
纳木措水体可培养丝状真菌优势种的时空特征研究
2022-02-19 12:28薛文凯朱攀德吉郭小芳
生态环境学报 2022年12期
薛文凯,朱攀,德吉,郭小芳
西藏大学理学院,西藏 拉萨 850000
青藏高原是世界上海拔最高、面积最大的高原,其高海拔、低氧、复杂的地貌和脆弱的生态环境形成了独特的生态系统(Tang et al.,2017)。青藏高原的湖泊系统由1500多个大小不同的湖泊组成,其面积占中国湖泊总面积的49.5%(Wan et al.,2014)。青藏高原具有山脉众多,太阳辐射强、降水少、蒸发量大等地理和气候特征,从而形成了以咸水湖和盐湖为主的湖泊系统(Zhang et al.,2019)。纳木措作为青藏高原代表性的咸水湖,海拔高达4720 m,水面积超过500 km2,是地球上最高的湖泊之一(Spencer et al.,2014)。水体中的微生物在有机物和污染物的生物降解或转化中起着关键作用,维持着水体的生态平衡。某些微生物的存在与特定类型的污染密切相关,是作为评估水质和气候的重要生物指标(Glasl et al.,2017)。
丝状真菌作为组成水体微生物的重要类群,它们适应力强,广泛分布于各种生境中,即便在高盐度的死海中仍能存活较长时间(Kinsey et al.,2003;Kis-Papo et al.,2003)。目前关于青藏高原水体微生物的研究主要包括细菌和真菌的多样性及影响因子(王艳红等,2021;郑艳艳等;2022);自然生态系统条件下细菌群落的多样性(Wang et al.,2018;Antunes et al.,2022);生态位方面的研究以原生动物为主(安瑞志等,2021;张鹏等,2022)。但用水体丝状真菌的群落演变特征来反映水文环境变化的研究较少。冰川和高山湖泊生态系统是对气候变化和人为影响极为敏感的环境。随着旅游业的发展,人类活动对高原环境的影响日益凸显,已有研究表明近年来青藏高原周围地区的大气污染和黑烟越来越多,导致高原上污染物的沉积增加(Xu et al.,2009;Cong et al.,2013;Spencer et al.,2014)。受全球变暖的影响,降水、冰川和永久冻土的消融增强,导致湖泊储水量增加和水文条件发生改变(Zhang et al.,2020),高原湖泊正面临着污染和气候变化带来的威胁。
优势种是群落中占优势地位的种群,研究优势种及其生态位对理解群落中各种群间在时空上的相互关系以及对资源的竞争程度具有重要意义(Grinnell,1917;Helaouët et al.,2009)。同时低温的湖水环境也有较大几率分离到耐冷的菌株。因此本研究以纳木措春、夏、秋 3个季节的沿岸水体为样本,分离纯化出其中的可培养丝状真菌,探究丝状真菌优势种的时空生态位和季节变化特征以及其群落中物种组成的变化规律,以期为利用微生物物种组成特征作为水质和气候变化的生物指标提供理论基础,为低温环境污水的生物修复提供菌株资源。
1 材料与方法
1.1 水样的采集
在2020年8月以及2021年5月和10月,沿湖区周边选择 20 个样点(91°02′06″—90°15′31″E,30°54′32″—30°30′14″N),各样点相距 15 km 左右(纳木措禁止船只航行;为了在纳木措湖周边均匀设置样点,在 Google地图上按纳木措湖周长与样点数量的比值定位标记各采样点)。用消毒过的水样采集器,采集离湖岸15 m处的水样,各样点采5 L水,盛于2个无菌桶中,用于环境因子的测定和丝状真菌的分离。在样点信息记录中,除 19号样点为观景台,样点存在地表径流外,他样点周边植被稀疏,明显特征。
1.2 水体环境因子的测定
测定纳木措水体的 8种环境因子,水体温度(t)、pH、总溶解固体量(TDS)、电导率(EC)和盐浓度(Salt),以多功能参数仪(Multi-Parameter PCSTestrTM35)进行原位测定,3次重复。铵态氮(NH4+-N)、总氮(TN)以及总磷(TP)的测定,由西藏博源检测公司完成,其中,铵态氮(NH4+-N)测定采用纳氏试剂分光光度法;总氮(TN)采用碱性过硫酸钾消解紫外分光光度法;总磷(TP)采用钼酸铵分光光度法(邢殿楼等,2016)。
1.3 丝状真菌的分离、纯化及鉴定
用孔径0.45 μm、直径50 mm的无菌醋酸纤维素滤膜,抽滤50 mL和100 mL的湖水,每梯度各抽滤3个重复,抽滤后将滤膜贴于固体RBCH、酸性PDA、酸性YM平板培养基上进行20 ℃的恒温培养,培养基配比参照亚萍等(2013)。3—5 d后统计平板上的菌落数,挑取单个菌落接种在RBCH斜面培养基上纯化,继续恒温培养相同时间后观察并记录其形态特征(若有杂菌污染则再次纯化),将纯化好的菌株保存在体积分数为 30%的甘油管中(−20 ℃),用于高原微生物种质资源库的建设。
分别用点植法和插片培养法培养纯化菌种,观察丝状真菌的菌落形态和显微特征,根据真菌的分类鉴定指标对所属类群做初步鉴定(Ainsworth,2008)。分子鉴定由上海生工完成,其中DNA的提取、PCR的扩增采用CTAB法(孙立夫等,2009),引物为 ITS1(5′-TCCGTAGGTGAACCTGCGG-3′)和 ITS4(5′-TCCTCCGCTTATTGATATGC-3′),用BioEdit软件校对 DNA序列并去除引物和双峰序列,然后在NCBI核酸序列数据库中搜索同源性菌株(选相似度98%以上的模式菌株)(Landeweert et al.,2003),最终确定丝状真菌的分类地位。
1.4 数据处理与分析
利用R 4.0.2和Excel 2018软件计算丝状真菌的多样性指数、优势种的更替率(r)、优势度(Y)、出现频率(fi)以及生态位宽度(Bi)、生态位重叠(Oik),并对水环境因子的差异性、丝状真菌的多样性、优势种的时空异质性以及优势种与水环境因子的相关性进行分析。
1.4.1 多样性指数
通过计算丰富度指数(R)、Shannon-Wiener多样性指数(H')、Pielou均匀度指数(J')、Simpson多样性指数(D)来判断纳木措水体丝状真菌的多样性。

式中:
S——丝状真菌物种数;
Pi——i种的个体数与总个体数的比值;
N——某样点物种的总数。
1.4.2 优势种的划分及更替率
确定优势种采用McNaughton优势度计算方法(McNaughton,1967;Habib et al.,1997),优势种的季节更替率计算参考文献(张皓等,2016;梁淼等,2018)。
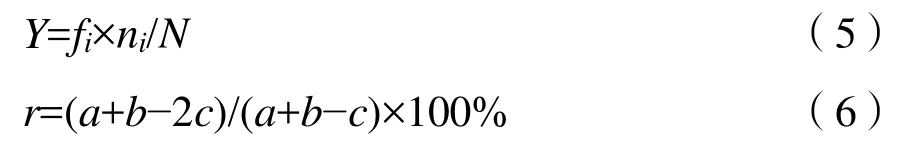
式中:
fi——i种的出现频率;
N——样点内的物种总数;
ni——i物种的总数;
a、b——相邻两季优势种的种类数;
c——相邻两季共有优势种的种类数,(Y>0.02为优势种,Y>0.1为绝对优势种)。
1.4.3 生态位
生态位宽度值的计算采用Colwell et al.(1971)加权修正的Levins指数。

生态位重叠值的计算采用Pianka指数(1973)。
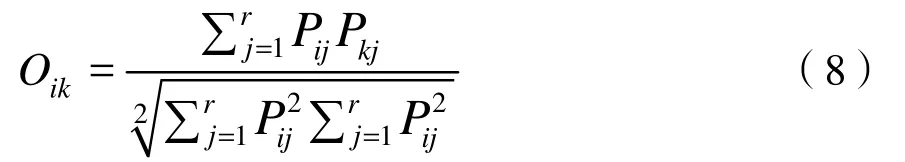
式中:
Bi——物种i的生态位宽度值;
Pij——空间生态位中代表i种在j样点的个体数占该样点总个体数的比例;
r——总样点数;
Pij——时间生态位中代表i种在j季节的个体数占i种在所有季节总个体数的比值;
r——总季节数;
Oik——物种i与物种k的生态位重叠指数,时空生态位宽度=时间生态位宽度×空间生态位宽度。
2 结果与分析
2.1 纳木措不同季节水体环境因子的差异性分析
对纳木措3个季节的8种水环境因子做差异性分析(图1)。pH在春季和夏季、春季和秋季间存在极显著差异(P<0.01),其中春季高于秋季、夏季;温度在3个季节间的差异均为极显著(P<0.01),且夏季>秋季>春季;NH4+-N在春季和秋季、夏季和秋季间呈显著差异(P<0.05);TP在春季和秋季间呈显著差异(P<0.05),在夏季和秋季间呈极显著差异(P<0.01);TN仅在春季和夏季间存在显著差异(P<0.05),以上3种环境因子均为春季最高,秋季最低,此外,TDS、EC和Salt在3个季节中均无显著差异。总体而言,湖水中的部分环境因子在季节更替中出现了显著变化。
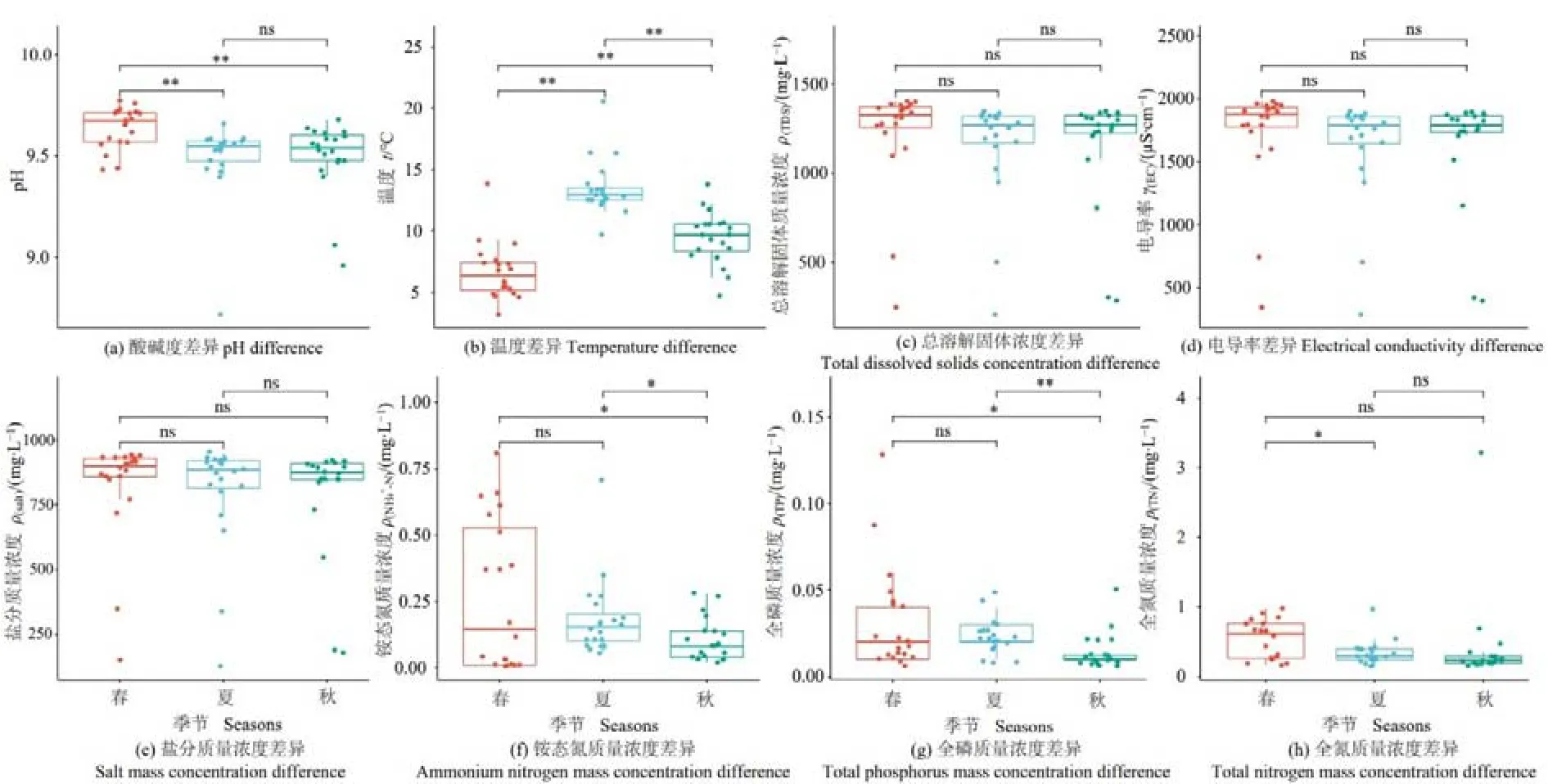
图1 纳木措不同季节水体环境因子差异Figure 1 Differences of water environmental factors in different seasons in Nam Co Lake
2.2 纳木措不同季节水体丝状真菌群落组成与多样性的比较
在纳木措3个季节中共鉴定出3359株丝状真菌,如图2a所示,丝状真菌总丰度在夏季与春季、夏季与秋季间呈极显著差异(P<0.01),且夏季最多,为1412株,其次为秋季(1026株),春季最少(921株)。丝状真菌物种数在秋季与夏季、秋季与春季间呈极显著差异(P<0.005),其中春季包括 20属62种,优势种9个,夏季包括22属47种,优势种13个,秋季包括13属47种,优势种7个。丝状真菌的多样性指数(图2b),除Shannon-Wiener多样性指数、Simpson多样性指数在春季与夏季间无显著差异外,其他季节间的各指数均存在显著(P<0.05)和极显著差异(P<0.01),其中秋季的Shannon-Wiener多样性指数、Pielou均匀度指数均为最高,春季的Simpson多样性指数最高。
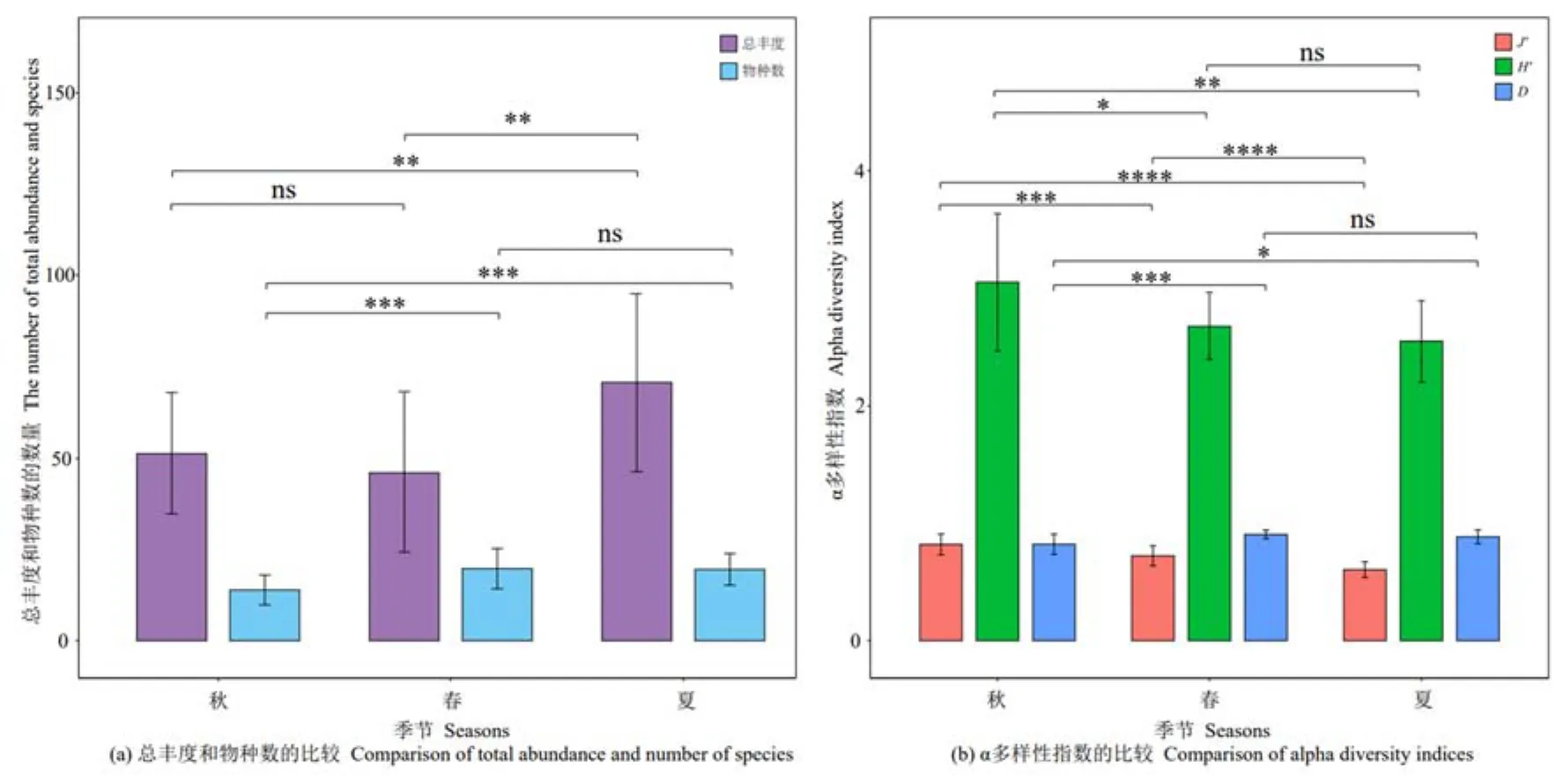
图2 纳木措不同季节水体丝状真菌多样性指数的比较Figure 2 Comparison of diversity indices of filamentous fungi in different seasons in Nam Co Lake
为进一步分析纳木措各季节水体丝状真菌优势种的时空异质性(图 3),采用 Bray-Curtis距离的非度量多维尺度(NMDS)分析,按季节分为 3组,发现秋季的2号和15号以及夏季的14号样点中优势种群落结构与本季节其他样点差异较大,其余各样点的优势种群落结构在同一季节内的差异较小。ANOSIM 结果显示,R=0.939,P<0.01,说明各季节间的差异显著大于季节内样点间的差异。
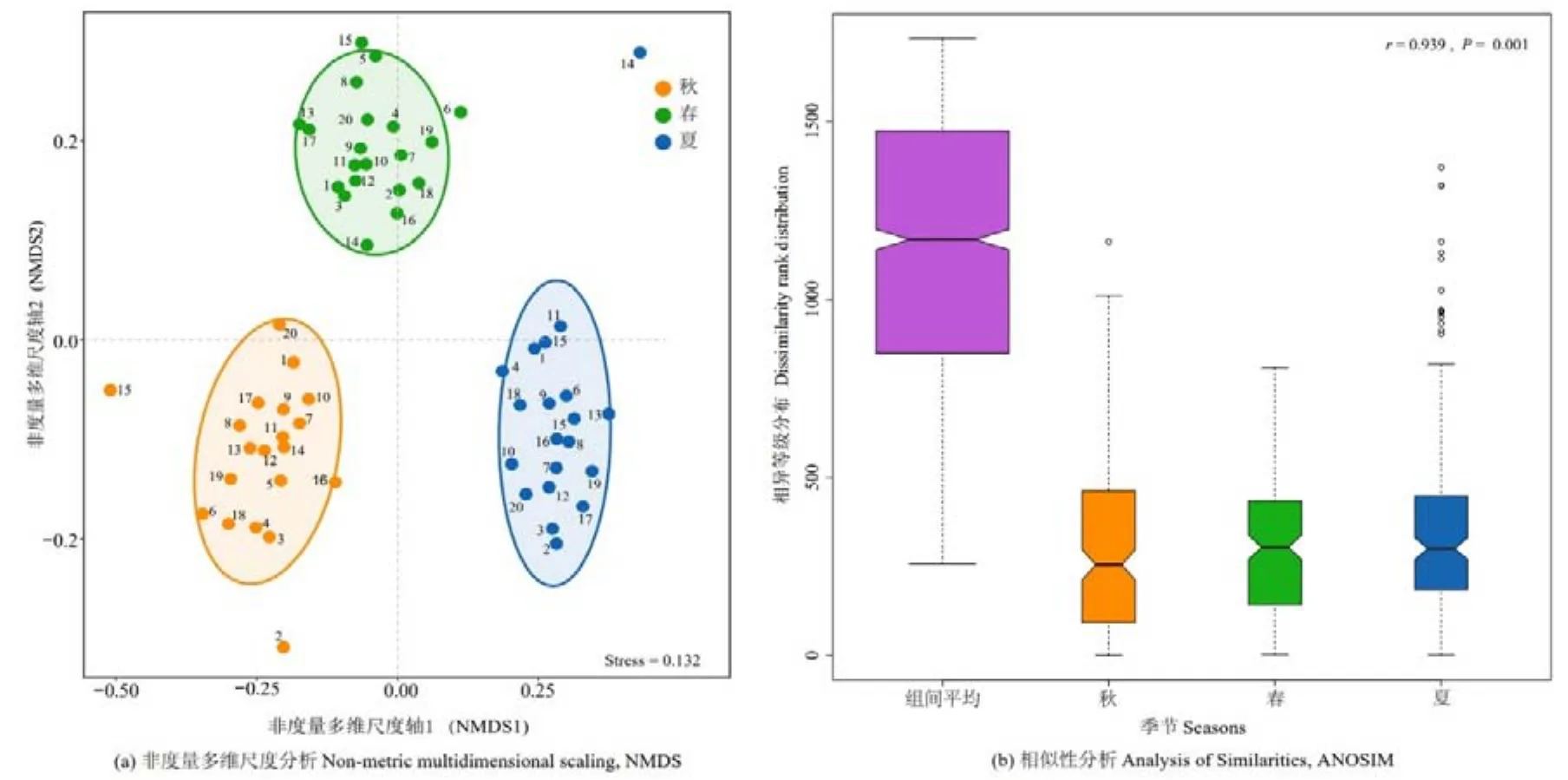
图3 纳木措不同季节水体丝状真菌优势种的空间变异分析Figure 3 Spatial variation analysis of the dominant species of filamentous fungi in different seasons in Nam Co Lake
2.3 纳木措不同季节水体丝状真菌优势种和更替率
对纳木措3个季节的水体丝状真菌优势种进行划分,共得到优势种23种,如表1所示,普通青霉Penicillium commune为3个季节的共有优势种,酒色青霉Penicillium vinaceum、冻土毛霉Mucor hiemalis为春季和夏季的共有优势种,壳青霉Penicillium crustosum、灰玫瑰青霉Penicillium griseoroseum为春季和秋季的共有优势种。春季和秋季有独有优势种4个,夏季有独有优势种10个。由春季进入夏季时,优势属Penicillium的占比下降,优势种的数量增加;由夏季进入秋季时,优势属Penicillium的占比上升,优势种的数量下降。经计算,夏季与秋季优势种的更替率最高,为95%,春季与夏季优势种的更替率次之,为84%,春季与秋季优势种的更替率最低,为77%,说明纳木措水体丝状真菌优势种的季节性变化较大。
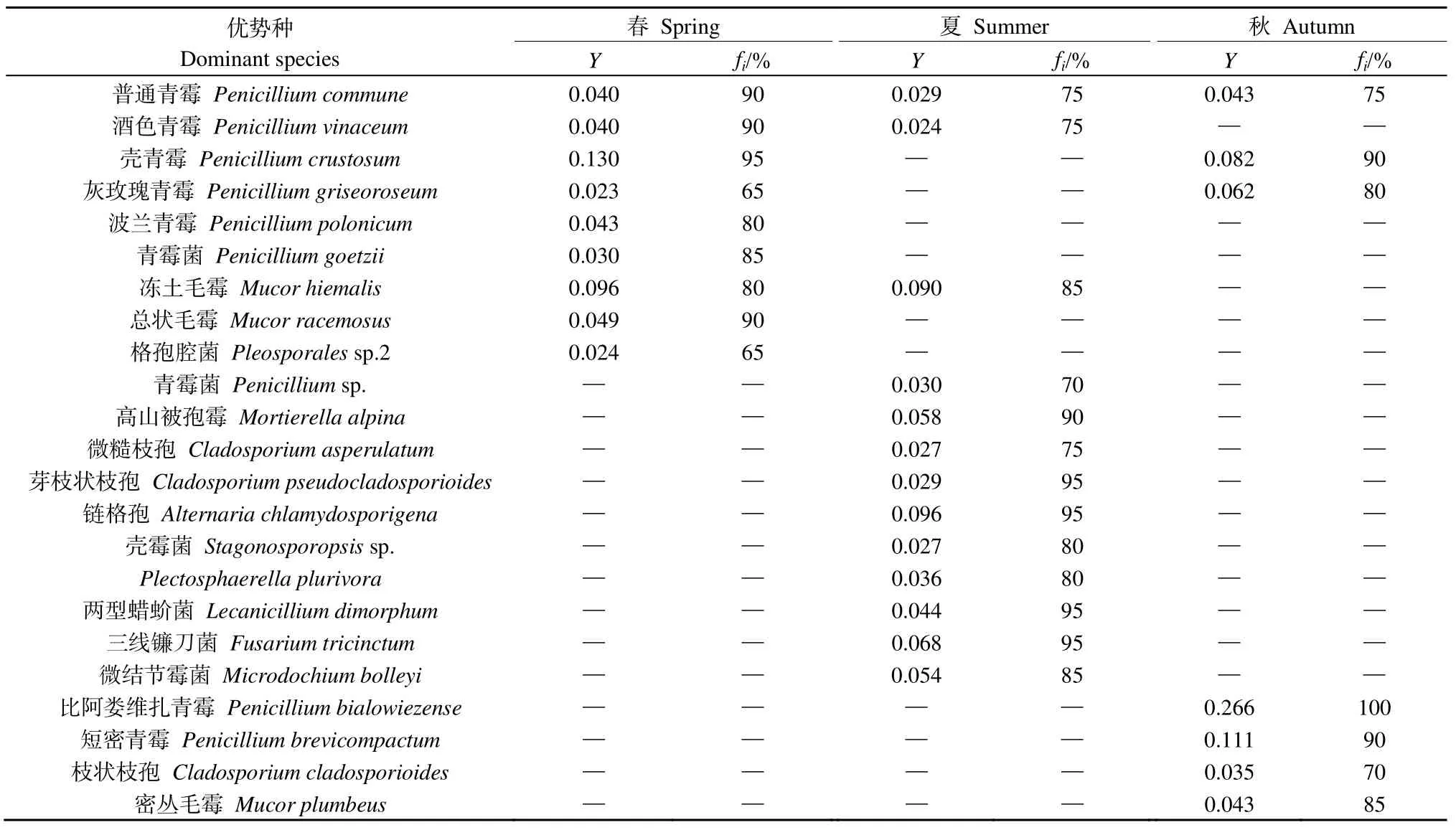
表1 纳木措水体丝状真菌优势种在不同季节的优势度(Y)与出现频率(fi)Table 1 Dominance (Y) and frequency (fi) of dominant species of filamentous fungi in different seasons in Nam Co Lake
2.4 纳木措不同季节水体丝状真菌优势种的生态位
如表2所示,纳木措水体丝状真菌优势种的空间生态位宽度值区间在 0.017—0.942,其中壳霉菌Stagonosporopsissp.的空间生态位宽度值最高,比阿娄维扎青霉Penicillium bialowiezense的空间生态位宽度值最低。时间生态位宽度值区间在 0.333—0.979,其中P.commune的时间生态位宽度值最高,波兰青霉Penicillium polonicum等19个菌种的时间生态位宽度值均较低。丝状真菌优势种的时空生态位宽度值均较小,区间在 0.006—0.314,其中P.bialowiezense的时空生态位宽度值最低,Stagonosporopsissp.的时空生态位宽度值最高,且处于窄生态位(0≤Bi<0.15)的菌种占比较大,这表明纳木措水体丝状真菌优势种易受时空变化的影响。
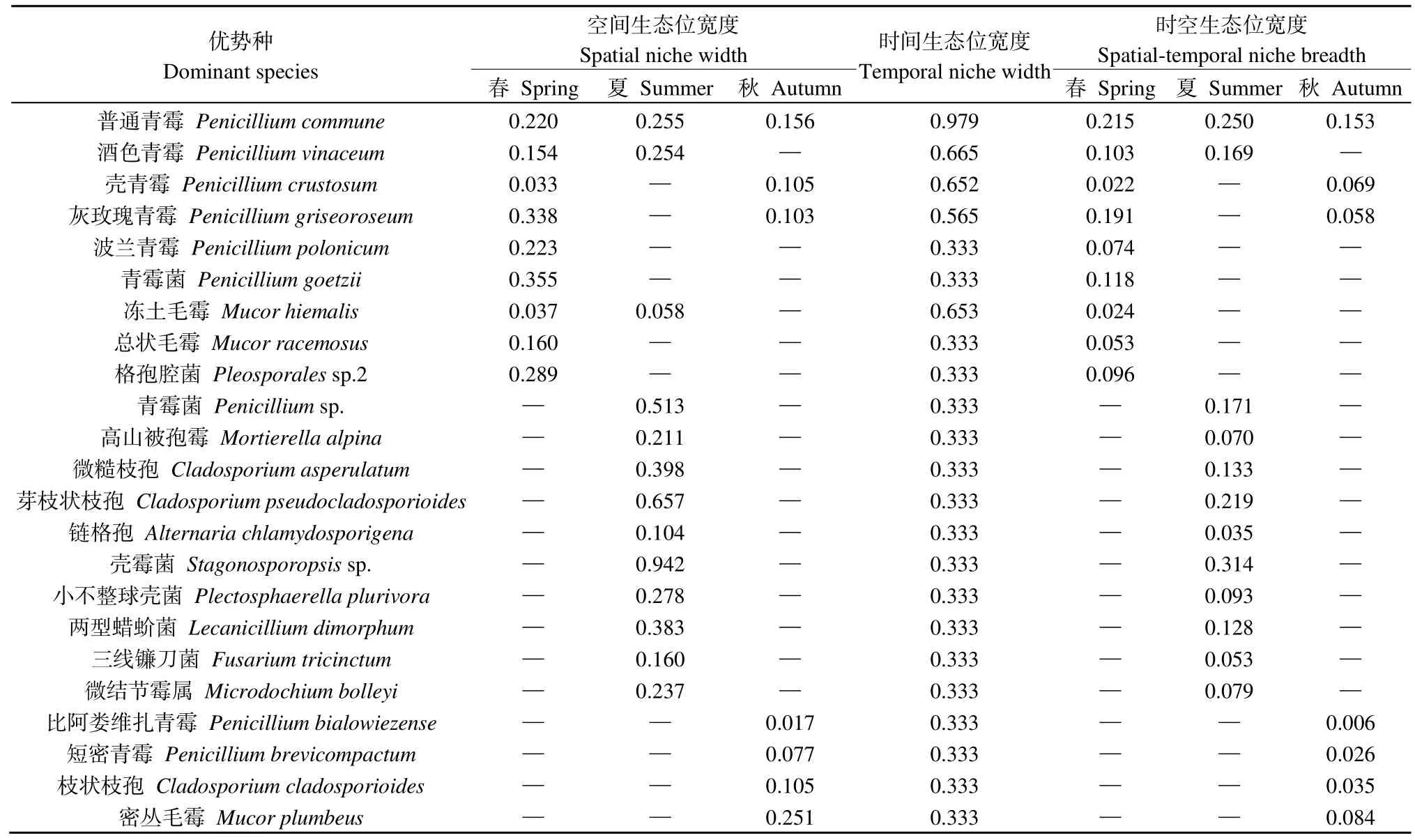
表2 纳木措水体丝状真菌优势种在不同季节的生态位宽度(Bi)Table 2 Niche width of dominant species of filamentous fungi in different seasons in Nam Co Lake
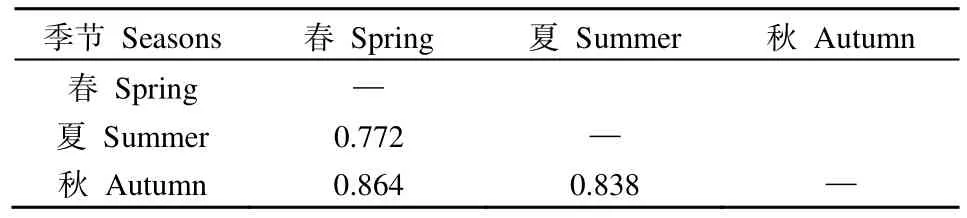
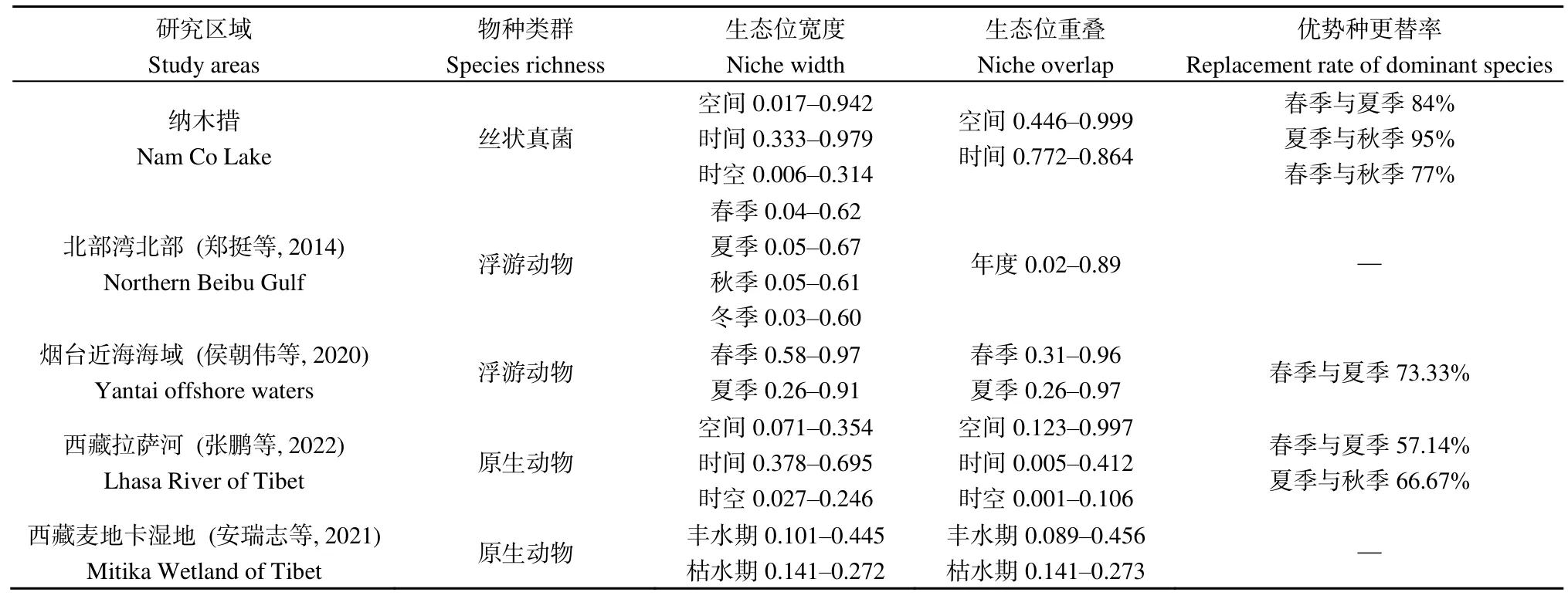
在空间维度上(表3),纳木措各样点丝状真菌优势种所组成的群落间的生态位重叠度较高,其中丝状真菌群落在160对样点间高度重叠(Oik>0.8),占总对数的 84%;在 24对样点间中度重叠(0.6 表3 纳木措各样点水体丝状真菌群落的生态位重叠(Oik)Table 3 Niche overlap of filamentous fungal communities at different sites in Nam Co Lake 表4 纳木错各季节水体丝状真菌群落的生态位重叠(Oik)Table 4 Niche overlap of filamentous fungal communities in different seasons in Nam Co Lake 分析纳木措不同季节的水环境因子,发现总溶解固体量、电导率和盐浓度在3个季节中均无显著差异,其他 5种环境因子在各季间存在显著(P<0.05)或极显著差异(P<0.01)。水环境因子与丝状真菌优势种的Spearman等级相关性分析(图4)显示,在春季,优势种P.vinaceum、M.hiemalis分别与全磷、铵态氮呈极显著负相关和极显著正相关(P<0.01)。在夏季,优势种Stagonosporopsissp.、高山被孢霉Mortierella alpina、M.hiemalis与电导率、盐浓度、总溶解固体量呈显著(P<0.05)或极显著负相关(P<0.01);优势种两型蜡蚧菌Lecanicillium dimorphum与温度、pH分别呈显著正相关和显著负相关(P<0.05),温度又与小不整球壳菌Plectosphaerella plurivora呈显著正相关(P<0.05),pH与微结节霉菌Microdochium bolleyi呈显著负相关(P<0.05);优势种微糙枝孢Cladosporium asperulatum与全氮、铵态氮呈显著负相关(P<0.05),同样全氮与链格孢Alternaria chlamydosporigena呈显著负相关(P<0.05);优势种M.hiemalis与全磷呈显著正相关(P<0.05),与铵态氮呈极显著正相关(P<0.01)。秋季,优势种枝状枝孢Cladosporium cladosporioides与盐浓度、电导率、总溶解固体量呈显著正相关(P<0.05),P.bialowiezense与铵态氮、pH以及P.griseoroseum与全磷均呈显著负相关(P<0.05)。 图4 纳木措不同季节水体环境因子与丝状真菌优势种丰度的相关性Figure 4 Correlation between water environmental factors and the abundance of dominant species of filamentous fungi in different seasons in Nam Co Lake 纳木措3个季节的水体中共鉴定出3359株丝状真菌,各季节的多样性均较高,且α多样性指数间大多存在显著差异(P<0.05)。时空异质性的结果表明,丝状真菌优势种在各季节间的差异显著大于季节内各样点的差异。有相似研究表明,生境差异能使湖泊中细菌群落的多样性和组成产生异质性(Jiao et al.,2020;Zhang et al.,2021);纳木措不同季节间丝状真菌的多样性和群落结构存在显著的时间异质性,且时间异质性大于空间异质性,各季节间环境因子的差异形成生境的差异,可能是造成这一结果的重要原因。丝状真菌优势种的数量和物种组成随着季节的改变发生显著变化,各季节优势种的更替率也较高,如其中的优势属Penicillium,由春季进入夏季,夏季进入秋季,其占比先下降后上升,这可能与Penicillium在低温环境中的优势更强有关(Cantrell et al.,2011)。结合不同季节间环境因子的变化幅度来看,环境的变化越大可能是群落越不稳定,更替率越大的重要原因。 生态位宽度是反映种群对环境适应能力和资源利用能力的重要指标(李德志等,2006;彭松耀等,2015)。在空间维度上,Stagonosporopsissp.的空间生态位宽度值最高,受空间资源的限制较小;P.bialowiezense的空间生态位宽度值最低,受空间资源的限制较大(何雄波等,2018)。在时间维度上,P.commune的时间生态位宽度值最高,受季节变化的限制较小;P.polonicum等19个菌种的时间生态位宽度值均较低,受季节变化的限制较大(黄保宏等,2005;马玲等,2012)。丝状真菌优势种的时空生态位宽度值均较小,且窄生态位的菌种占比较大,总体上纳木措水体丝状真菌优势种易受时空变化的影响。生态位重叠值反映着种群间对资源的竞争强弱(求锦津等,2018),纳木措各样点丝状真菌优势种所组成的群落间的生态位重叠度较高,在空间维度上,各样点丝状真菌优势种组成的群落高度重叠的占84%;在时间维度上,3个季节的丝状真菌优势种所组成的群落间的生态位重叠度同样较高(77.2%—86.4%)。表明各丝状真菌群落的竞争较为激烈,且群落结构不稳定正处于发展的状态。虽然不同季节的丝状真菌群落间不会直接产生竞争关系,但它们之间较高的生态位重叠正是构建各个季节不同群落结构的重要动力。总体而言,不同水域或不同季节塑造了水体中丝状真菌不同的群落结构,不同微生物种群对各种环境因子有着不同的耐受度和适应力(Williams et al.,2008),竞争优势随着环境变化而改变,群落发生演替,表现为各季节丝状真菌的多样性和物种组成产生异质性以及优势种在各季节间较高的更替率。 微生物群落的多样性和物种组成受到CO2、有机物、氮、磷和温度等各种环境因素的影响(Zhang et al.,2014;Li et al.,2019;Xia et al.,2020)。时空变化对丝状真菌群落结构的影响本质上是不同水域和不同季节的水环境因子与丝状真菌各种群相互作用的体现,组成微生物群落的各物种彼此之间对资源存在广泛的竞争(Esser et al.,2015;Hassani et al.,2018),环境的变化能直接影响它们的竞争力,因此,环境条件的变化造成了微生物群落的演替。相关研究表明水体真菌的组成和分布等受环境因子的影响(王艳红等,2021),本研究中,不同季节的优势种与水环境因子的相关性差别较大,如夏季的两个优势种与温度具有显著的正相关(P<0.05),且夏季湖水的温度显著高于春季和秋季,说明温度的升高可能是造成二者逐渐成为优势种的原因之一。 对环境响应敏感,受水体环境限制较强的某些原生动物类群作为环境指示种被认为有着可靠的生态学意义(汪志聪等,2010)。通过与其他学者相似的研究比较发现(表5),本研究中的丝状真菌优势类群对水环境的变化有着更敏感的响应,在季节变化中,群落的更替率和构成群落的物种变化更大。目前已有研究表明,青藏高原的变暖率远高于全球平均水平(Liu et al.,2000),全球变暖造成冰川消融增加,冰川融水的大量输入改变了高原湖泊的盐度、养分和生物多样性(Milner et al.,2009),这些水文条件的变化可能对湖泊生态系统的结构和功能产生严重影响(Lin et al.,2017)。且微生物对环境变化的高度敏感性,使得用生物方法评判水质和气候的细微变化成为可能,虽然微生物检测法需要在统一的标准下(如采样方法、培养基的种类、稀释倍数、培养环境等)才有可比性,但随着技术的发展和分离方法的完善,利用微生物的多样性、群落结构、生态位等信息作为环境变化的指标在未来具有巨大的应用潜力。 表5 纳木措微生物与其他区域微生物的生态位研究对比Table 5 Comparison of niche studies of microorganisms in Nam Co Lake with those in other regions (1)纳木措不同季节间丝状真菌的多样性和群落结构存在显著的异质性,时间资源比空间资源的影响更显著,各季节环境因子的差异以及不同时期环境因子与丝状真菌之间相关性的差异是驱动其群落构建的主要动力。 (2)在时空生态位宽度以及生态位重叠上,大多优势种处于窄生态位,且群落间重叠度较高,丝状真菌的群落结构不稳定,在不同季节的环境条件下,一些菌种的竞争优势随之改变,群落发生演替,塑造了各季节丝状真菌的多样性和组成差异,表现为优势种在各季节间较高的更替率。 (3)相比于其他类型的水体微生物生态位,丝状真菌优势类群对水环境的变化有着更敏感的响应,这种高度敏感性,可以用来评估水质和气候的微小变化,具有较大的应用潜力。
2.5 纳木措不同季节水体环境因子与丝状真菌优势种的相关性分析
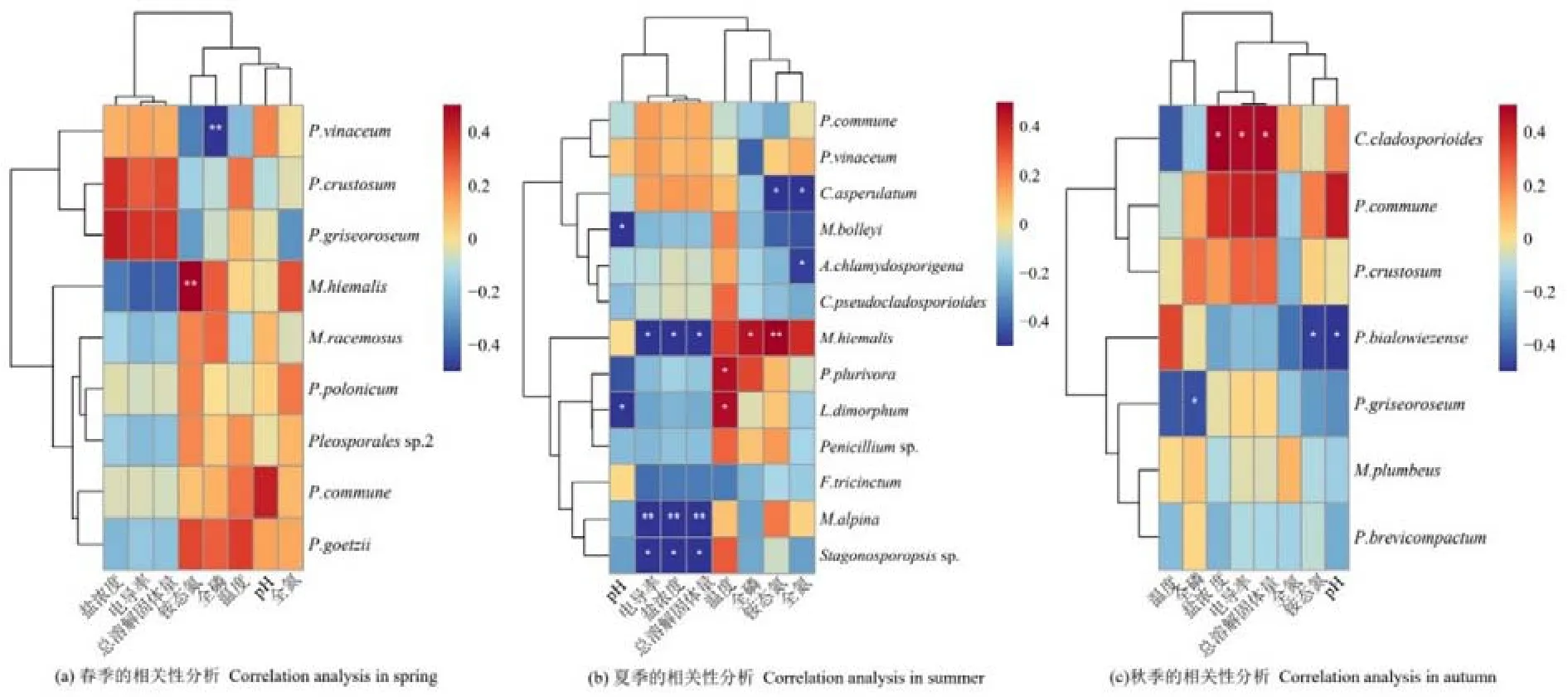
3 讨论
4 结论
猜你喜欢
生物化工(2022年4期)2022-09-20
昆明医科大学学报(2022年2期)2022-03-29
食品安全导刊(2021年20期)2021-08-30
公民与法治(2020年22期)2020-12-14
河南科学(2020年3期)2020-06-02
今日华人(2019年7期)2019-10-13
西藏人文地理(2015年4期)2016-03-04
中外医疗(2015年5期)2016-01-04
草业科学(2015年9期)2015-12-11
恋爱婚姻家庭·养生版(2015年10期)2015-05-14
