig.8 The effect of L.paracasei on relative abundance of intestinal microbiota and the correlation analysis among indicators
3 结论
本研究主要针对2株不同来源的副干酪乳杆菌,研究其缓解便秘的可能机制。通过应用洛哌丁胺构建便秘模型,及对便秘相关指标的检测,发现来源于健康成人粪便的L.paracaseiLPC-F具有缓解便秘的作用。通过对便秘相关胃肠调节肽、血清中炎症因子、ICC的数量、肠道菌群及其代谢产物SCFAs等指标进行检测,并进一步建立肠道菌群与便秘检测指标之间的相关性研究,以解析L.paracaseiLPC-F缓解便秘的可能途径。结果表明,L.paracaseiLPC-F可能通过改变肠道菌群的结构,提高肠道内乙酸的水平,乙酸通过刺激肠嗜铬细胞释放5-HT,释放入肠腔的5-HT通过与其受体相结合,促进ICC的增殖,从而最终增强肠蠕动缓解便秘。
猜你喜欢
福建医科大学学报(2022年3期)2022-09-10
食品研究与开发(2022年14期)2022-07-26
食品界(2022年12期)2022-01-01
临床内科杂志(2021年9期)2021-09-17
天津医科大学学报(2021年4期)2021-08-21
乳业科学与技术(2019年5期)2019-12-24
中国油脂(2019年1期)2019-01-23
中国当代医药(2018年12期)2018-06-16
医食参考(2017年12期)2017-04-01
爱你(2016年20期)2016-11-25
