
维生素C对虾类生长和健康调节作用研究进展
2022-02-20 02:46:30谭青松孔凡双朱文欢
动物营养学报 2022年1期
谭青松 孔凡双 朱文欢
(1.华中农业大学水产学院,武汉 430070;2.水产养殖国家级实验教学示范中心,武汉 430070;3.武汉市水产技术推广指导中心,武汉 430012)
虾类一直是国内外重要的水产养殖对象。但当前养虾业中,很多虾类还在使用活饵料,还有些虾的营养研究匮乏,其饲料配制参常考其他虾的营养需求[1]。针对特定养殖虾品种的营养需求来开发高效配合饲料是未来的研究焦点[2]。随着高密度养殖的持续发展,一些虾类在养殖过程出现生长变慢、免疫能力变低等问题[3]。此外,高密度养殖还造成环境污染,使得全球虾类养殖易受到较大规模的流行病的影响,包括早死症、黑死病等[4]。传统的虾类疾病防治主要依靠化学药物和抗生素,其过度使用易造成许多副作用,如药物残留、病菌耐药性、自身免疫抑制、微生态平衡破坏等,严重影响虾类养殖的可持续发展[5]。
维生素C是一种强效抗氧化剂和免疫调节剂,可用作饲料添加剂来改善养殖动物健康,减少抗生素的使用[6]。鱼虾类需要摄入维生素C才能保持良好的健康状况,其生长性能也与饲料中维生素C含量密切相关[7]。因此,利用维生素C来调控虾类的生长和健康,对养虾业健康发展意义重大。本文概述维生素C的结构和基本性质及其对虾类生长、免疫和抗氧化的影响及相关作用机制,归纳维生素C在虾类饲料中的适宜添加量和高效价剂型,并指出未来维生素C在虾类中的研究方向,为虾类的健康养殖和饲料生产提供参考。
1 维生素C的结构、特性和生物合成
1.1 维生素C的结构与特性
维生素C,又称L-抗坏血酸,简称为VC、抗坏血酸,其化学名称为2-氧代-L-苏式-己酮-1,4-内酯-2,3-烯二醇,相对分子质量为176.12,分子式为C6H8O6,其结构式如图1所示。
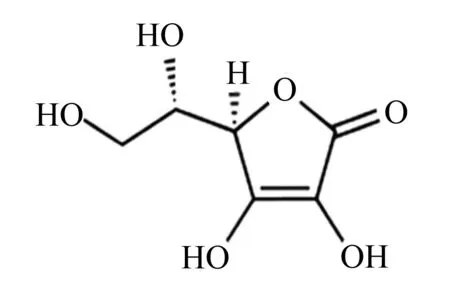
图1 维生素C的分子结构Fig.1 Molecular structure of vitamin C[7]
维生素C具有很强的还原性,其C2和C3之间的基团(在双键上有2个OH基团)为烯二醇,很容易产生2个质子和电子,转化为脱氢抗坏血酸(dehydroascorbate,DHA,维生素C的氧化形式)的二酮基。该基团的存在决定了维生素C的强还原性和酸性[8]。由于L-抗坏血酸具备还原性,可以在生物体内转化为L-DHA。这2种维生素C形式(被还原和被氧化)具有相同维生素活性。由于L-抗坏血酸的酸性,在C3处的羟基质子易失去,形成抗坏血酸钙或铁的盐[8]。在哺乳动物中的研究表明,抗坏血酸盐是单电子反应中最丰富的水溶性化合物,其在许多金属依赖的加氧酶催化的反应中扮演辅助因子的作用。例如,对于Fe2+/α-酮戊二酸依赖性双加氧酶来说,L-抗坏血酸可将不活跃的含Fe3+的酶重新转化为有活性的含Fe2+的形式[9]。
1.2 维生素C的合成及其对虾类的必需性
植物和哺乳动物体内可合成维生素C,其合成过程[10]如图2所示。一般由D-葡萄糖或D-半乳糖作为合成底物,其中葡萄糖-6-二磷酸被4个连续的中间体转化为L-古洛糖酸-γ-内酯,后经L-古洛糖内酯氧化酶(L-gulonolactone oxidase)氧化为2-酮基-L-葡萄糖酸内酯,进而生成L-抗坏血酸。

图2 维生素C的合成途径Fig.2 Synthesis pathway of vitamin C
一般认为,虾体内因缺乏L-古洛糖内酯氧化酶而不能自身合成维生素C,必须从饲料中获得足量的维生素C以满足需求[11]。维生素C缺乏会导致虾类的饲料转化率低、生长不良、抗应激能力降低、死亡率高、伤口愈合受损和黑化损伤[12-14]。徐志昌等[15]报道了饲料中维生素C可以改善中国对虾(Penaeuschinensis)的蜕壳反应,但高剂量维生素C下的蜕壳频率及生长反而受到抑制。在加州对虾(Penaeuscaliforniensis)和南美蓝对虾(Penaeusstylirostris)上也有类似结果的报道[16]。在马氏沼虾(Macrobrachiummalcolmsonii)的研究中还发现,补充维生素C改善了虾的消化酶分泌[11]。
2 虾类对维生素C的营养需求
确定维生素C最适需求量的评判标准包括:特定生长率、增重率、存活率、饲料系数、组织累积量等。同时,虾类对维生素C的需求量受其种类及发育阶段、养殖水温、饲料其他成分以及维生素C剂型等因素影响[7,17]。
2.1 虾类对维生素C需求量的研究
表1列出了国内外几种常见养殖虾品种对维生素C的需求量。由表可见,不同虾类的维生素C需求量存在较大差异,这也提示不同种类虾的维生素C需求量不能相互参照,需要精确试验来确定。评价指标、生长阶段和剂型等也是影响维生素C需求量的重要因素。先前对凡纳滨对虾(Litopenaeusvannamei)的研究表明,饲料维生素C添加量在150 mg/kg时,促生长效果最好[18]。但近年来,Chen等[19]通过对存活率进行折线回归分析得到的凡纳滨对虾对饲料维生素C的需求量更高。最近一项克氏原螯虾(Procambarusclarkii)对维生素C的营养需要研究表明,基于特定生长率回归分析得到的最佳饲料维生素C含量因维生素C剂型不同而存在差异[20]。Celada等[21]根据淡水通讯螯虾(Pacifastacusleniusculus)生长率和存活率作为评判指标,发现幼虾由于生理代谢旺盛、生长迅速,相比成年虾对维生素C的需求量要更高。
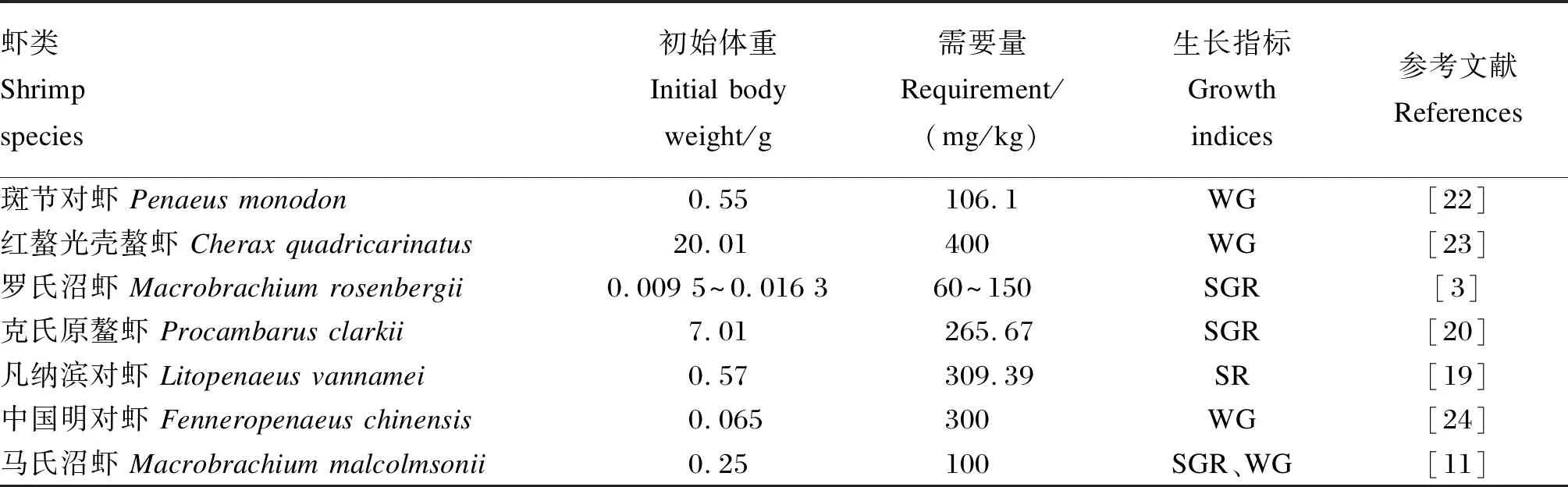
表1 虾类基于生长指标对维生素C的需求量Table 1 Vitamin C requirement for shrimp species based on growth indices
2.2 维生素C剂型在虾类饲料中的应用
维生素C有较强的还原性和结构不稳定性,通常要避光避热保存[8],在加工和储存过程中也很容易降解[21]。通过在结晶维生素C表面进行包被处理,或对维生素C碳环上的活性羟基基团进行修饰,可获取稳定性更高的保护型维生素C,提升其生物效价[25]。早期常采用乙基纤维素、脂质包膜来稳定维生素C,后来又发现可在维生素C结构中引入硫酸基团(抗坏血酸-2-硫酸酯)来改善维生素C的稳定性[26]。近年来报道,维生素C磷酸衍生物的稳定性和生物利用率更高[7]。表2列出了几种常见剂型维生素C的抗坏血酸含量及其稳定性。由于不同抗坏血酸剂型在加工、运输和储存过程中的活性损失,饲料配方设计时,需要综合其剂型、需要量和其他营养物质的交互作用等情况来供给维生素C,以确保充足活性成分的维生素C[26]。
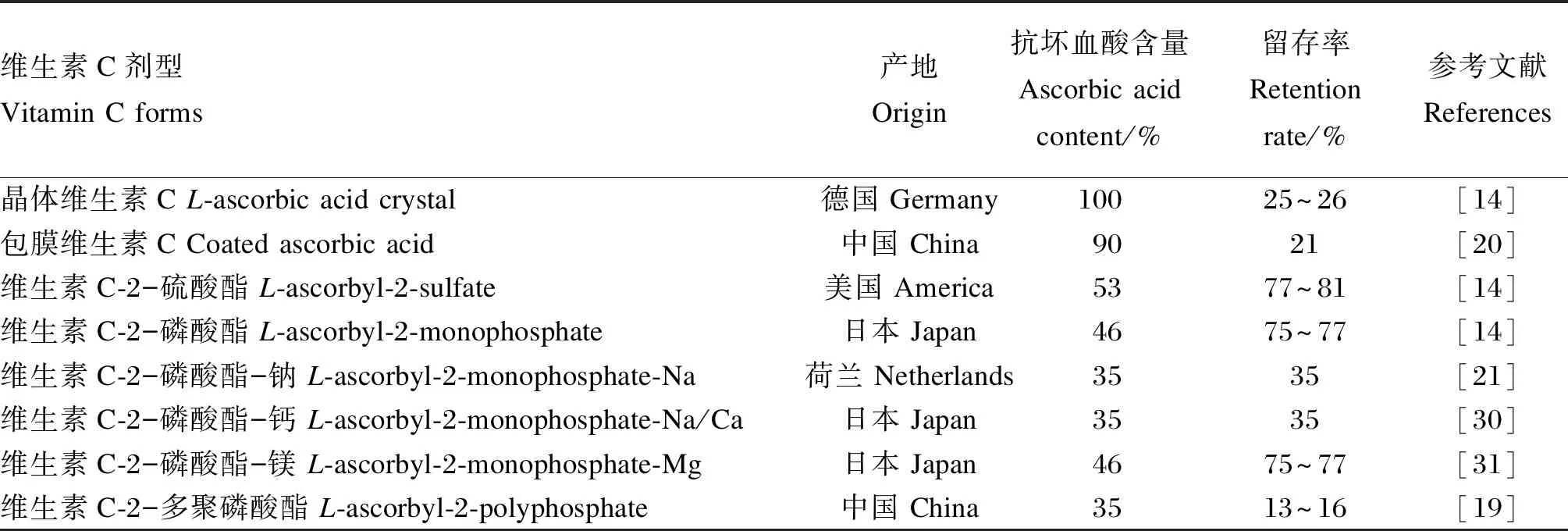
表2 常见维生素C剂型的抗坏血酸含量及其稳定性Table 2 Ascorbic acid content in common vitamin C forms and its stability
维生素C剂型在各种虾类中的生物学效价并不完全一致。马甡等[27]用含包膜维生素C和维生素C磷酸酯镁的2种饲料投喂中国对虾幼虾后发现,包膜维生素C组的存活率比维生素C磷酸酯镁组的要高;在两者提供相同的维生素C效价时,前者的需求量相对较低。李爱杰[28]用几种不同剂型维生素C进行中国对虾养殖试验时发现,稳定型维生素C比结晶维生素C的促生长效果好;在稳定型维生素C中,以维生素C磷酸酯镁和维生素C硫酸酯的平均增重率较高,而维生素C多聚磷酸酯的效果较差,但其平均存活率却最高。孔凡双[29]分别将含包膜维生素C和维生素C磷酸酯的试验饲料饲喂克氏原螯虾,结果表明包膜维生素C可以增强虾血清中更全面的免疫酶活性,并且基于特定生长率对2种维生素C剂型的最佳需求量比较得出,克氏原螯虾对包膜维生素C的需求量更低,由此可知包膜维生素C更容易被克氏原螯虾吸收和生物利用,其生物学效价更高。但值得注意的是,维生素C磷酸酯在此试验饲料中的活性成分抗坏血酸的留存率比包膜维生素C高很多。这些研究进一步说明,维生素C的需求量会受到不同评价指标以及不同剂型维生素C的效价等因素的综合影响。值得针对养殖虾类品种进一步研究各维生素C剂型的效价。
3 维生素C对虾类生长发育的调节作用
3.1 维生素C对虾类的促生长作用
维生素C对虾类的促生长作用已经得到广泛证实。早期在斑节对虾(Penaeusmonodon)上的研究表明,饲料中添加适量的维生素C可以显著促进斑节对虾的生长和存活,并降低饲料系数[22,32]。宋理平等[24]研究发现,饲料中添加0.03%的维生素C可显著促进中国明对虾(Fenneropenaeuschinensis)的增重率。近几年对罗氏沼虾(Macrobrachiumrosenbergii)[11]和克氏原螯虾[20,33]的研究也表明,维生素C提高了饲料利用率,改善了虾的生长。Zhou等[18]还较系统地研究了维生素C对凡纳滨对虾不同成长阶段的影响,发现维生素C可大幅度提高幼虾的生长率,而对成虾的促生长效果不明显。
虾类生长过程中需不断进行蜕壳。当饲料中营养成分缺乏时,虾类蜕壳会受到不良影响,进而限制其生长[28]。宋理平等[24]总结了维生素C促进虾类脱壳的作用机理:维生素C参与甲壳最外层的硬化,该硬化层由一种外壳蛋白和苯醌经交联作用而成,而体内苯醌的形成需要维生素C的参与。徐志昌等[15]确定了促进中国对虾蜕壳周期和生长的最佳饲料维生素C含量为40 mg/kg。但值得注意的是,该研究还表明过量的维生素C会抑制中国对虾的蜕壳和生长。吴东蕾等[23]还从肝胰腺代谢和蜕壳基因层面开展了研究,证实维生素C能促进虾类蜕壳基因的表达,并发现维生素C含量过高时,红螯光壳螯虾的生长大幅度减慢。不过该研究的时间较短,没有探究养殖后期维生素C对虾类蜕壳频率的影响。
3.2 维生素C对虾类繁殖性能的影响
适量的维生素C具有改善虾类繁殖性能的作用。Alava等[34]研究表明,饲料中补充500~1 000 mg/kg维生素C可改善日本对虾(Penaeusjaponicus)的性腺发育,而补充1 500 mg/kg或未补充维生素C均可导致其性腺发育不良。Xu等[35]发现,0.06%饲料维生素C含量可以加快克氏原螯虾的性腺发育,促进性激素的分泌。Ferraz等[36]对凡纳滨对虾繁殖阶段的研究发现,饲料中添加适量维生素C可以提高虾的产卵率、孵化率,而维生素C含量过高时会产生负面作用。邵光明等[33]认为,维生素C改善虾类的繁殖性能可能与降低性腺中的丙二醛(MDA)含量、保护性腺免受氧化损伤相关,尤其是在卵巢中的作用更明显。宋光同等[37]还研究了多种营养素组合对克氏原螯虾繁殖性能的影响,结果发现,维生素C、维生素E间具有协同作用,促进虾体内类固醇激素的合成和分泌,加速虾的性腺和胚胎发育,但关于具体的适宜添加量组合仍有待进一步研究。
4 维生素C对虾类营养健康的调节作用
4.1 维生素C对虾类体内营养成分的调节作用
通过在饲料中补充维生素C,马氏沼虾体的粗蛋白质含量得到了改善[11]。近期在克氏原螯虾上的研究也表明了饲料中补充维生素C可增加虾体中粗蛋白质的含量,并阐述了维生素C在机体蛋白质代谢中的重要作用[20]。López等[38]报道了1 500 mg/kg的维生素C可以增加凡纳滨对虾体内的糖原含量以作为机体能量来源,从而增加膳食蛋白质吸收后在体内沉积。在克氏原螯虾中还发现,以含维生素C饲料喂养的虾体粗脂肪含量显著高于缺乏维生素C饲料投喂的虾体[20],这表明维生素C还具有较强的促进机体脂肪合成的能力[39],在罗氏沼虾上也报道了维生素C具有促进体脂增加的效果[11]。肝体比(hepatosomatic index,HSI)是生物体健康和能量储备的指标[20]。Cavalli等[40]报道了饲料补充维生素C可以增强罗氏沼虾等甲壳类动物肝胰腺中蛋黄前体的合成,进而提高虾的HSI。在日本囊对虾(Marsupenaeusjaponicus)[41]和克氏原鳌虾[20]的研究中都得到了维生素C改善虾的HSI这一结果。
4.2 维生素C对虾类非特异性免疫的调节作用
在绝大多数哺乳动物的免疫应答中,白细胞(特别是中性粒细胞和单核细胞来源的巨噬细胞)是伤口愈合的主要参与者。维生素C可影响中性粒细胞几个重要的功能:对炎症介质反应的迁移(趋化)、吞噬和杀死微生物以及巨噬细胞凋亡和清除[42]。简言之,维生素C在吞噬细胞中积累,并增强其细胞的趋化性、吞噬作用和活性氧(reactive oxygen species,ROS)的产生,从而杀死病原微生物[42]。其中,促炎症转录因子核因子-κB(nuclear factor-kappa B,NF-κB)在炎症反应的信号通路中发挥核心的作用[42]。Ellulu[43]报道了维生素C在人体的抗炎作用机制归因于其调节NF-κB-DNA结合活性的能力。因此,维生素C可通过直接增强免疫细胞的功能或调节敏感的细胞信号通路来调节免疫反应。
虾类等甲壳类动物易遭受其生活水环境中的不同寄生虫和病原体的侵扰。然而,虾类缺乏脊椎动物中典型的抗体和基于T/B细胞的后天免疫[44],所以虾类完全依赖于其先天免疫系统。血液成分是重要的生理、病理和毒理学指标,其各种变化被广泛用于评估水生动物的健康和营养状况[45]。酚氧化酶(PO)、溶菌酶(LZM)、酸性磷酸酶(ACP)、呼吸爆发(RB)活性以及血细胞总数(THC)等血液学指标常用来评价虾类的免疫能力。饲料中添加0.06%~0.12%的维生素C增强了克氏原螯虾血清PO、LZM和ACP活性,但在提高添加量至0.15%并不会使上述免疫酶的活性持续升高[20]。在斑节对虾[31]和克氏原螯虾[20]中,与缺乏维生素C饲料相比,补充维生素C的饲料显著提高了虾的RB活性和THC。这表明,维生素C不仅会影响虾类的生长和饲料利用,还能调节虾类的免疫能力[7]。但值得注意的是,马氏沼虾[11]和凡纳滨对虾[18]的THC在高水平维生素C下并没有明显改善。这可能是由于试验虾的消化代谢、研究方法和取样时段存在较大差异。目前还缺乏维生素C调控虾类免疫功能的机制研究,这可能是今后的研究重点。
4.3 维生素C对虾类抗氧化的调节作用
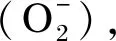
肝胰腺是甲壳类动物最重要的组织,在排毒、营养代谢和免疫防御中起着重要作用[47]。一般认为,甲壳类肝胰腺中的超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、谷胱甘肽过氧化物酶(GPx)和还原型谷胱甘肽(GSH)是对抗自由基和其他氧化损伤的关键抗氧化酶防御体系[20]。Asaikkutti等[11]报道了25~100 mg/kg的维生素C可以提高马氏沼虾的SOD和CAT活性,进而改善抗氧化能力。在克氏原螯虾中发现,与缺乏维生素C组相比,补充维生素C可以明显增强CAT、GPx活性和GSH含量,并以此改善生长性能[20]。此外,MDA作为脂质过氧化过程的最终产物,可以引起细胞毒性应激,是评估氧化应激水平的生物标志物[48]。饲料中添加维生素C降低了凡纳滨对虾[19]和马氏沼虾[11]的MDA含量,这进一步证实了维生素C对虾类的抗氧化作用。此外,维生素C具备抗黑变作用,能改善虾肉品质。郝云彬等[49]通过检测中华管鞭虾(Solenoceracrassicornis)中的多酚氧化酶(PPO)来探究维生素C等组合试剂对该指标的抑制作用,由此得出最佳维生素C含量在0.010~0.018 g/L的复合抑制剂既可以通过与PPO活性中心络合,直接对PPO发挥抑制作用,还通过还原中间产物醛类,抑制虾的黑色素生成。周宇芳等[50]通过中华管鞭虾PPO活性探究维生素C对其作用机理,也得出了相似的结论。庄秋丽等[51]进一步探究了维生素C在复合保鲜剂中的最适剂量,发现1.5 g/L的维生素C搭配0.5 g/L溶菌酶和7.0 g/L柠檬酸能够抑制微生物的生存,并减少组织氧化,改善虾肉品质。这些结果为虾类养殖中的抗氧化能力改善以及肉品质提升提供了参考。
4.4 维生素C对虾类抗应激和抗病毒的调节作用
目前,维生素C也作为具有抗应激作用的营养素被加入到饲料中,用于缓解高密集养殖[52]和冷休克等造成的应激[53]。补充维生素C可抑制受应激鱼类的皮质醇含量,并调节热休克蛋白70(HSP70)的表达水平[7]。关于维生素C对虾类抗应激的研究还很有限。Wang等[46]研究发现,在饲料中补充维生素C提高了凡纳滨对虾的抗氧化与抗氨氮应激的能力。Niu等[54]研究发现,饲料中补充维生素C可以缓解凡纳滨对虾在低溶解氧环境的应激。近年来,维生素C在其他水产动物上的抗应激作用得到了较多研究[55-56],这些研究都证实了维生素C可以降低应激反应,为虾类健康养殖提供了理论指导。但是,关于这方面的基础研究相对滞后,针对其机理方面探究较少。
白斑综合征(white spot syndrome,WSS)作为一种重要的虾类病毒性疾病,广泛危害着世界各地的商业化虾类养殖。所有的虾类基本都能够作为白斑综合征病毒(white spot syndrome virus,WSSV)的宿主被感染[57]。目前针对虾类对WSSV等病毒的抗性机制尚不完全清楚。Ocampo等[58]研究表明,凡纳滨对虾等虾类在病毒感染中得以幸存下来,个体免疫反应起着至关重要的作用:病毒在特定组织中的浸润、吞噬作用和包封;淋巴器官是虾循环系统必不可少的部分,可以用作病毒颗粒过滤器。WSSV感染日本对虾后,最典型的免疫应答是开放循环系统中的血淋巴细胞数量急剧下降,淋巴细胞的凝结功能降低[59]。维生素C作为一种免疫调节剂,可以通过增强免疫反应来提高虾类的抗病能力。近期针对克氏原螯虾饲喂5种不同维生素C含量(0.03%、0.06%、0.09%、0.12%、0.15%)饲料后的攻毒试验证实了饲料中补充维生素C可以提高虾对WSSV的抗病力[20]。这为克氏原螯虾等虾类养殖中的WSS防治提供了参考。
5 小 结
综上所述,维生素C不仅能够促进虾类的存活与生长发育,改善繁殖性能,还可以提升虾类的非特异性免疫应答、抗应激和抗氧化能力。但是,关于虾类维生素C的营养研究尚存在不足:1)养殖虾类的维生素C营养研究非常有限,仅对少数养殖虾品种开展了研究;2)已有的虾类维生素C需求量的探究缺乏相对统一的试验条件(试验环境、虾种类和维生素C剂型)和评判标准(评价指标和数据分析方法),这使得现有研究结果的推广应用价值受限;3)没有采用多种指标结合来系统评价虾类的维生素C需求量,尤其是早期研究仅考虑生长性能的影响;4)缺乏对维生素C在虾类中生理生化功效的分子机理探究。
针对上述不足,未来关于虾类的维生素C营养研究应开展营养学与免疫学、分子生物学和细胞学的交叉研究,结合多种研究手段来进行全面而系统的虾类维生素C营养研究,并从分子层次深入挖掘其对虾类的作用机制,探究相关基因和蛋白的调控网络。此外,还需要针对不同养殖模式、养殖环境系统开展维生素C的剂型和添加方式研究。针对维生素C在虾类病害防治、抗应激、抗氧化、促蜕壳、促生长、促产卵等方面的作用,还值得探究其与其他添加剂组合使用的效应以开发功能性饲料和饲料添加剂。
猜你喜欢
当代水产(2022年8期)2022-09-20 06:47:02
学苑创造·A版(2021年10期)2021-10-30 10:34:43
农民致富之友(2020年27期)2020-09-12 14:07:33
今日农业(2019年15期)2019-01-03 12:11:33
中成药(2017年3期)2017-05-17 06:08:45
山西大同大学学报(自然科学版)(2016年6期)2016-01-30 08:29:25
分析测试学报(2015年8期)2016-01-13 06:19:33
肝博士(2015年2期)2015-02-27 10:49:49
世界热带农业信息(2014年11期)2015-01-05 18:01:03
应用化工(2014年11期)2014-08-16 15:59:13
