
渡鸦鸣叫所隐藏的神经状态解析*
2022-02-15 07:08杨利琼刘昉昉
实验动物科学 2022年6期
杨利琼 徐 帆 刘昉昉
(1.成都医学院药学院,成都 610500)(2.成都医学院公共卫生学院,成都 610500)(3.西南民族大学,成都 610041)
在动物界中,声音交流无处不在。为了发现人类语言和语音之间的异同[1],许多研究都聚焦于动物声音的交流上。有研究调查了自由生活的几内亚狒狒(Papiopapio)的声音库,并检查了动物声音信号的结构和用法。结果表明,灵长类动物的声音结构在很大程度上受该物种的社会系统的影响,而且它们可以更灵活地调整发声功能,从而反映它们的社交互动[2]。有研究古氏土拨鼠(Cynomysgunnisoni)警报鸣叫的结果显示,它们对地面掠食者发出多音符警报鸣叫,其掠食者的声学结构根据所引起的刺激而变化。这表明有关刺激大小和形状的信息是编码于土拨鼠的警报鸣叫中[3]。更有趣的是,有实验验证了黑松鸡可以识别乌鸦的警报鸣叫。它们显示出典型的颈部伸展状态,然后对乌鸦进行头部扫视,最终逃脱[4]。一些研究人员认为这些声音是基于情感而产生的,反映了声音发出者内心的情绪状态[5-6]。例如,Conner等从弗吉尼亚州西南部的渡鸦(Corvuscorax)发出的1 210个声音片段中,推测不同类型的鸣叫与兴奋、激烈的互动、攻击、警觉、求爱、服从和沮丧有关[7]。
大脑中的神经回路在有节奏的运动或复杂的自发动作等环境刺激中起着主导作用[8]。神经状态是指自主神经系统的活动受到中枢神经元的控制,这些中枢神经元对广泛的传入的信息作出反应[9]。在对传入信息进行整合后,自主神经的传入活动经过调节,从而根据主要器官系统作出适当的反应[10]。鸟类的鸣叫依赖于发育完善的鸣管并接受各级发声中枢组成的机能控制系统的调控[11]。鸣禽发声学习的控制系统主要由一条直接神经通路和一条辅助神经通路组成[12]。鸟类鸣叫富含神经生物学意义,比如鸟类的情绪、认知功能和对外界事物的反应状态。这些声学信号的特征有助于推断系统发育和进化关系[13]。从神经学的角度来看,有4个主要的分类,包括警报、飞行、乞求和歌唱[14]。首先,异种警报鸣叫在不同种类的鸟中尤为常见。在2018年,有研究测试了自由放养的渡鸦(Corvuscorax)对特定警报鸣叫的响应,并比较了它们对寒鸦(Corvusmonedula)异种警报鸣叫的响应。结果显示,渡鸦意识到了寒鸦警报鸣叫的含义,但渡鸦只有在受到高掠夺威胁的情况下,才会做出反应[15]。其次,鸟类通常在飞行过程中产生飞行鸣叫。2016年,实验证明橙尾鸲莺(Setophagaruticilla)的飞行鸣叫可能具有性别和个体特征,这是鸟类飞行鸣叫具有个体性或性别二态性的第一个证据,而飞行鸣叫可能比简单群体联系和凝聚的功能更具体[16]。除此之外,成年乌鸦经常分享歌曲。它在社会学习和社会团员的融合中起着重要作用。群体歌曲与特定的社交互动密切相关,例如减少攻击和增进群体成员之间的关系[17]。
乌鸦表现有较强的智力和社会性活动。在过去的十年中,它已被用于许多特定的思维和大脑研究中。例如,乌鸦被用于记忆研究中,首先,它们能够预测未来事件[18];其次,可以使用工具以获得奖励[19],并且识别物体的密度[20]。除此之外,乌鸦在觅食[21]、保卫自己的领土[22]、发情、交配[23]和养育子女[24]时经常发出鸣叫来表达情绪。尽管我们可以听到各种不同的声音,但是常常缺乏背景或者行为信息,而难以对鸣叫背后的生物学意义进一步探究。
早在20世纪40年代初期,McCulloch 和Pitts提出了M-P模型的概念,该模型提供了神经元的基本模型及其工作模式[25]。此后,Widrow和Hoff提出了基于多层感知器的最小均方算法,有效地提高了神经网络权重优化的速度和准确度[26]。目前,用于鸟类鸣叫分类的算法主要有支持向量机(support vector machine,SVM),这是基于核的算法,支持向量机算法根据区域中的样本计算该区域的决策曲面,由此确定该区域中未知样本的类别。该算法的泛化能力比较强,但是对核函数的高维映射解释力不强,对缺失数据很敏感[27-28]。其次,朴素贝叶斯算法(naive bayesian algorithm,NBA)是一种利用概率统计进行分类的算法。这种算法基于贝叶斯定理来预测未知类别的样本属于各类别的可能性。由于贝叶斯定理的成立需要条件独立性假设前提,而此假设在实际情况中经常是不成立的,因而其分类准确性就会下降[27]。而本实验运用的反向传播神经网络(back propagation neural network,BPNN)算法,分类的准确度高、并行分布处理能力强、分布存储及学习能力强而且具备联想记忆的功能。
在这里,我们假设4个来自渡鸦行为标记的鸣叫具有不同的声学参数,表明渡鸦的神经状态。由此,我们使用了BPNN来识别渡鸦鸣叫背后的神经生物学意义,并对其进行分类。这项研究可能有助于我们更好地了解鸟类鸣叫的神经生物学意义,这不仅局限于同一物种中,还可以跨物种研究。
1 材料和方法
1.1 研究对象
渡鸦(Corvuscorax)是一种全身黑色的大型雀形目鸦属鸟类,体型较乌鸦大。渡鸦和乌鸦近缘,同属于鸦科。渡鸦的喙比乌鸦厚得多,羽衣更为蓬松,尤其是喉部的羽毛。渡鸦的羽毛光亮,还带有蓝色或紫色光辉。主要分布在整个全北区。渡鸦通常独栖,也可聚成小群活动觅食。主要是吃腐肉及连带的蛆及埋葬虫科[29]。
1.2 数据来源
从xeno-canto鸟类声音数据库中(https://www.xeno-canto.org/species/Corvus-corax)下载了851个渡鸦的声音样本,并将它们的详细信息整理到Excel文件中,包括了以下11个变量:通用名称、学名、记录持续时间、录制人、记录时间、国家、位置、高度、鸣叫类型、行为标志和音频文件分类号。
1.3 行为类别及其发声
由行为分类定义的变量类型,包括警报、飞行、乞求和歌唱,记录了渡鸦的具体行为。
1.4 数据清理
为了减少噪音影响并增强分类效果,排除音频中含两个以上渡鸦鸣叫的样本。选择了4种不同类别的声音样本(警报、飞行、乞求和歌唱)。因此,最终数据分析中仅包含100个声音样本。
1.5 声学参数提取
使用MATLAB脚本提取以下声学参数[30-31]:(1)平均基频(mean fundamental frequency,平均f0)(Hz);(2)最大f0(Hz);(3)最小f0(Hz);(4)f0范围(Hz);(5)起始f0(Hz);(6)终止f0(Hz);(7)中值f0(鸣叫一半时间的f0)(Hz);(8)斜率S-M(从起始f0到最大f0的斜率;Hz/s);(9)斜率M-E(从最大f0到终止f0的斜率;Hz/s);(10)音调变化率(频率变化次数/s);(11)谐噪比(Harmonicity,HNR)(dB);(12)基频微扰(连续f0测量值的绝对差/平均周期);(13)f0变异(所有f0变化总和/声音持续时间);(14)最小幅度范围(dB)。
1.6 统计学分析
构建典型的三层BP网络的拓扑结构,它包含输入、隐藏和输出层,参见图1。这三层完全互连,而隐藏层可以单个或多个存在。在层之间有两种信号:工作信号(实线)和误差信号(虚线)。在施加输入信号后,工作信号会向前传播,直到输出产生实际输出为止。而误差信号(虚线)由网络的实际输出与预计输出之间的差异表示。它从输出端逐层传播,以调整每一层的权重和阈值,直到结果到达输入层。

图1 BP 神经网络结构
2 结果
2.1 渡鸦鸣叫声学参数的提取
我们选择了4种不同的渡鸦鸣叫类别,包括警报、乞求、飞行和歌唱。表1显示了从MATLAB中提取的这4种渡鸦鸣叫的声学参数。由表1可以看出,在参数振幅范围中,飞行鸣叫的范围最大,警报鸣叫范围最小。然而在参数音调变化率中,警报鸣叫的变化最大,飞行鸣叫的变化最小。在参数起始f0中,警报鸣叫的值最大,歌唱鸣叫的值最小。特别在声音的斜率M-E中,渡鸦的乞求鸣叫最大,飞行鸣叫的斜率M-E最小,这在4组之间具有统计学差异(P<0.05)。

表1 渡鸦4种鸣叫声学参数的单因素方差结果
2.2 BPNN对渡鸦鸣叫的分类
使用BPNN方法对渡鸦鸣叫的生物学意义进行分类。首先,选择使用14个声音参数作为训练样本。测试前100个声音样本,其中包括从训练数据集中导入的所有数据。其次,对数据进行预处理:由于现实的复杂性,使用偏差归一化的标准过程[(x.Std.=(x-min)/(max-min)]来执行改进的神经网络方法并减少整体的负面影响,新数据是通过无量纲处理构建的。第三,我们进行了初步参数的选择。
典型的拓扑结构,其中输入层中有14个神经元,输出层中有一个神经元。此外,如图2,隐藏层中有12个神经元。BPNN构造的唯一目的是对数据进行分类。BPNN构造使用了双传递S形函数和线性函数,结构在输入层-隐藏层中使用“tansig”函数,在隐藏层-输入层中使用线性函数表示。此外,使用“算法”方法(梯度下降法与牛顿法相结合)进行演算,并在达到最佳效果时终止。详细的演算执行过程如图3所示。
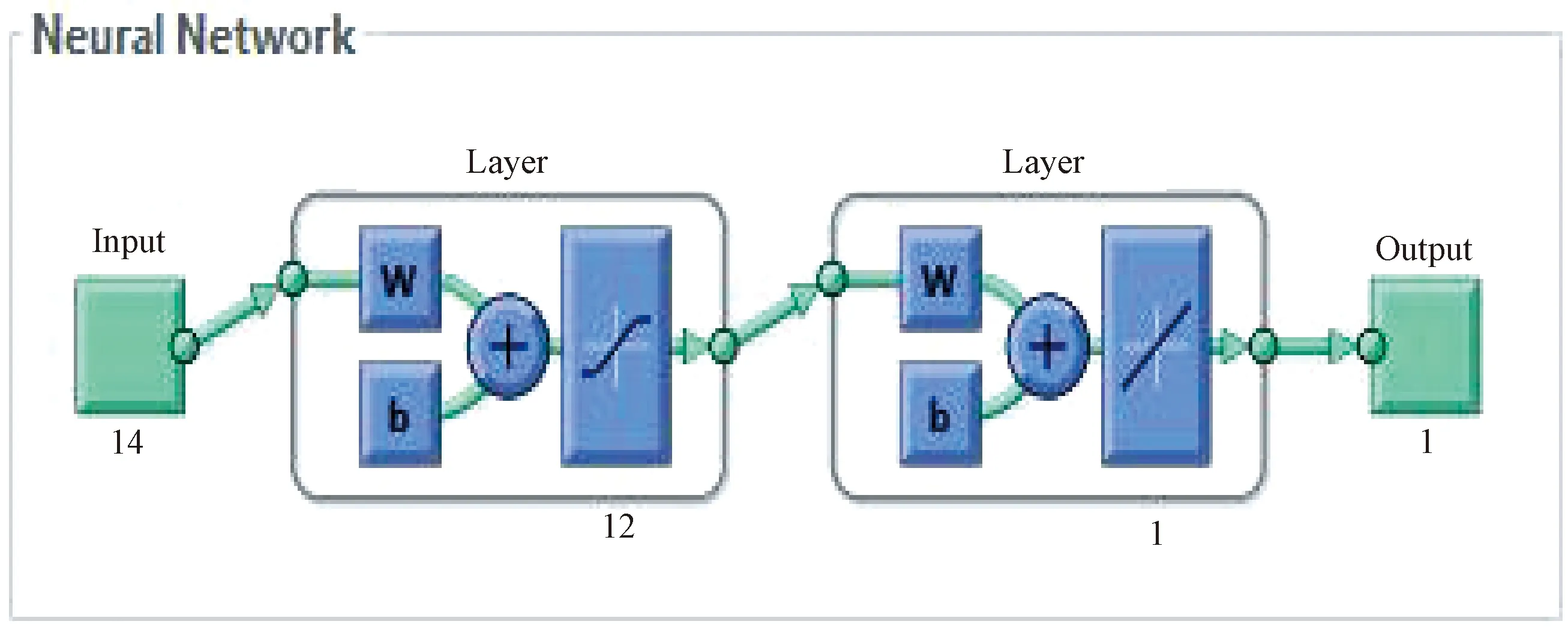
图2 神经网络模式

图3 演算过程
为了在0.3秒内接近集合,一共反复执行了367次(367次实际演算),见图 3。演算误差性能小于设定的目标误差。演算结束时,误差面梯度达到预设值。确认检验值为6,当样品的误差曲线连续6次反复均未下降时,结束演算。
2.3 预测模型检验
验证预测效果:利用演算网络预测渡鸦叫声的类型,以检测网络的预测和提升能力。如图4所示,该模型模拟了一条曲线来拟合源数据点,其中蓝线表示声音类型,红星表示真实的声音类型。
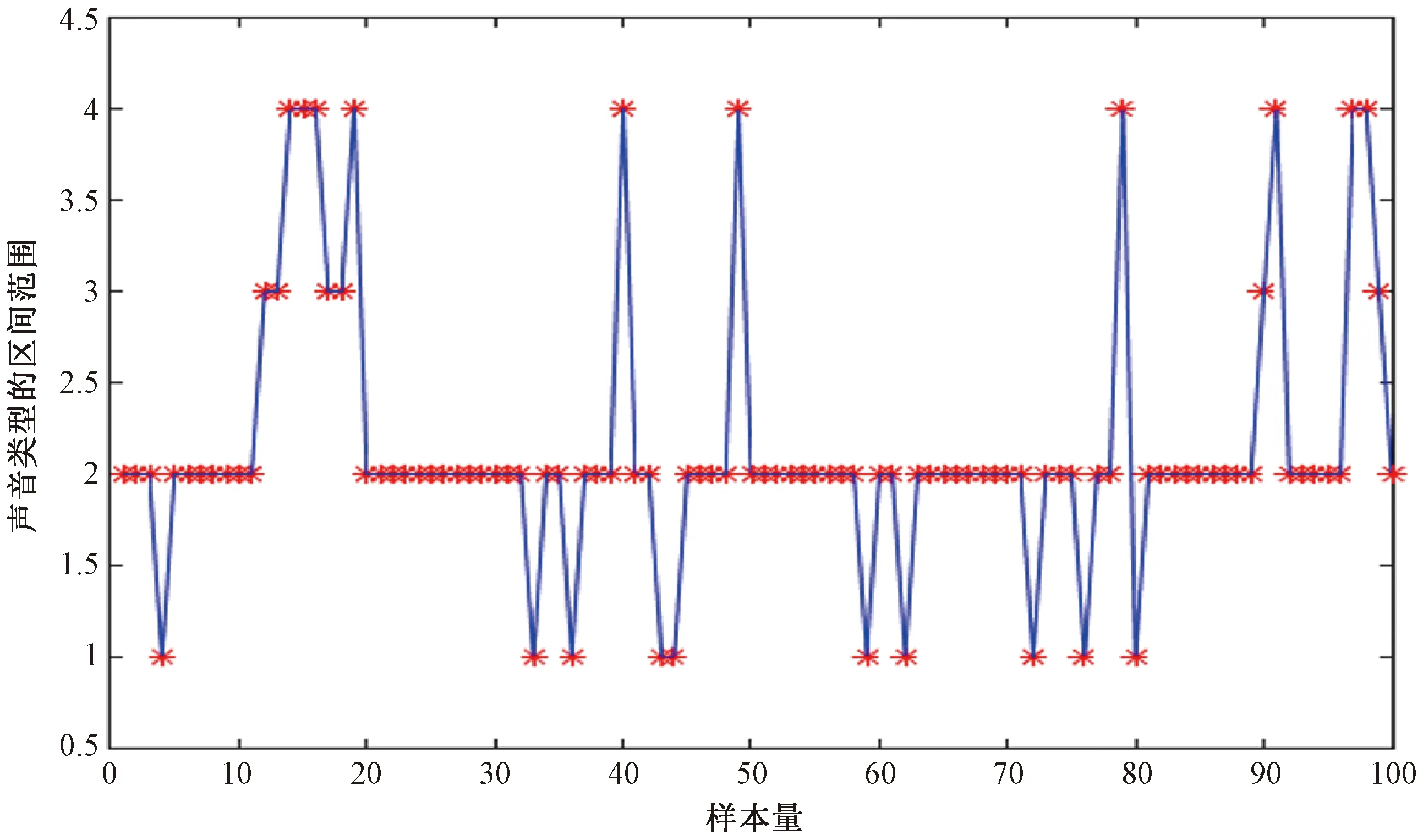
图4 预测效果
由图4可以看出,预测值的总体趋势与实际值较为接近。从而推断出,模型的预测在某个参考值上是准确的。结果表明,实际分类和预测分类的平均绝对误差和平均相对误差分别为1.0×10-9和1.0×10-10。
3 讨论
根据渡鸦的行为描述,展示了4种渡鸦鸣叫类型:警报、乞求、飞行和歌唱。通过使用BPNN方法,成功地对这些行为进行了分类。BPNN具有出色的非线性接近能力、泛化能力和易用性,主要用于分类和预测。该方法在图像处理、信息处理、智能控制、故障检查、企业管理和市场分析等方面均取得了显著的成果[32-33]。例如,使用BP神经网络进行肺声识别,其中以肺声数据为输入量,以肺声类别为输出,识别成功率为92.5%[34]。使用BPNN神经网络通过声音质量预测异常车辆[35]。
动物的声音交流是将信息从一个动物转移到另一个动物的有效方法。例如,鸟类的警报鸣叫是响应危险信息而发送的信号。有研究记录了白尾雷鸟(Lagopusleucurus)对自然发生和模拟捕食者警报鸣叫的反应。结果表明,80%的幼鸟对警报蜂鸣声有反应[36]。在动物的交流方面,声音的产生不仅取决于多种器官协调的过程[37-39],还取决于中枢神经系统的调节[40]。在漫长的进化过程中,动物通过构建一系列的条件反射,成功适应了环境[41]。乌鸦被认为是最聪明的鸟类[42-43]。它已经被用于研究大脑思维、工具的使用、意识、学习和记忆。有研究表明,乌鸦鸣叫可以区分它们的性别和年龄[44],并指出乌鸦可以通过它们的记忆预测未来[18]。
声音不仅包含了生物体自身器官的振动信息,还具有潜在的神经状态的信息。因此,区分声音背后的神经状态具有重要意义。我们可以通过了解渡鸦声音背后的神经状态信息,从而更好的了解渡鸦鸣叫的神经生物学意义。例如,警报鸣叫通常在危险事件临近时发生,这种鸣叫用于避免有害事件并生存[45-46]。飞行鸣叫发生在战斗中,失败的一方会屈服于占优势的一方,以避免对其身体和社会地位造成进一步伤害[47-48]。此外,当鸟群为生活来源而乞求时,就会发生乞求鸣叫[49-50]。另外,在动物愉快的交流或寻求配偶时,可能会发生歌唱。但是,具体动机仍存在争议。这种人工网络可以帮助区分不同渡鸦声音的行为类型。BPNN区分的结果可以加强我们对这些声音的生物学和进化意义的理解。
但是,本研究的渡鸦鸣叫样本来源于鸟类数据库,其分类依据数据库上鸟类学家的分类,同时筛选后4种鸣叫的样本量存在分布比例不均衡。因此,其鸣叫的行为意义分类和数据可靠性还需声音回放性实验验证。
综上所述,我们认为BPNN方法是一种很有前途的渡鸦鸣叫分类的候选方法。此外,此方法有助于我们进一步研究渡鸦的神经生物学。
猜你喜欢
学与玩(2022年9期)2022-10-31
卫星应用(2022年3期)2022-05-23
文苑(2020年12期)2020-04-13
科普童话·百科探秘(2019年8期)2019-09-18
小太阳画报(2019年1期)2019-06-11
小天使·一年级语数英综合(2018年5期)2018-06-22
少儿科学周刊·少年版(2018年12期)2018-01-26
小学生必读(低年级版)(2017年5期)2017-08-12
Coco薇(2017年8期)2017-08-03
小学阅读指南·低年级版(2017年2期)2017-03-23
