
江心洲景观类型和格局对植物多样性的多尺度影响
——以闽江流域福州段为例
2022-02-19 12:28玄锦李祖婵邹诚秦子博吴雅华黄柳菁
生态环境学报 2022年12期
玄锦,李祖婵,邹诚,秦子博,吴雅华,黄柳菁
福建农林大学风景园林与艺术学院,福建 福州 350000
近年来,景观格局对植物多样性的影响受到国内外学者的广泛关注。一方面,不同土地利用类型形成的景观格局可以对植物群落的组成及多样性进行有效地塑造和调节(Walz,2015)。国内外学者基于不同的土地利用类型进行研究,发现森林、农田、放牧用地、未利用地及建筑用地的形状、破碎程度等往往会对植物多样性产生较大的影响(Hubble et al.,2009;董翠芳等,2014;李羽翎等,2022)。另一方面,整体景观中不同的景观特征指数及其配置方式对植物多样性也有较大的影响(彭羽,2016)。景观斑块的面积(Tulloch et al.,2016;李祖政等,2018)、形状(Moser et al.,2002;Schindler et al.,2013;彭羽等,2020)等特征,以及破碎化(Amici et al.,2015)、连通性(Reitalu et al.,2012)等斑块的配置方式会直接影响植物多样性,并通过生物地球化学过程、生态系统的生物交互作用产生持续的间接影响(Amici et al.,2015)。此外,许多学者发现景观格局对植物多样性的影响存在空间尺度效应,景观格局与植物多样性的相关性随空间尺度的不同而表现不同(Higgins et al.,2012;Schindler et al.,2013;Mõisja,2016;彭羽,2016),小尺度景观格局也对植物多样性有显著影响(Mõisja et al.,2016),多尺度下景观格局指数对物种的影响尚未形成统一的结论。因此,探讨不同尺度下景观格局指数对植物多样性的影响模式十分重要。
闽江流域是中国流域生态补偿重要试点之一(Zhang et al.,2020),为多种动植物提供生长、繁殖的栖息环境,在生物多样性保护中发挥重要作用。流域内江心洲分布较广,其作为河流湿地生态系统组成部分,对防止河岸水土流失、净化水质、调节微气候、维持生态系统结构和功能的动态平衡有重要作用(Hubble et al.,2009)。然而,受洪水、潮汐等自然因素和过量采砂、水口电站蓄水拦沙等人为因素的长期影响,江心洲河岸被严重侵蚀,近十余年来江心洲岸线呈逐渐缩小趋势(马宇龙等,2017;尹久娜等,2019),生物多样性以及河岸带的自然植被受到不同程度的破坏,生态环境已变得极为脆弱(David,2005;Mendez et al.,2014)。此外,江心洲内部农业种植活动较频繁,许多自然土地被侵占为农田、牧场,也有部分土地用来做经济林的培育基地,这大大改变了原有的景观格局,江心洲内部原生植被面积大大减少。目前,由于江心洲原生植被呈现斑块状分布,且缺乏不同尺度下景观格局对其植物多样性影响的探讨,导致对江心洲的认识和科学管理缺乏理论资料,使得对江心洲生态系统的保护和植被恢复工作困难重重。
因此,本研究以闽江流域福州段的5个江心洲为研究区,对江心洲景观格局与植物多样性的相关性进行研究,揭示江心洲景观格局对植物多样性的生态影响效应,进而丰富景观格局对植物多样性影响的理论成果,因地制宜地为闽江流域水环境保护制定具有针对性的参考建议,以期为闽江流域生态保护修复工作提供新视角。
1 研究区域与研究方法
1.1 研究区概述
闽江流域福州段位于闽江流域下游,起于闽清县雄江镇,止于连江县琯头镇长门口,长度约 125 km,是“山水城市”的重要载体,也是福州重要的发展廊道与景观、生态轴线。流域内多低山、丘陵地形,河流水系众多。该区属亚热带季风气候,台风频发,常见极端暴雨天气,具有明显的潮汐效应。流域内经济发达,人地矛盾导致生态系统功能退化,水土流失、水环境污染等生态问题多发,沿岸植物多样性破坏严重。
选择闽江流域内中房洲、绿洲、古山洲、厚美沙洲和大洋鹭洲作为研究区(图1),位于闽侯大桥与橘园洲大桥(26°3′9″—26°7′6″N,119°11′5″—119°13′29″E)之间,面积分别为 65.51、20.83、50.65、72.26、35.32 hm2,植被资源相对丰富。5个江心洲受人类活动影响较为明显,其中放牧、种植等农业活动对江心洲影响较大,且存在撂荒现象,生态问题较为明显。
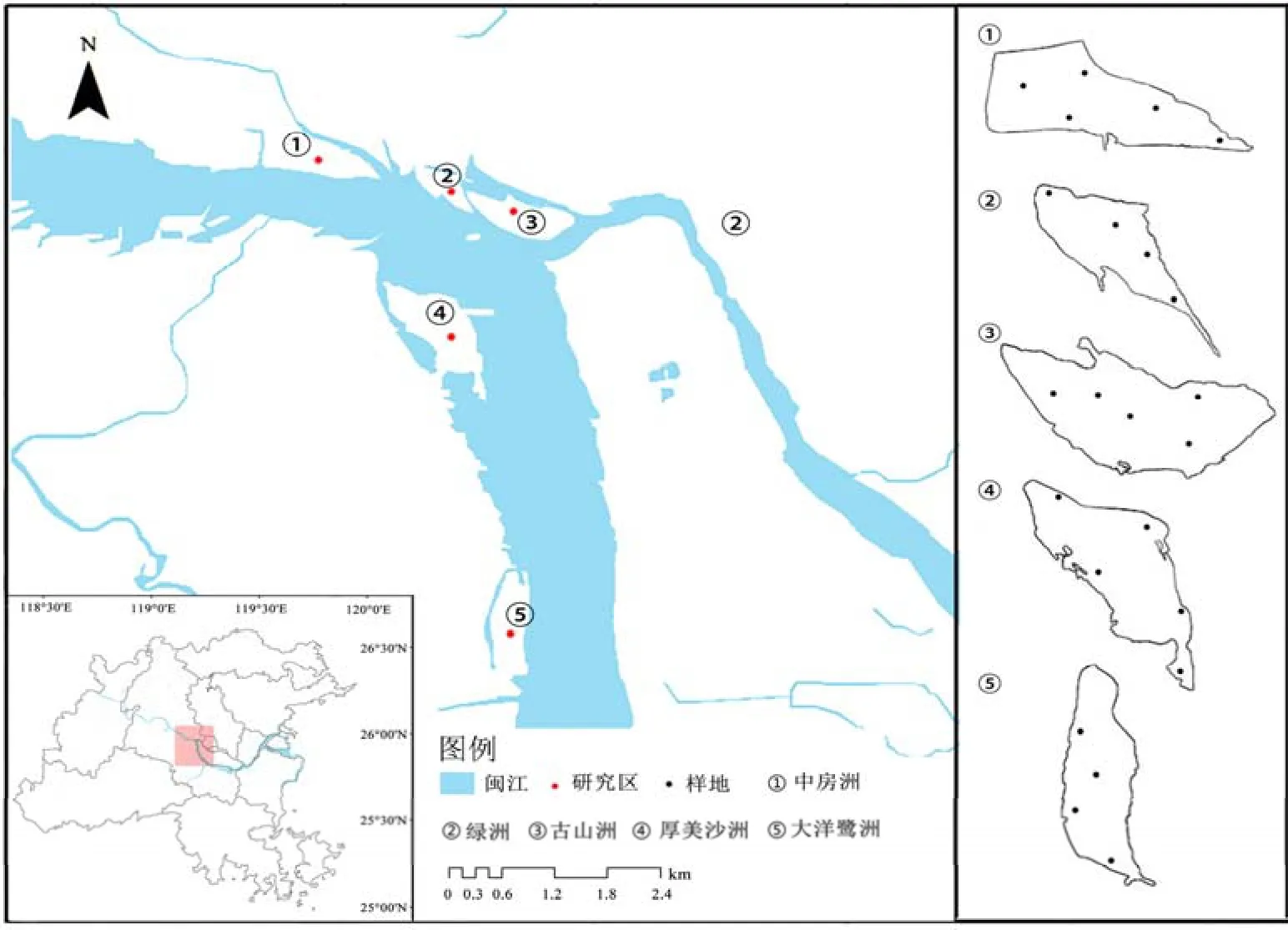
图1 研究区地理位置及样地分布示意图Figure 1 Location of the study area and the distribution of sample points
1.2 研究方法
1.2.1 植物数据采集
2021年10—12月对5个江心洲内的植物进行群落调查,每个江心洲选择 4—5个受人为干扰较小的样地,样地之间距离至少100 m,共23块(图1)。每个样地共设置4个5 m×5 m乔木样方、8个2 m×2 m的灌木样方,12个1 m×1 m草本样方,记录每个调查样方内乔木的名称、高度、枝下高、胸径、冠幅、数量;灌木的名称、高度、冠幅、基茎、盖度及草本植物的名称、高度、盖度(Aggemyr et al.,2012)。植物调查的基础上,记录江心洲的植物组成并计算每个样地内的 Shannon-Wiener指数(H)、Simpson指数(D)和Pielou指数(J)3个植物多样性指数,计算公式如下(李羽翎等,2022):
工程措施包括:排水沟516.5m,土地整治5.70hm2,铅丝石笼防护1 125.05 m,浆砌石护坡585 m3,浆砌石挡墙 1 200 m3,覆土面积 5 908.31 m3,土石方开挖3 127.45 m3,回填 1 238.4 m3。

式中:
Pi——样地内第i个物种的多度占总物种多度之和的百分比;
即:

其中:
Ni——第i种的个数;
N——所有物种的个数;
S——样方内物种种类数。
1.2.2 景观数据采集
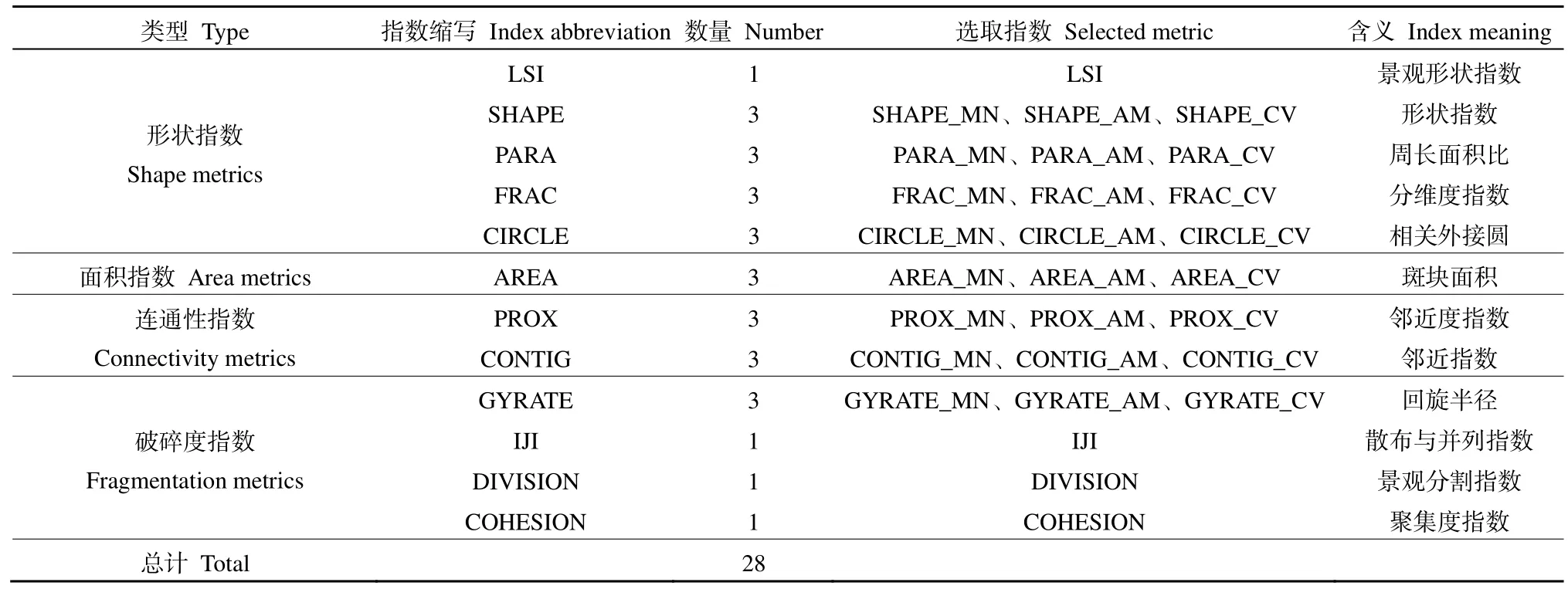
表1 本研究选取的景观格局指数Table 1 Selected landscape pattern metrics in the study
1.2.3 数据处理
通过Canoco 5软件中的冗余分析(RDA)研究江心洲景观格局指数与植物多样性指数的相关性,在每个景观尺度下分别以景观水平和类型水平下景观格局指数为解释变量,以植物物种多样性指数为因变量进行分析,筛选出每个尺度下对植物多样性最具有解释能力的景观格局指数。应用SPSS 24软件计算各尺度内计算主要景观指数与植物多样性的Pearson相关系数。为消除数据的共线性,在数据处理前对所有原始数据进行lg(x+1)转换(侯朝伟等,2020)。
2 结果与分析
2.1 江心洲植物组成特征
在本次调查的闽江流域江心洲 23个样地中,共记录123种植物,隶属于62科96属。其中,草本植物共99种,占比为80.5%;灌木11种,占比8.9%;乔木13种,占比10.6%。通过生活型占比可以看出,研究区的植物种类以草本为主。各植物样方的植物多样性指数如表2所示。

表2 各样地植物多样性指数Table 2 Plant diversity index of each sample sitet
2.2 景观类型水平指数与植物多样性的关系
根据图2和表3,研究区内优势景观类型为林地,总面积达 157.55 hm2,占研究区总面积的64.49%;其次为草地和未利用地,江心洲的草地多为放牧用的草场及撂荒之后形成的草地,未利用地多为沿河岸带的沙地、滩涂草地及荒草地,面积分别为47.32、31.78 hm2,分别占总面积的19.37%、13.01%;农田面积为6.53 hm2,占总面积的2.67%;建筑用地和水体的面积均不足1 hm2,江心洲建筑用地多为依附于农田、林地的临时建筑,分布较为分散,而水体方面仅在2个江心洲内有较稳定的分布,二者占研究区面积均在1%以下。
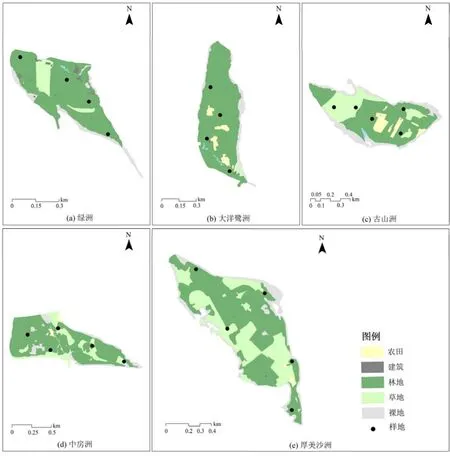
图2 各江心洲景观类型及样方分布图Figure 2 Landscape type and sample distribution distribution of middle bar

表3 江心洲景观类型面积占比Table 3 Proportion of landscape type area in Jiangxinzhou
由图3和表4可知,在不同尺度下,对植物多样性影响最大的景观类型及景观指数不同。在100 m尺度范围内,景观类型格局指数对植物多样性的影响排序为FSHAPE_AM>FFRAC_AM>USHAPE_AM,排序轴间夹角为钝角,相关系数为−0.591—−0.606,均呈显著负相关。据此推测在100 m尺度范围内,林地和未利用地的形状指数对植物多样性具有抑制作用。在200 m尺度范围内,景观类型格局指数对植物多样性的影响排序为ACIRCLE_MN>AAREA_MN>AGYRATE_MN,与植物多样性排序轴夹角皆为钝角,且相关系数为−0.497—−0.735,呈现显著负相关,表明在此范围内,农田对植物多样性影响较大,其形状越规则,面积越大,破碎度越高植物多样性越小。在300 m尺度范围内,景观类型格局指数对植物多样性的影响排序为 AAREA_MN>BPARA_MN>AAREA_AM>APARA_MN,其中 AAREA_MN、BPARA_MN、AAREA_AM与植物多样性排序轴箭头夹角钝角,且相关系数为−0.775—−0.156,呈负相关;APARA_MN与3个植物多样性排序轴箭头夹角为锐角,且相关系数为 0.545—0.639,均呈显著正相关。表明在此尺度下农田的面积越小、形状越复杂,对植物多样性的影响越显著,建筑形状越复杂,植物多样性越低。在400 m尺度范围内,景观类型格局指数对植物多样性的影响排序为 AAREA_MN>BAREA_MN>USHAPE_AM,且均与植物多样性排序轴夹角呈钝角,相关系数为−0.644—−0.353,均呈负相关。表明在此尺度下,农田的面积越大,植物多样性越小,建筑用地的面积和未利用地的形状对植物多样性有一定的阻碍作用。在500 m尺度范围内,景观类型格局指数对植物多样性的影响排序为 AAREA_MN>UPROX_AM>APARA_MN。AAREA_MN与多样性指数排序轴夹角为钝角,且相关系数为−0.752—−0.573,呈负相关关系;APARA_MN、UPROX_AM与植物多样性排序轴箭头夹角为锐角,且相关系数为 0.450—0.670,因此呈显著正相关。表明农田面积对植物多样性有抑制作用,此尺度下农田的形状复杂程度和未利用地的连通性对植物多样性有促进作用。在600 m尺度,景观类型格局指数对植物多样性的影响排序为UPROX_MN>UPARA_CV>UPROX_ AM,均与植物多样性排序轴箭头夹角为锐角,且相关系数为0.431—0.670,呈显著正相关,表明此尺度下未利用地的连通性和复杂程度有利于植物的多样性。
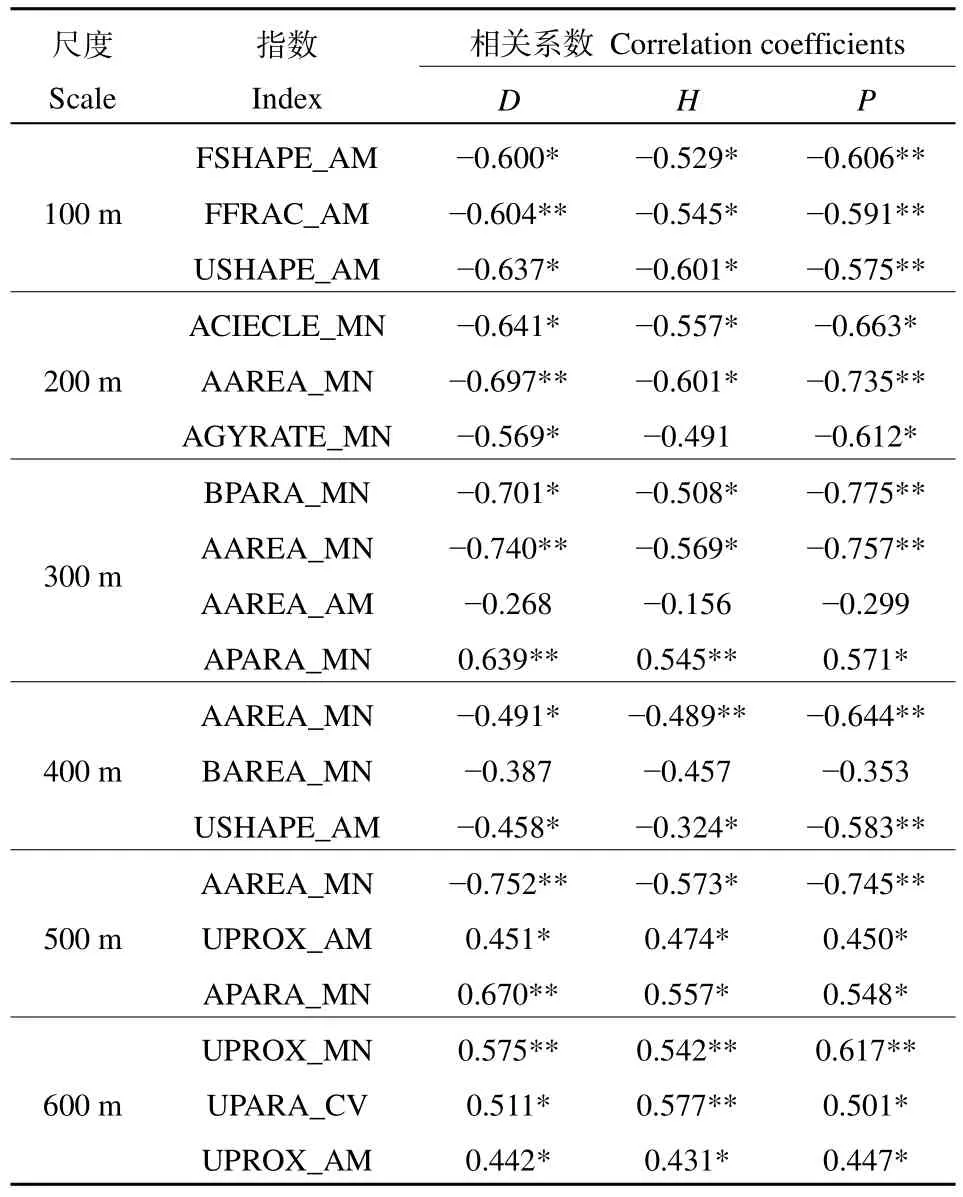
表4 不同尺度下类型水平指数对植物物种多样性影响的相关系数Table 4 Correlation coefficients between landscape-class leve metrics and plant species in six spatial scales
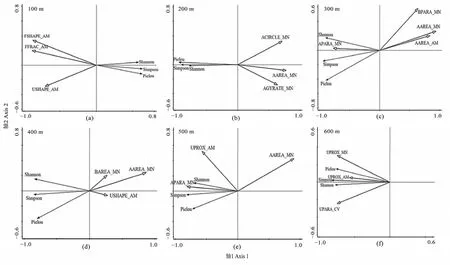
图3 不同尺度下类型水平指数与植物物种多样性的冗余分析Figure 3 RDA between landscape-class leve metrics and plant species in six spatial scales
总体上,农田、未利用地、林地和建筑用地的景观格局指数给植物多样性指数造成影响,其中,农田的面积指数AAREA_MN在200—500 m尺度下对植物多样性均有较大影响,其相关系数为−0.75—−0.489,呈显著负相关,对植物多样性有抑制作用;农田的形状指数在200、300、500 m尺度下与植物多样性呈显著相关,在200 m尺度下,形状规则指数ACIECLE与植物多样性相关系数分别为为−0.641、−0.557、−0.663,形状复杂指数 APARA_MN在300 m和500 m相关系数为0.545—0.670,因此可以推断农田形状越复杂,植物多样性越高。
未利用地的形状指数植物多样性的影响相对较小,且非正向的;连通性在500—600 m尺度连通性指数与植物多样性指数呈显著正相关(0.442—0.617)。建筑用地的形状和面积分别在300 m和400 m尺度显现出对植物多样性的抑制作用(−0.775—−0.353);林地的两个形状指数仅在100 m尺度下对植物多样性有影响,与植物多样性的相关系数为−0.529—−0.606,均为显著负相关。因此农田的形状和面积指数对江心洲植物多样性的影响更明显。
2.3 景观整体格局与植物多样性的关系
通过 RDA分析,得出不同尺度下主要景观指数对植物多样性指数的总解释量分别为 47.9%、39.0%、50.3%、73.6%、56.0%、51.7%,其中在400 m尺度范围内,景观格局对植物多样性具有最高的解释量。
根据表5和图4,在100 m尺度主要影响指数排序FRAC_MN>SHAPE_MN>IJI>CIRCLE_CV>PARA_AM>LSI>CONTIG_AM。其中CIRCLE_CV、CONTIG_AM与多样性排序轴箭头之间为锐角(图4),且相关系数为 0.554—0.064,为正相关;IJI、FRAC_MN、SHAPE_MN、PARA_AM与多样性排序箭头夹角为钝角,且相关系数为−0.556—−0.189,因此为负相关。在200 m尺度下,主要影响指数排序为FRAC_MN>CIRCLE_MN>SHAPE_MN>IJI>PROX_CV。PROX_CV与植物多样性排序轴箭头夹角为锐角,且相关系数为 0.105—0.245,因此呈正相关;FRAC_MN、CIRCLE_MN、SHAPE_MN、IJI与植物多样性排序轴箭头夹角钝角,且相关系数为−0.409—−0.258,因此为负相关。对于300 m尺度,主要影响指数的排序为FRAC_MN>CIRCLE_MN>IJI>SHAPE_MN,与植物多样性指数均夹角均为钝角,且相关系数−0.559—−0.225,呈负相关。在此尺度下,景观斑块的形状、破碎度会影响植物的多样性。在 400 m尺度,主要影响指数排序为SHAPE_MN>FRAC_MN>CONTIG_CV>GYRATE_CV>PARA_MN>IJI>CIRCLE_AM。CONTIG_CV、PARA_MN、GYRATE_CV与3个植物多样性指数排序轴均为锐角,其相关系数为 0.512—0.601,因此为正相关;SHAPE_MN、CIRCLE_AM、FRAC_MN、IJI与多样性指数排序轴箭头夹角为钝角,其相关系数为−0.660— −0.126,呈负相关。在500 m尺度下,对植物多样性影响较大的景观格局指数分别为 FRAC_MN>SHAPE_MN>PARA_MN>IJI>CIRCLE_MN。SHAPE_MN、CIRCLE_MN、FRAC_MN、IJI与植物多样性指数排序轴箭头夹角为钝角,且相关系数为−0.550—−0.168,因此呈负相关;PARA_MN对植物多样性排序轴箭头夹角为锐角,且相关系数为0.297—0.424,因此为正相关。在600 m尺度下,主要影响指数排序为 PARA_MN>SHAPE_MN>CONTIG_MN>GYRATE_CV>FR AC_MN>SHAPE_AM>PARA_AM>CONTIG_AM。PARA_MN、GYRATE_CV、PARA_MN、GYRATE_CV与表征植物多样性的 3个指数排序轴箭头间夹角为锐角,且相关系数为 0.102—0.580,都呈正相关;FRAC_MN、CONTIG_MN、SHAPE_MN与植物多样性指数排序轴夹角为钝角,且相关系数为−0.577—−0.136,呈负相关。因此,在所有尺度上形状指数对植物多样性的影响最大。其中,FRAC_MN、SHAPE_MN在100—600 m尺度上均与植物多样性呈负相关,相关系数为−0.641—−0.328,表明景观斑块的复杂程度对植物多样性有较为稳定的影响。FRAC_MN和SHAPE_MN在所有尺度上均具有统计学意义,可以用来比较它们在不同空间尺度上对植物多样性的影响。表5可以看出,在400 m尺度下相关系数最大,相关系数为−0.660—−0.557,且该尺度下景观指数对植物多样性指数的总解释量最高(73.6%),因此 400 m尺度为研究 FRAC_MN和SHAPE_MN对江心洲的植物多样性最适宜的尺度。

图4 不同尺度下景观水平指数与植物物种多样性的冗余分析Figure 4 RDA between landscape level metrics and indigenous plant species in six spatial scales
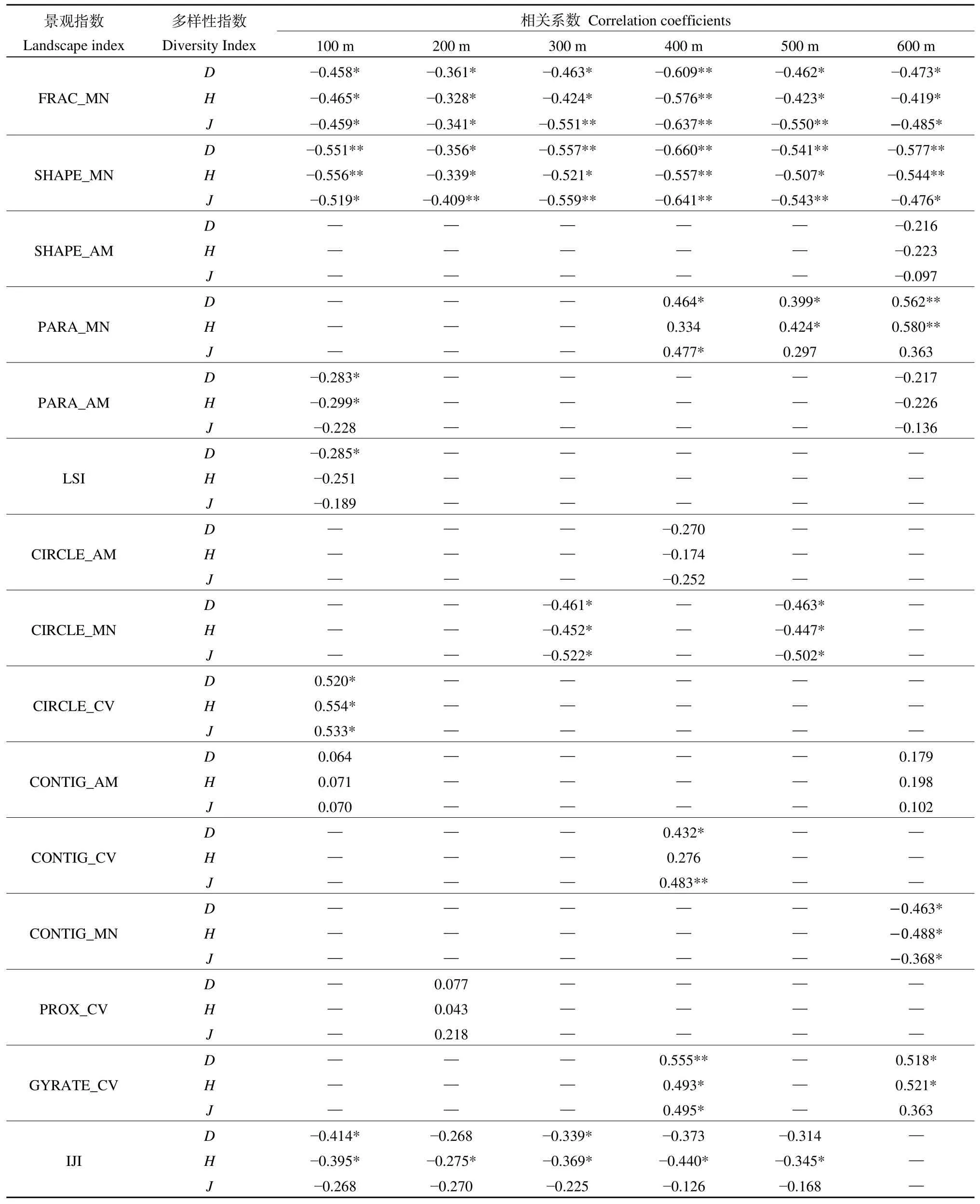
表5 主要景观指数的Pearson双变量分析Table 5 Pearson’s correlation coefficients of of major landscape indices
破碎度指数对植物多样性也有较为显著的影响,其中IJI在100—500 m尺度下对植物多样性均与植物多样性呈负相关,其相关系数为−0.414—−0.126,表明斑块分布分散且与其他斑块相邻多对植物多样性有一定抑制作用。连通性指数在 100、200、400、600 m尺度对植物多样性有较小影响,且相关性不稳定。
3 讨论
3.1 景观类型格局对植物多样性的影响
农田、林地、草地及未利用地是江心洲植物的重要生境,其斑块面积大小、形状复杂程度及连通性对江心洲植物多样性有重要影响。
农田的形状指数和面积指数都对植物多样性有较大影响。本研究中多个尺度下江心洲农田形状的复杂程度与植物多样性指数多呈正相关,这与Honnay et al.(2003)、Poggio et al.(2010)、董翠芳等(2014)学者的结论相一致。这是因为某些边缘、形状不规则的农田栖息地斑块的环境梯度数量更多(Honnay et al.,2003),本研究中江心洲的农田由于种植活动的不规范,其形状较多样。人为管理活动(除草、施肥等)往往会使农田形状趋于规则,因此通常形状复杂的农田斑块拥有更丰富的植物种类。农田面积指数与植物多样性在200—500 m尺度下呈现负相关,面积越大植物多样性越少,人们在农田中锄草、喷洒农药等种植活动影响了植物多样性,随着面积的增大,这种人为的影响会增加。
本研究中未利用地复杂程度与植物多样性指数呈负相关,这是因为常用的景观指数很难处理线性元素,常常把狭窄、细长的景观要素定义为具有高度复杂形状的要素(Moser et al.,2002)。而江心洲的未利用地多分布于河岸带,因此未利用地多为分布较狭长,容易被识别为高度复杂的形状。此外河岸带作为陆域与水域的生态交错带(魏雯等,2018),其淹水期、落干期的交替作用对植物特别是草本植物生长影响严重(陈丽慧等,2022),因此未利用地的植物数量及种类较少,多为荒草地、沙地和沼泽地,所以二者呈现负相关关系。未利用地的连通性在500—600 m尺度下对植物多样性具有促进作用,可能是由于未利用地在小尺度内斑块间距离更近,受管理活动的影响小,因此斑块连通性更强(Reitalu et al.,2012),此时连通性对植物多样性的影响表现不明显。而随着空间尺度增大,江心洲管理活动(道路、树篱等)的影响使连通性下降,受影响的斑块其物种数量及传播距离被限制,因此在较大尺度下植物多样性对连通性的抑制作用会更显著。
林地形状复杂度指数在100 m尺度范围内与多样性指数影响较大,这可能是由于斑块面积太小,整个斑块可能都会受到边缘效应的影响(Zbigniew et al.,1988)。而在范敏(2018)的研究中,100 m尺度下,景观多样性和形状指数对植物多样性影响显著。植物多样性与不同景观类型的形状指数的相关趋势并不一致(Schindler et al.,2013),因此形状指数与物种多样性的关系也需要进一步研究。
此外,许多研究显示建筑用地侵占了林地、草地等自然、半自然地表,其面积、形状及连通性会导致破坏植物生境,进而对植物多样性产生影响(Schmidt et al.,2014;Zhang et al.,2017)。本研究中江心洲建筑用地的形状复杂指数在 300—400 m尺度范围对植物多样性有较弱且不稳定影响,但是其与植物多样性相关并不明显,这是由于江心洲内搭建的临时建筑布局较为分散,且往往是依靠农田建设,因此其产生的影响并不明显,但是可能会在较小尺度内会对江心洲的植物多样性产生一定的影响。
3.2 景观整体格局对植物多样性的影响
不同尺度下表征斑块复杂度的指数对植物多样性都有较大的影响,FRAC_MN和 SHAPE_MN与植物多样性的关系均呈负相关,这可能是因为江心洲内林地这一种景观类型分布较广,且受其他类型的干扰很小,其基质连通性高,边界呈较为明确规则,植物物种丰富。其他研究中斑块形状复杂度对植物多样性的影响尚无确切定论,在不同尺度上、不同生态系统景观格局与植物多样性的相关性均有所不同。本研究仅针对江心洲这种特殊的生态系统得出结论,后续可以进一步研究复杂度指数与植物多样性的关系,确定复杂度指数如何作为优势指数对江心洲植物产生影响,研究如何根据复杂度来预测植物的多样性。
表征景观破碎度和连通性的指数与在多个尺度下对植物多样性有较小影响。本研究中,林地、草地斑块中镶嵌着农田和建筑这类人工形成的斑块,这些斑块使江心洲景观朝着破碎化发展。同时道路、树篱等线状廊道的出现使得斑块间连通性下降,导致栖息地内物种减少。许多研究显示,景观破碎化指数会随着景观尺度增加逐渐成为影响植物多样性的重要影响因子(Luis,2005;Amici et al.,2015)。但景观破碎化对物种多样性的影响可能是正向的也可能是反向的。一方面,景观破碎化会形成小的生境斑块,使生境斑块越来越独立,而斑块上种群的数量小,就会导致部分物种灭绝(Honnay et al.,2003;Aggemyr et al.,2012;Ramalho et al.,2018);另一方面,景观破碎化为植物生长提供了多样化的生境,这在一定程度上会提高景观内物种的迁入率,从而提高物种的多样性(Tscharntke et al,2012)。因此可以对新产生的生境斑块进行改造,使之成为自然、半自然的生境,促进斑块间的物种流通,从而提高植物多样性。
4 结论
通过对闽江流域 5个江心洲景观格局植物及植物多样性在指数的相关性分析,验证了江心洲景观类型及格局与植物多样性间存在相关性。结果如下:
在类型水平上,(1)未利用地、林地和建筑用地的景观格局指数会给植物多样性指数带来影响。(2)农田面积在200—500 m尺度下均对植物多样性有最大影响,且对其有抑制作用;农田的形状指数在200、300、500 m尺度下对植物多样性有影响;
在景观水平上,(1)景观格局的形状指数FRAC_MN、SHAPE_MN在所有尺度上均与植物多样性呈显著负相关;400 m尺度下 FRAC_MN和SHAPE_MN可以更好地指示江心洲的植物多样性。(2)破碎度指数在100—600 m尺度下对植物多样性也有一定影响,其中IJI在100—500 m尺度下均与植物多样性呈负相关。(3)连通性指数在 100—600 m尺度对植物多样性有较小且不稳定的影响。
因此,在对江心洲进行保护和建设时,应该在考虑到景观格局尺度效应的同时,根据需要对景观格局进行优化,以保护和增加植物多样性。
致谢:感谢吴怡婧、刘佳佳、范娟同学在江心洲植物调查工作中提供的帮助。
猜你喜欢
中学生天地(A版)(2022年11期)2022-11-25
中学生数理化·七年级数学人教版(2022年11期)2022-02-14
科普童话·学霸日记(2020年1期)2020-05-08
今日农业(2019年13期)2019-08-12
人民调解(2019年5期)2019-03-17
小天使·一年级语数英综合(2019年2期)2019-01-10
新世纪智能(英语备考)(2018年11期)2018-12-29
小天使·五年级语数英综合(2016年12期)2016-12-09
小说月刊(2015年12期)2015-04-23
中国火炬(2009年9期)2009-07-24
