
lncRNA TUG1靶向TGF-β/Samds信号通路促人牙周膜成纤维细胞纤维化的作用机制研究
2022-02-17 08:03孙海涛冯小东首都医科大学附属北京世纪坛医院北京100038
中国免疫学杂志 2022年1期
孙海涛 高 涛 冯小东 (首都医科大学附属北京世纪坛医院,北京 100038)
人牙周膜成纤维细胞(human periodontal ligament fibroblasts,HPLDFs)位于牙周膜组织胶原纤维中的间质细胞,在牙周组织的发育、损伤修复等过程中发挥关键调节作用[1]。正常生理状态下,HPLDFs 可促进合成和降解胶原,分泌多种细胞活性因子、刺激成骨细胞和破骨细胞以完成牙周组织的修复与再生,并促进牙槽骨及牙本质胶原不断更新和重塑以维持牙槽骨的稳态[2-3]。受外界刺激时,HPLDFs 的生物学活性异常活化,激活并促进牙周组织损伤,最终引起牙周结缔组织纤维化及牙槽骨病变[4]。目前,体外分离培养HPLDFs是研究牙周膜组织疾病的重要手段之一。
长链非编码RNA(long non-coding RNA,lncRNA)是一类长度超过200 nt 的非编码RNA[5]。最新测序研究表明,lncRNAs 参与细胞的发育分化、增殖、迁移及凋亡等生物功能[6]。多项研究表明,某些lncRNAs 通过表观遗传、转录前后等方式调控成纤维细胞的增殖、迁移和活化,从而参与多种疾病的发生发展[7-8]。MITCHELL等[9]证实lncRNA MALAT1可参与内调控皮细胞功能和血管生长。lncRNA MALAT1 下调可抑制糖尿病大鼠心肌细胞凋亡,减弱左室功能,抑制视网膜内皮细胞的增殖和迁移,从而减弱视网膜血管损伤的炎症反应[10]。TAO等[11]研究发现,下调lncRNA H19可抑制心脏成纤维细胞增殖,可能与导致DUSP5/ERK1/2 轴相关的部分基因异常表达以及引起细胞周期相关的基因表达上调有关。已有研究证明,过表达lncRNA TUG1与食管鳞状细胞癌的肿瘤分型和分期密切相关[12]。上调lncRNA TUG1 表达可促进骨肉瘤细胞和膀胱癌细胞的增殖[13-14]。因此,lncRNA TUG1 在发育和功能维持方面有重要作用,其表达失调会导致多种疾病。但lncRNA TUG1在HPLDFs 中参与调控牙周组织功能的作用及机制未见报道。
本实验通过研究高表达lncRNA TUG1 对HPLDFs 增殖、迁移及胶原合蛋白成的影响,进一步检测其对TGF-β1/Smads 信号通路蛋白表达影响以探讨相关的分子机制,为lncRNA 调控牙周组织病变、修复及再生过程提供新思路,为进一步治疗牙周病提供理论依据。
1 资料与方法
1.1 资料
1.1.1 标本来源 健康志愿者(12~20 岁)前磨牙或智齿,取自首都医科大学附属北京世纪坛医院、北京积水潭医院及首都医科大学附属北京同仁医院口腔科正畸拔除的牙齿,均无龋坏、牙周炎及根尖周疾病等,患者及其家属均知情同意。本研究经首都医科大学附属北京世纪坛医院伦理委员会批准。
1.1.2 主要试剂 DMEM 培养基及胎牛血清购自美国Life 公司;波形丝蛋白及角蛋白抗体购自Abcam 公司;t-Smad3、p-Smad3 及 TGF-β1 抗体购自Cell Signaling Technology 公 司 ;GAPDH 抗 体 购 自Proteintech 公司;青霉素、链霉素和Ⅰ型胶原酶购自美国Invitrogen 公司;DAPI 染色液购自碧云天生物技术公司;TRIzol 试剂购自美国Thermo Scientific公司;逆转录试剂盒、RT-PCR 试剂盒购自日本TaKaRa 公司;所有PCR 引物均由生工生物公司提供;lncRNA TUG1 过表达慢病毒由上海吉玛制药技术有限公司构建;CCK-8 试剂盒购自日本同仁化学研究所。
1.1.3 主要仪器 ABI 7500 实时定量PCR 仪购自Applied Biosystems 公司;微量移液器购自Eppendorf公司;生物安全柜购自Heal Force 公司;多功能酶标仪购自 Thermo Scientific 公司;SynergyTM2 酶标仪购自美国Bio-Tek公司。
1.2 方法
1.2.1 HPDLCs 的分离培养及鉴定 HPDLCs 采用组织贴壁消化法分离而得。拔取新鲜牙齿标本后迅速置于含双抗预冷的PBS 中,清洗去除血污。将牙齿标本立即转移至实验室超净工作台,于无菌条件下刮除牙根上1/3 牙周膜组织。将牙周膜组织置于无菌离心管,1 200 r/min 离心5 min。弃上清液,加入0.5%Ⅰ型胶原酶500 μl,置于37℃恒温摇床,20~30 min 后加入 500 μl DMEM 完全培养基中和,1 200 r/min 离心5 min。弃上清液,挑取细胞沉淀分别铺于培养皿中,加入少量培养基,微微倾斜置于37℃恒温CO2培养箱中。30 min 后缓慢放平培养皿,继续培养1 周。待细胞长满后使用0.25%胰酶37℃消化3 min,加入完全培养基中和,按1∶3 比例传代。收集第3代细胞培养至细胞融合达60%后进行后续实验。
1.2.2 HPDLCs 的鉴定 采用免疫细胞化学染色方法鉴定HPDLCs标志物。使用0.25%胰酶消化并收集第 3 代细胞,以 1×105个/ml 接种于培养小室,孵育培养过夜。4%多聚甲醛固定10 min,PBS 清洗3次,每次5 min。加入含 0.5%Triton 的 5%BSA 进行细胞穿孔及抗原封闭,室温静置40 min。加入100 μl一抗(波形蛋白 Vimentin,1∶200;角蛋白 Keratin,1∶200),4℃ 孵育过夜,同时以PBS 代替抗体作为阴性对照。PBS 清洗 3 次,每次 5 min。加入 100 μl 二抗,室温孵育45 min,PBS 清洗,梯度乙醇脱水封片。光学显微镜下观察并拍照。
1.2.3 重组lncRNA TUG1 慢病毒感染构建过表达HPDLCs[15]取第 3 代生长对数期的 HPDLCs 接种于6 孔板,细胞分为过表达lncRNA TUG1 慢病毒组(Lv-lncRNA TUG1)、慢病毒空载体组(Lv-NC)和空白对照组(Ctrl)。过表达lncRNA TUG1 慢病毒组、慢病毒空载体组分别加入Lv-lncRNA TUG1、Lv-NC对照空白质粒病毒和培养基转染HPDLCs,空白组不做处理,置于37℃恒温培养箱。培养24 h后,吸弃含病毒培养基,更换2 ml 10%FBS 完全培养基,37℃继续培养;病毒感染72 h后,消化细胞行后续处理。
1.2.4 RT-PCR 检测 各组细胞转染72 h 收获各组细胞 HPDLFs,置于 1.5 ml 无酶 EP 管,1 000 r/min离心5 min;弃上清液,每管加入1 ml Trizol,置于冰上充分裂解后,加200 μl氯仿,室温静置5 min;4℃、12 000 r/min 离心 5 min。取 500 μl 上清液,加入等量异丙醇混匀,-20℃ 静置30 min;4℃、12 000 r/min离心30 min后,可见白色沉淀,弃上清液;加入200 μl 75%乙醇,4℃、12 000 r/min离心5 min;弃上清,室温干燥5~10 min;加入20 μl DEPC 充分溶解。采用全自动酶标仪测定总RNA 纯度,控制A260/A280在1.9~2.1,RNA 电泳检测其完整性。检测浓度及纯度后取2 μg 总RNA 应用逆转录试剂盒合成第一链cDNA。逆转录条件如下:37℃反应15 min,85℃反应5 s。将逆转录所得cDNA 纯化后应用ABI 7500实时定量PCR 仪进行PCR 扩增。反应总体积为20 μl∶1.5 μl cDNA 模板,1 μl 引物,10 μl SYBR®Premix Ex TaqTMⅡ,7.5 μl H2O。PCR 反应条件为:95℃ 预变性30 s,扩增循环95℃ 5 s,60℃ 34 s,共40个循环。以 GAPDH 为参照,采用 2-ΔΔCt法计算 lncRNA TUG1 mRNA 的相对定量表达量;以 TGF-β1 为参照,采用2-ΔΔCt法计算COLⅠ、COLⅢ和COLⅣ mRNA的相对定量表达量。相关引物序列见表1。

表1 相关引物序列Tab.1 Relevant primer sequences
1.2.5 CCK-8 检测细胞增殖 过表达lncRNA 组、空白质粒病毒处理组和空白对照组细胞转染72 h收获各组细胞HPDLFs,分别接种于96孔板,接种密度 3 000 个/孔,37℃ 培养过夜。在 0 h 和 24 h 将10 μl/孔 CCK-8 试剂加入各孔细胞,37℃ 培养 1 h 后测定450 nm处吸光度值。
1.2.6 划痕实验检测细胞迁移 过表达lncRNA组、空白质粒病毒处理组和空白对照组细胞转染72 h收获各组细胞HPDLFs,分别接种于6 孔板,细胞单层融合80%~90%时,换成2%胎牛血清的DMEM 培养基。用200 μl 枪头在各组细胞中垂直划线,无菌PBS 清洗 1 遍,换成 2%FBS 培养基,培养 24 h,在倒置显微镜下观察并拍照。同时,对细胞划痕进行成像和测量,用虚线区域测量细胞迁移面积。
1.2.7 Western blot 检测蛋白表达水平 各组细胞转染72 h 收获各组细胞HPDLFs,用RIPA 裂解液提取各组细胞总蛋白。将提取的蛋白样本与含SDS的loading 缓冲液混合,通过10%SDS-PAGE 胶进行蛋白分离,湿法转移至PVDF膜,5%BSA溶液室温封闭1 h。按照抗体说明书稀释一抗(GAPDH,1∶1 000;t-Smad3、p-Smad3、TGF-β1,1∶1 000),4℃ 孵育过夜。用TBST 清洗3 次,加入二抗,室温下孵育1 h。采用ECL化学发光系统检测信号并拍照分析。
1.3 统计学分析 采用GraphPad Prism 8.0 统计学软件进行统计分析。计量资料以表示,组间比较采用t检验,多组比较采用单因素方差One Way ANOVA分析。P<0.05为差异有统计学意义。
2 结果
2.1 HPDLFs 鉴定 HPDLFs 培养 1 周后可见大量集落生长的细胞,呈纺锤形,有多个突起(图1A)。传代培养至P3 代,细胞形态均匀饱满,提示可能分离培养得到HPDLCs。采用免疫细胞化学染色对细胞免疫表型进行鉴定(图1B、C),与阴性对照组相比,波形蛋白Vimentin 呈阳性,角蛋白Keratin 呈阴性;阴性对照组均未见阳性染色结果(图1D、E)。表明提取的原代HPDLCs 纯度较高,无杂细胞,可用于后续体内外实验。

图1 代表性HPDLFs特性Fig.1 Characteristics of representative HPDLFs
2.2 重组慢病毒感染HPDLFs 中lncRNA TUG1 的表达水平 过表达lncRNA TUG1 慢病毒组(LvlncRNA TUG1)HPDLFs 细胞 lncRNA TUG1 表达水平较空白对照组(Ctrl)或空白质粒病毒处理组(Lv-NC)显著增加(P<0.01),而Ctrl 组或Lv-NC 组HPDLFs 细胞lncRNA TUG1 表达水平差异无统计学意义(P>0.05),见图2。

图2 3组HPDLFs细胞中lncRNA TUG1表达比较Fig.2 Comparison of lncRNA TUG1 expression in HPDLFs of 3 groups
2.3 过表达lncRNA TUG1 对HPDLFs 增殖的影响 Lv-lncRNA TUG1 组HPDLFs 细胞吸光度值在培养12 h、24 h和48 h时均较Ctrl组或Lv-NC组显著增加(P<0.05 或P<0.01),而 Ctrl 组或 Lv-NC 组HPDLFs细胞吸光度值在培养12 h、24 h和48 h时差异无统计学意义(P>0.05),见图3。

图3 不同培养时间点3组HPDLFs细胞增殖能力的比较Fig.3 Comparison of HPDLFs cell proliferation for different incubation time in 3 groups
2.4 过表达lncRNA TUG1 对HPDLFs 迁移的影响 Lv-lncRNA TUG1 组HPDLFs 细胞划痕面积均较 Ctrl 组或 Lv-NC 组显著减少(P<0.01),而 Ctrl 组或Lv-NC 组HPDLFs 细胞划痕面积差异无统计学意义(P>0.05),见图4。
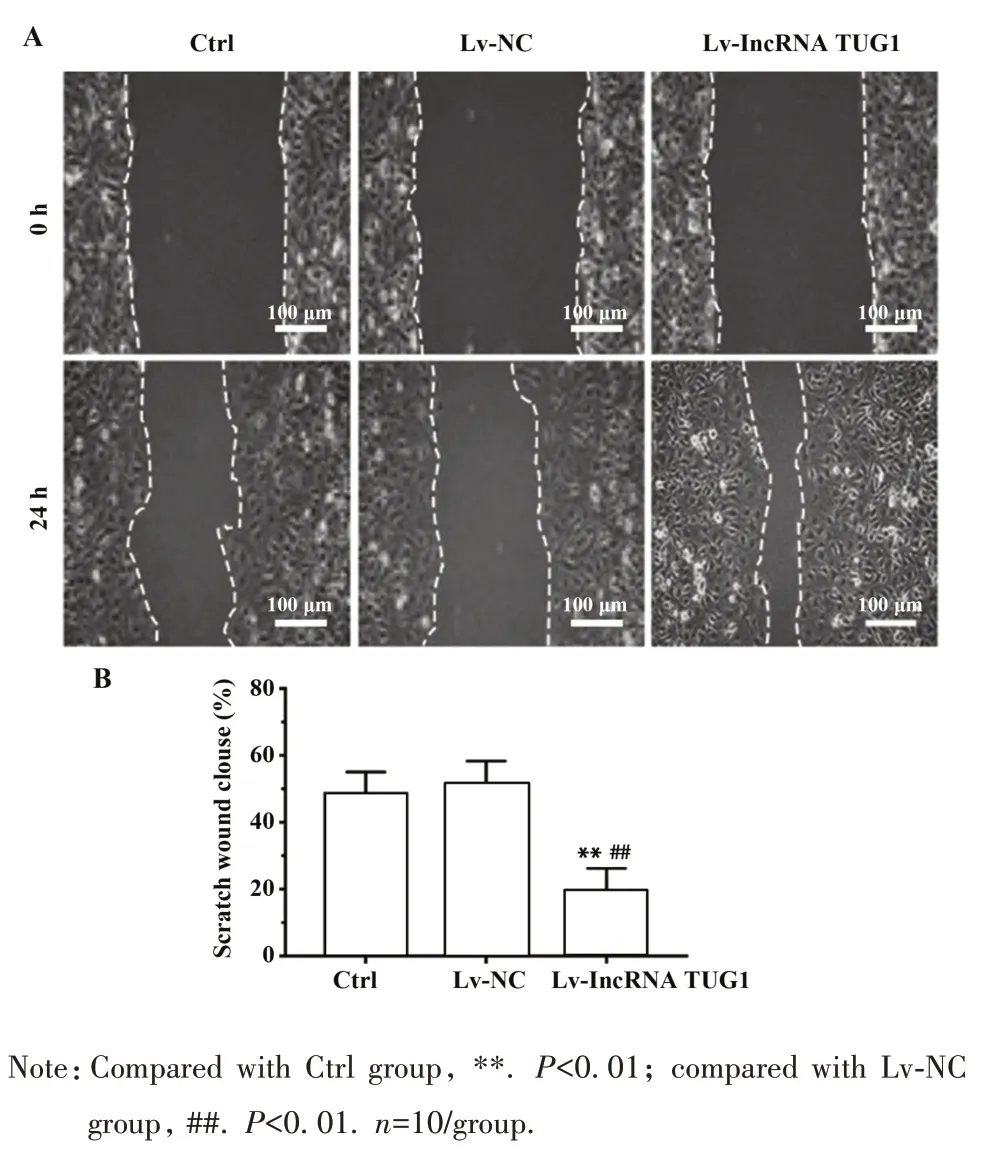
图4 3 组HPDLFs 细胞迁移情况(A)和细胞划痕面积(B)比较Fig.4 Comparison of HPDLFs cell migration condition(A)and cell healing area(B)in 3 groups
2.5 lncRNA TUG1 对 HPDLFs 纤维钙化的影响Lv-lncRNA TUG1 组 HPDLFs 细胞中 COLⅠ、COLⅢ、COLⅣ及MMP-2 mRNA表达水平均较Ctrl组或Lv-NC组显著增加(P<0.05 或P<0.01),而Ctrl 组或Lv-NC组 HPDLFs 细胞中 COLⅠ、COLⅢ、COLⅣ及 MMP-2 mRNA表达水平差异无统计学意义(P>0.05),见图5。

图5 3组HPDLFs细胞中COLⅠ、COLⅢ、COLⅣ及MMP-2相对表达量比较Fig.5 Comparison of relative expressions of COLⅠ,COLⅢ,COLⅣand MMP-2 in HPDLFs of 3 groups
2.6 lncRNA TUG1 在HPDLFs 细胞表型转化中的分子机制研究 Lv-lncRNA TUG1 组HPDLFs 细胞中 TGF-β1 蛋白表达水平均较 Ctrl 组或 Lv-NC 组显著增加(P<0.05 或P<0.01),而 Ctrl 组或 Lv-NC 组HPDLFs 细胞中TGF-β1 蛋白表达水平差异无统计学意义(P>0.05),见图 6A。Lv-lncRNA TUG1 组HPDLFs 细胞中p-Smad3 蛋白表达水平均较Ctrl 组或 Lv-NC 组显著增加,而 Ctrl 组或 Lv-NC 组 HPDLFs细胞中p-Smad3 表达水平差异无统计学意义。3 组HPDLFs 细胞中t-Smad3 表达水平差异无统计学意义(P>0.05),见图6B。

图6 3 组 HPDLCs 中 TGF-β1 和 Smad3 蛋白的表达情况比较Fig.6 Comparison of expressions of TGF-β1 and Smad3 proteins in HPDLFs of 3 groups
3 讨论
牙周炎在世界范围内的发病率约为十万分之一,其高复发率、长期疗效不佳易导致牙齿周围槽骨逐渐丧失,严重影响患者生活质量[16-17]。遗传因素是牙周组织发育及炎症发生发展的关键因素,因此,寻找新的标志物以开发更有效、安全的治疗策略具有重要临床意义[18]。HPLDFs 与牙周组织病变、修复及再生过程密切相关,能持续合成与降解胶原蛋白纤维以维持牙周组织的更新与重建,且HPLDFs 还能分泌多种细胞活性因子刺激成骨细胞和破骨细胞完成牙周组织的修复与再生,如骨形态发生蛋白家族BMP-2、BMP-4及BMP-7等[19]。因此,HPLDFs 异常活化也是牙周炎发病的主要决定因素之一。
研究表明,lncRNA 失调可导致多种包括炎症性疾病、纤维化和肿瘤等相关疾病[20-21]。近年来,lncRNA 参与多种纤维化疾病发生的研究日益受到关 注[22]。lncRNA TUG1 是 一个 长 度为 7.1 kb 的lncRNA,首次在小鼠视网膜细胞发育的基因组筛检中被发现[23]。近期研究表明,lncRNA TUG1 是一种新型癌基因,其表达上调可促进肿瘤细胞的增殖和迁移如lncRNA TUG1 表达下调可抑制肾细胞癌细胞的迁移、侵袭和增殖,并激活细胞凋亡过程[14]。最新研究发现,非小细胞肺癌中lncRNA TUG1 表达下调[24]。研究证实,lncRNA 对 HPDLFs 的影响主要为促进细胞增殖、胶原合成、成骨分化等[25]。然而,lncRNA TUG1 在HPDLFs 中的潜在功能尚不清楚。本研究发现,过表达lncRNA TUG1的HPDLFs增殖、迁移均显著增加,提示lncRNA TUG1 是治疗牙周病的新靶点,其表达上调可促进HPDLFs的活化。
TGF-β1 参与牙齿移动、牙周组织发育、损伤修复等多种生物学过程,可显著促进体外培养的HPDLFs 增殖和 ALP 活性[25-27]。体外研究表明,TGF-β1在早期可诱导Ⅰ型胶原mRNA 表达,随着时间延长,胶原的表达反而受到抑制[28]。但是,对于lncRNA在HPDLFs 纤维钙化方面的信号转导通路的研究国内外鲜见报道。本研究初步探讨了lncRNA TUG1对HPDLFs 增殖、胶原合成、纤维钙化等作用可能涉及的信号通路。与对照组相比,lncRNA TUG1 可上调 TGF-β1 和 p-Smad3 信号蛋白的表达量,提示lncRNA TUG1 可能通过激活 TGF-β1/Smads 途径促HPDLFs的异常活化。本研究还表明,lncRNA TUG1是治疗牙周病的新靶点,提示其可通过上调TGF-β1/Smads信号通路促进HPDLFs的增殖、迁移能力及纤维化。
综上所述,lncRNA TUG1 是治疗牙周病的新靶点,lncRNA TUG1 表达上调可显著促进HPLDFs 的增殖、迁移及促进COLⅠ、COLⅢ、COLⅣ和MMP-2的 mRNA 水平,同时上调 TGF-β1/p-Smad3 信号通路蛋白,提示 lncRNA TUG1 通过激活 TGF-β 途径促HPLDFs 增殖、迁移及纤维化过程,但其具体调控机制有待进一步探索。
猜你喜欢
锦州医科大学报(2022年2期)2022-05-07
皮革科学与工程(2022年1期)2022-01-15
食品安全导刊(2020年21期)2020-12-03
食品安全导刊(2020年18期)2020-12-03
数理医药学杂志(2020年10期)2020-10-17
湖南农业科学(2020年1期)2020-04-18
中国现代医生(2015年8期)2015-04-17
中国科技纵横(2014年14期)2014-08-27
食品工业科技(2014年13期)2014-03-11
体育科研(2013年2期)2013-05-31
