
不同碳氮源配比对脱氮生丝微菌FJNU-R8生长和PQQ合成的影响
2022-02-24 07:57:16丁灵涛刘东方黄建忠
福建农业科技 2022年12期
丁灵涛,刘东方,黄建忠
(福建师范大学生命科学学院/工业微生物发酵技术国家地方联合工程研究中心, 福建 福州 350108)
吡咯喹啉醌(pyrroloquinoline quinone, PQQ)是继烟酰胺和核黄素之后发现的第3种辅酶[1-2],可以在氧化还原反应中参与电子传递[3],具有特殊的生物活性和生理功能。研究发现,PQQ催化氧化还原反应能力极强[4],可以清除自由基[5-7],修复氧化损伤的神经细胞[8],刺激神经因子增长[9],促进线粒体合成[10],提高生物体生长和代谢水平[11]。匡炜等[12]学者研究发现利用PQQ对水稻种子进行浸种处理后直播,并在始穗期叶面喷洒PQQ,可显著提高结实率;李震等[13]研究发现灌根和施用一定量的PQQ,可以显著改良土壤理化性质;焦子伟等[14]通过试验证明PQQ处理可以促进玉米生长。因此,PQQ在医疗、农业和化妆品等领域有极为广泛的应用前景。
目前,合成PQQ的方法主要有化学合成[15]和生物合成[16]两种方法。其中,化学合成法存在工艺路线复杂、生产效率较低、副产物较多、下游纯化成本较高等问题。微生物发酵法生产PQQ则有成本低、反应条件温和、发酵过程添加物简单、易于分离纯化等诸多优势,成为工业化生产的优选策略。研究表明,用于合成PQQ的菌株多为革兰氏阴性菌[17-19],尤其以甲基营养菌的合成能力最强,虽然现阶段有关PQQ合成的代谢途径已基本阐明[20],但通过代谢工程手段改造原始菌株依然存在不小的困难。因此,目前提高PQQ产量的方法以筛选高产菌株、培养基和培养条件的优化为主,比如柯崇榕[21]通过高浓度甲醇诱变选育出了1株PQQ高产菌株;张静等[22]以HyphomicrobiumdenitrificansCGMCC 1.12893为出发菌,通过ARTP诱变筛选到1株PQQ高产突变菌株,进行发酵培养基优化后将产量提高了3.4倍。
然而,这些高产突变菌株实际发酵周期很长,势必影响实际工业化生产总量,究其原因无非是菌株延滞期和对数生长期时间较长、PQQ生产启动缓慢等主要因素。本研究以脱氮生丝微菌 FJNU-R8(HyphomicrobiumdenitrificansFJNU-R8)为出发菌株,仔细探究了不同的碳氮源对菌株生长和PQQ合成的影响,同时分析了菌株生长和PQQ合成的相互联系,以期为实际发酵策略调整提供理论上的参考。
1 材料与方法
1.1 试验材料
1.1.1菌株 本试验所选用的菌株为脱氮生丝微菌FJNU-R8,由本课题组从污水中筛选并诱变选育后获得,保藏于工业微生物教育部研究中心。
1.1.2培养基 种子培养基:4 g·L-1(NH4)2SO4、3 g·L-1KH2PO4、3 g·L-1Na2HPO4、1.6 g·L-1MgSO4·7H2O、25 mg·L-1CaCl2、5 mg·L-1ZnSO4、0.5 mg·L-1FeSO4、5.0 g·L-1CH3OH、0.4 mL·L-1微量元素和1 mL·L-1维生素;发酵培养基:2 g·L-1(NH4)2SO4、3 g·L-1KH2PO4、3 g·L-1Na2HPO4、1.6 g·L-1MgSO4·7H2O、25 mg·L-1CaCl2、5 mg·L-1ZnSO4、0.5 mg·L-1FeSO4、20.0 g·L-1CH3OH、0.4 mL·L-1微量元素和1 mL·L-1维生素;微量元素:15 mg·L-1NaCl、5 mg·L-1CuSO4·5H2O、5 mg·L-1MnSO4·4H2O、0.3 mg·L-1KI、0.1 g·L-1COCl2·6H2O、3 g·L-1H3BO3;维生素配方:2 g·L-1核黄素、4 g·L-1盐酸吡哆素、4 mg·L-1盐酸硫胺素、2 g·L-1叶酸、400 mg·L-1烟酸、20 mg·L-1对氨基苯甲酸、40 mg·L-1泛酸钙、2 mg·L-1生物素、20 mg·L-1肌醇。所用化学试剂均为分析纯。
1.1.3仪器与设备 双层恒温摇床ZWB-3212B购自上海智城分析仪器制造有限公司;高效液相色谱仪e2695购自沃特世科技(上海)有限公司;紫外可见分光光度计UV-1800购自日本岛津公司;贝克曼J-26冷冻离心机购自美国贝克曼有限公司;科瑞峰超纯水系统SP-D1-20L购自四川德立世科技有限公司。
1.2 试验方法
1.2.1摇瓶培养条件 从-80℃冰箱中取出保种管,在平板上稀释涂布,培养7~8 d。挑取单菌落在平板上培养3~4 d,全部洗下并接种于100 mL/500 mL的液体种子培养基中,于220 r·min-1、30℃条件下培养至对数期后,按3.5%(V/V)接种量转接至挡板摇瓶中,振荡培养[23]。
1.2.2不同碳氮源摇瓶培养基成分 探究单一碳源和氮源对菌株OD650和PQQ产量的影响。(1)碳源包括:甲醇、乙醇、甘油、葡萄糖,以碳原子浓度0.625 mol·L-1(合甲醇浓度20 g·L-1)为标准设计添加量;(2)氮源包括:硫酸铵、硝酸铵、氨水、蛋白胨,以氮原子浓度0.030 mol·L-1(合硫酸铵浓度2 g·L-1)为标准设计添加量。
1.2.3不同复合碳源和氮源配比设计 设计不同配比复合碳源和氮源,(1)复合碳源∶甲醇和甘油配比为20∶80、40∶60、60∶40、80∶20;(2)复合氮源∶硫酸铵和蛋白胨配比为20∶80、40∶60,60∶40,80∶20。
1.2.4复合碳源和氮源不同添加量的设计 (1)基于甲醇∶甘油(20∶80)的复合碳源,设计该配比下总碳源不同的添加量(5、10、20、30、40 g·L-1;等比换算甲醇浓度);(2)基于蛋白胨∶硫酸铵(40∶60)的复合氮源,设计该配比下总氮源不同的添加量(1、2、3、4、5 g·L-1;等比换算硫酸铵浓度)。
1.3 测定方法
取样时间点:48 h前每隔6 h取一次样,48 h后每隔24 h取1次样;每次取样均测OD650,36 h后的样品采用高效液相色谱仪测PQQ的含量。(1)生物量测定方法:使用紫外可见光分光光度计,在波长650 nm下测定摇瓶菌液吸光光度值OD650,控制OD650范围在0.2~0.8以内。(2)PQQ含量测定方法:采用高效液相色谱仪对PQQ含量进行检测[24]。将发酵液10 000 r·min-1、离心3 min,取上清液用0.22 μm的水系膜过滤。HPLC条件为色谱柱UItimate XB C18 4.6 mm×250 mm(5 μm)。检测波长330 nm、柱温40℃、流动相为V(水+1‰ TFA)∶V(乙腈+1‰ TFA)=85∶15、流速为0.5 mL·min-1。
1.4 数据处理
对试验数据采用Excel软件进行数据处理和图表绘制。
2 结果与分析
2.1 单一碳源和单一氮源对菌株OD650和PQQ产量的影响
2.1.1单一碳源对菌株OD650和PQQ产量的影响 由图1、2可知,以乙醇或葡萄糖为单一碳源时,菌株几乎不会生长;以甲醇为单一碳源时,菌株在40 h生长至稳定期并保持4左右的OD650,随后大量合成PQQ,在118 h时PQQ产量为43 mg·L-1;以甘油为单一碳源时,菌株在长达50 h延滞期后开始生长,并在118 h OD650达到9.8,但是不生产PQQ。说明甘油的利用需要菌株更长的准备时间,然后优先利用碳源于细胞碳骨架的构建以增加OD650。综合甘油为碳源时菌株的OD650最高和甲醇为碳源时菌株PQQ产量最高,设计以甲醇和甘油复配的复合碳源。

图1 单一碳源对菌株OD650的影响Fig.1 Effect of the sole carbon source on the OD650 of strain

图2 单一碳源对菌株PQQ产量的影响Fig.2 Effect of the sole carbon source on the PQQ yield of strain
2.1.2单一氮源对菌株OD650和PQQ产量的影响 由图3、4可知,以氨水为单一氮源时,菌株几乎不会生长;以硫酸铵或硝酸铵为单一氮源时,菌株在50 h生长至稳定期并保持OD650,随后在118 h时,PQQ产量分别为29.8、17.0 mg·L-1;以蛋白胨为单一氮源时,菌株前期生长速度更快并在70 h生长至OD650(7.3)的稳定期,但是几乎没有PQQ的合成。因此,设计以硫酸铵和蛋白胨混合的复合氮源,以期提高前期菌株的生长速度,同时兼顾PQQ的产量。
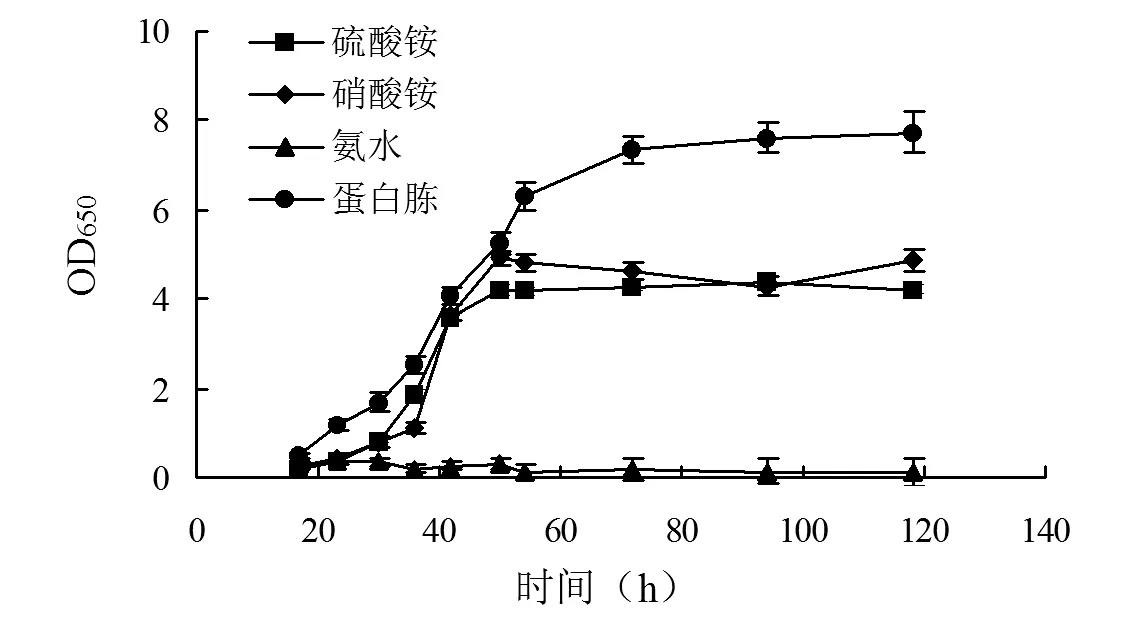
图3 单一氮源对菌株OD650的影响Fig.3 Effect of the sole nitrogen source on the OD650 of strain

图4 单一氮源对菌株PQQ产量的影响Fig.4 Effect of the sole nitrogen source on the PQQ yield of strain
2.2 不同配比复合碳源和复合氮源对菌株OD650和PQQ产量的影响
2.2.1不同配比复合碳源对菌株OD650值的影响 由图5、6可知,PQQ合成菌株在不同配比的复合碳源培养基中的前期生长速度均高于以甲醇或甘油为单一碳源的培养基,且菌株的最终OD650高于以甲醇为单一碳源的培养基,说明甘油的添加确实可以提高OD650,甘油的比例越高,最终OD650也越高。但只有以甲醇为单一碳源时,可以在120 h检测到PQQ产量为53 mg·L-1;在其他复合碳源的培养基中,仅有甲醇∶甘油(20∶80)的复合碳源检测出5 mg·L-1的PQQ产量。上述结果表明,复合碳源可以有效提高OD650,但是干扰了PQQ合成;而以甲醇为单一碳源时,OD650最低,但是PQQ产量最高。为了探讨是否因为甘油的加入导致菌株优先利用碳源用于自身碳骨架的构建(更高的OD650),造成PQQ合成途径碳源供给不足,后续试验以甲醇∶甘油为20∶80的比例出发探讨复合碳源不同添加量的影响。

图5 不同配比复合碳源对OD650的影响Fig.5 Effects of different ratios of composite carbon sources on OD650

图6 不同配比复合碳源对PQQ产量的影响Fig.6 Effects of different ratios of composite carbon sources on the PQQ yield
2.2.2不同配比复合氮源对菌株OD650和PQQ产量的影响 由图7、8可知,PQQ合成菌株在不同复合氮源培养基中的前期生长速度均高于以硫酸铵或蛋白胨为单一氮源时的生长速度。其中蛋白胨∶硫酸铵为60∶40或80∶20时,生产菌株在120 h时的OD650分别为8.6和8.5,远高于同时期以硫酸铵为单一氮源时的OD650(4.3)。从PQQ的合成能力分析,120 h时蛋白胨∶硫酸铵为40∶60的PQQ产量为65 mg·L-1,而同时期下以硫酸铵为单一氮源的PQQ产量仅为42 mg·L-1,其余氮源培养基的PQQ产量均低于上述两个条件。从生长曲线和PQQ合成产量出发,探讨蛋白胨∶硫酸铵为40∶60配比时不同添加量的影响。
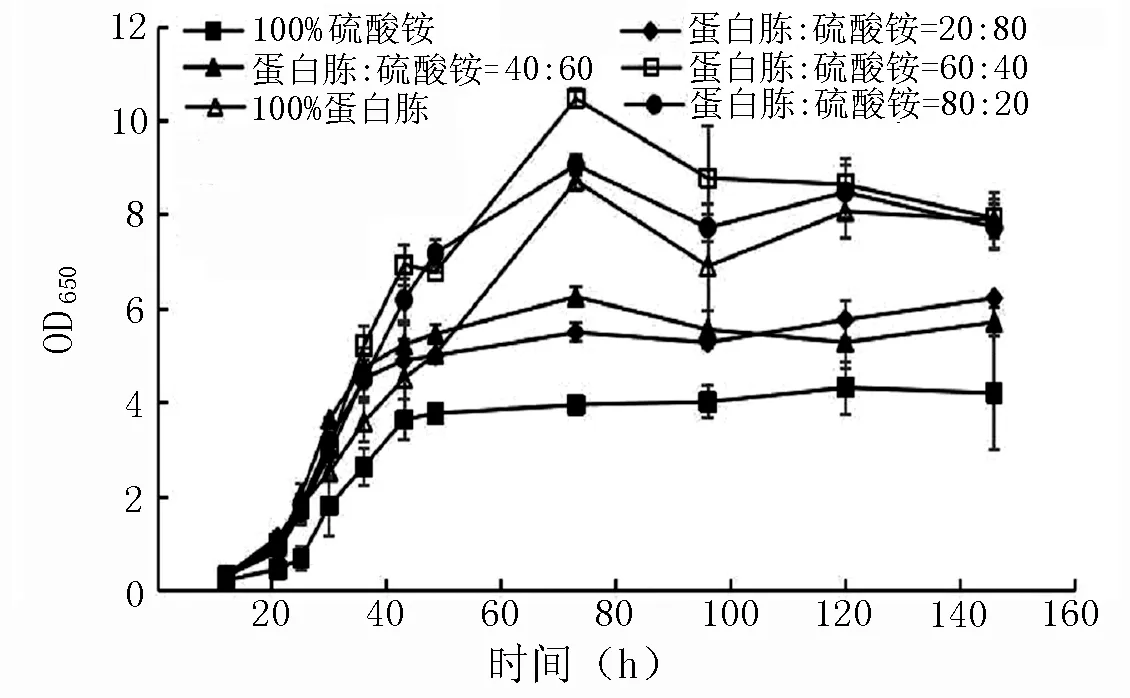
图7 不同配比复合氮源对OD650的影响Fig.7 Effects of different ratios of composite nitrogen sources on OD650

图8 不同配比复合氮源对PQQ产量的影响Fig.8 Effects of different ratios of composite nitrogen sources on the PQQ yield
2.3 复合碳源和复合氮源不同添加量对菌株OD650和PQQ的影响
2.3.1复合碳源(甲醇∶甘油=20∶80)不同添加量对菌株OD650和PQQ产量的影响 由图9、10可知,以甲醇∶甘油为20∶80的复合碳源时,在30 h时,两个低浓度添加量(5、10 g·L-1)生长明显延滞,156 h时OD650仅为1.8和3.6,对应的PQQ的产量为17和15 mg·L-1。高浓度添加量(20、30、40 g·L-1)条件下,培养40 h左右生长减缓,大概经过30 h后继续生长,添加浓度越高,最终的OD650也越高。浓度40 g·L-1条件下156 h时OD650可达10.8,对应的PQQ产量仅为8 mg·L-1。上述结果表明,添加甘油的复合碳源有利于PQQ生产菌株OD650的增加,且复合碳源的添加量越高, OD650也越高;复合碳源添加量的变化并不能有效提高PQQ产量,说明不是由于碳源不足导致PQQ合成的降低,而是甘油的进入影响了PQQ合成途径,而且甘油添加浓度越高,对PQQ合成的干扰越大。此外,研究发现OD650基本稳定的菌株PQQ产量高,推测菌株中后期稳定的OD650更有利于PQQ的合成。

图9 复合碳源不同添加量对OD650的影响Fig.9 Effect of different amount of composite carbon sources on OD650

图10 复合碳源不同添加量对PQQ产量的影响Fig.10 Effect of different amount of composite carbon sources on the PQQ yield

图11 复合氮源不同添加量对OD650的影响Fig.11 Effect of different amount of composite nitrogen sources on OD650
2.3.2复合氮源(蛋白胨∶硫酸铵=40∶60)不同添加量对菌株OD650和PQQ产量的影响 由图11、12可知,在以蛋白胨∶硫酸铵为40∶60的复合氮源条件下,复合氮源不同添加量似乎对菌株的生长和PQQ的合成影响很小。浓度1 g·L-1条件下,156 h的OD650为7.3,PQQ产量为11 mg·L-1;其余条件下156 h的OD650为5.5左右,PQQ产量为45 mg·L-1左右。由此推测,在碳源一定的情况下,只要维持基本氮源量,复合氮源的添加量不是影响菌株生长和PQQ合成的关键因素。此外,研究发现OD650波动明显的菌株PQQ产量最低,推测菌株中后期稳定的OD650更有利于PQQ的合成。

图12 复合氮源不同添加量对PQQ产量的影响Fig.12 Effect of different amount of composite nitrogen sources on the PQQ yield
3 讨论与结论
PQQ作为氧化还原酶的辅因子,广泛应用于医药、化妆品和农业等领域。目前,PQQ的合成以甲基营养菌发酵为主,其产量的提高多依靠诱变筛选高性能菌株,缺乏基因工程的介入从而改造菌株实现PQQ产量的大幅提高,究其原因在于这类菌株体内代谢网络十分复杂,很多相关的代谢途径至今尚不明确,此外对其限制修饰系统[25]也鲜有研究,从而导致基因操作难度大幅度上升。虽然现在已经有很多条件优化[16]来进一步提高PQQ的产量,却难以实现大幅度的提升。
本研究从最基础的碳氮源出发,探讨了不同配比碳源和氮源对生丝微菌FJNU-R8生长和PQQ合成的影响。试验结果表明,以甲醇为单一碳源时PQQ产量最高,但是最终OD650只有4.4,这个结果印证了PQQ是甲醇脱氢酶的辅酶,即在利用甲醇时需要大量的甲醇脱氢酶将甲醇转化为甲醛,进而进入代谢途径用于菌株物质和能量的合成,因此菌株需要在较早时期积累PQQ。添加甘油后的复合碳源可以显著提高最终OD650,但是检测出PQQ的产量远低于以甲醇为单一碳源时的PQQ产量,这可能是由于甘油的添加影响了感知甲醇的双组分系统[26-27]的信号传递,从而下调了甲醇脱氢酶甚至PQQ合成基因簇的表达。以硫酸铵为单一氮源时PQQ产量最高,但是最终OD650仅为4.2;添加蛋白胨的复合氮源可以提高菌株生长速度和最终OD650,当蛋白胨∶硫酸铵为40∶60时还可以提高PQQ产量。PQQ的合成与生长的关系为部分偶联型,在中后期保持稳定的OD650更有利于积累PQQ。
本研究发现甘油的添加可能干扰PQQ的合成,但并非完全阻遏PQQ生产,因此可以只考虑复合碳源提高总OD650,然后选用流加甲醇为单一碳源的方式积累PQQ。值得思考的是,混有甘油的复合碳源利用速度较慢,如何提高甘油的快速利用达到前期快速积累OD650仍需要进一步探究。此外,复合碳源和复合氮源添加量的变化差异很大,如何调整合适的碳氮比使菌株前期专注于生长,中后期专注于PQQ合成值得进一步讨论。在未来试验中,可以进一步筛选合适的复合碳源和氮源并探讨相应的添加方式以期获得更好的发酵策略,并在发酵中后期维持发酵总OD650从而实现PQQ产量的提高。
猜你喜欢
中国化肥信息(2022年6期)2023-01-06 05:36:01
中国化肥信息(2020年3期)2021-01-07 02:12:58
中国化肥信息(2018年8期)2018-10-08 03:17:10
能源(2017年7期)2018-01-19 05:05:04
中学科技(2017年11期)2017-12-26 10:14:01
中国民族医药杂志(2016年4期)2016-05-09 07:41:11
中国酿造(2016年12期)2016-03-01 03:08:11
中国酿造(2014年9期)2014-03-11 20:21:03
食品工业科技(2014年9期)2014-03-11 18:15:28
西部中医药(2014年6期)2014-03-11 16:07:47
