
中国南瓜×印度南瓜种间杂交自交系及组合的遗传多样性分析
2022-02-15 02:55古斌权张香琴
华北农学报 2022年6期
宋 慧,古斌权,张香琴
(宁波市农业科学研究院 蔬菜研究所,宁波市瓜菜育种重点实验室,浙江 宁波 315040)
利用表型和分子标记技术分析生物遗传多样性,是作物育种的重要环节[1-2],是资源鉴定和品种亲缘关系评价的重要手段[3-4]。方法成熟,报道较多[5-9],主要开展自然群体关联分析[10]、作物品种指纹图谱构建[11]和地方种质资源多样性分析[12-13]等研究。但这类报道多停留在为育种实践提供理论依据的层面,对进一步通过引种、种间杂交或诱变等育种手段,拓宽作物遗传多样性的研究较少;进而利用表型和分子标记技术,比较和分析育种后代与现有资源的遗传多样性的报道就更少。
目前,市场畅销的食用南瓜类型主要有中国南瓜的蜜本南瓜、香芋南瓜类型,印度南瓜的贝贝南瓜、板栗南瓜、贵族南瓜类型,以及美洲南瓜的长条笋瓜、圆形笋瓜和小菊瓜等。近年来,相同类型的南瓜品种,商品名称更换很快,但是同质性很强[14],推陈出新的后劲不足。有学者从南瓜商品种鉴定分类的角度,对各地搜集的商品种进行聚类分析,结果发现,中国南瓜、印度南瓜和美洲南瓜3个种之间的差异大,种内的遗传相似度高[15],相似系数为0.830~1.000[16-17]。地域来源相同的商品种通常会聚为一类[18-19],但是张天明等[20]研究却发现,由于当今种子市场繁荣,各地区相互引种育种,导致同质遗传型品种再组合,品种(系)间遗传多态性降低,地域差异趋同[14,16]。周琳等[21]曾利用SSR分子标记对路易斯安那鸢尾品种、杂交后代和诱变材料进行遗传多样性分析,结果表明,诱变材料与亲本遗传差异大。种间杂交也是拓宽南瓜遗传多样性的有效手段[22-25]。为此本研究院前期通过中国南瓜(Cucurbitamoschata)×印度南瓜(Cucurbitamaxima)种间远缘杂交和胚胎拯救技术,获得种间杂种[26],并经过回交和多代自交,获得耐热的自交系[27]。现通过与骨干亲本配制杂交组合,选育南瓜新品种。
为了进一步揭示种间杂种与现有南瓜商品种和核心亲本具有明显差别,本研究对目前市售的食用南瓜商品种、本研究院长期积累的骨干亲本、以及中印南瓜种间杂交自交系及其新配组合,进行遗传多样性分析。系统评价现有食用南瓜品种和育种材料的亲缘关系,明确中印南瓜种间杂种自交系及其新配组合的聚类地位,在表型和分子层面分析南瓜种间杂种拓宽南瓜遗传背景的可能性。此外,有关南瓜植物学性状调查的报道较多,从种子形态、果实形态以及植株长势等多方面,调查指标多达70余项[28]。繁琐的调查条目,不利于育种材料的高效调查与分析。为此,本研究拟利用主成分分析方法,解析30个表型性状的主成分构成,简化食用南瓜评价表型指标。
1 材料和方法
1.1 植物材料
供试材料53份,包括32份印度南瓜、5份中国南瓜和1份美洲南瓜,以及15份本研究院前期获得的中印南瓜远缘杂交自交系及自配杂交组合(表1)。其中商品种10份,农家种3份,骨干亲本25份,自配组合15份。商品种涵盖市场畅销的各种食用南瓜,涉及中国南瓜的蜜本南瓜香芋南瓜类型,印度南瓜的贝贝南瓜、板栗南瓜、贵族南瓜类型,以及美洲南瓜的小菊瓜。试验在宁波市农业科学研究院实验基地和瓜菜育种重点实验室进行。每份材料定植15株,采用随机区组设计,重复3次,常规栽培管理。
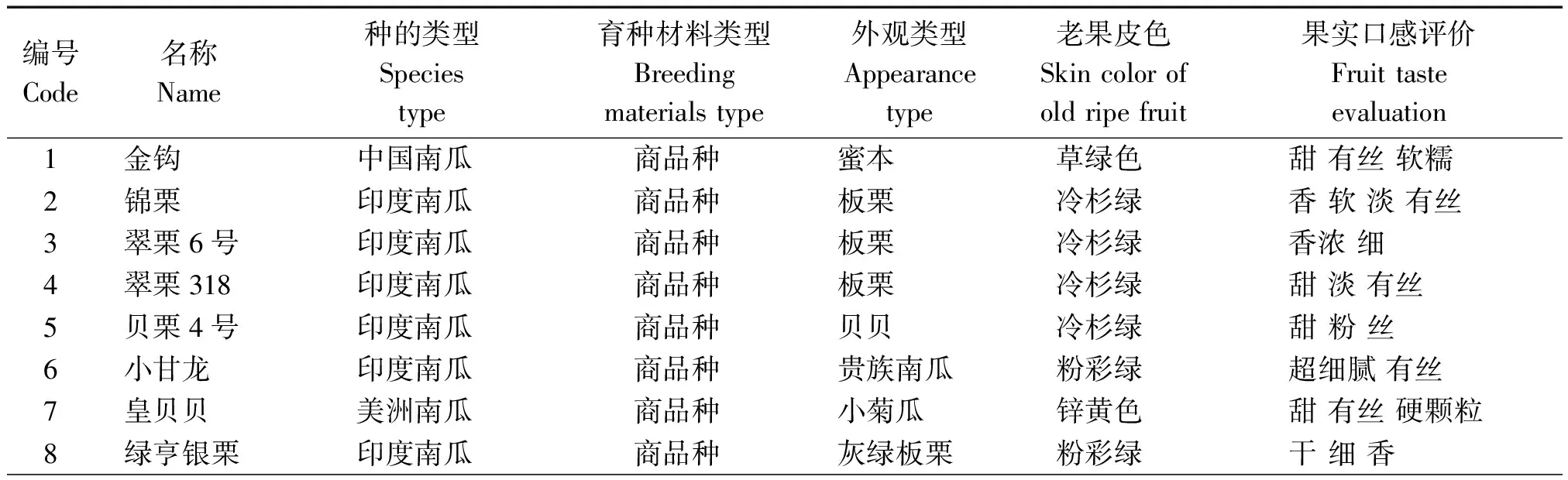
表1 供试南瓜名称、类型和部分表型性状Tab.1 The name,type and partial morphological traits of pumpkins tested
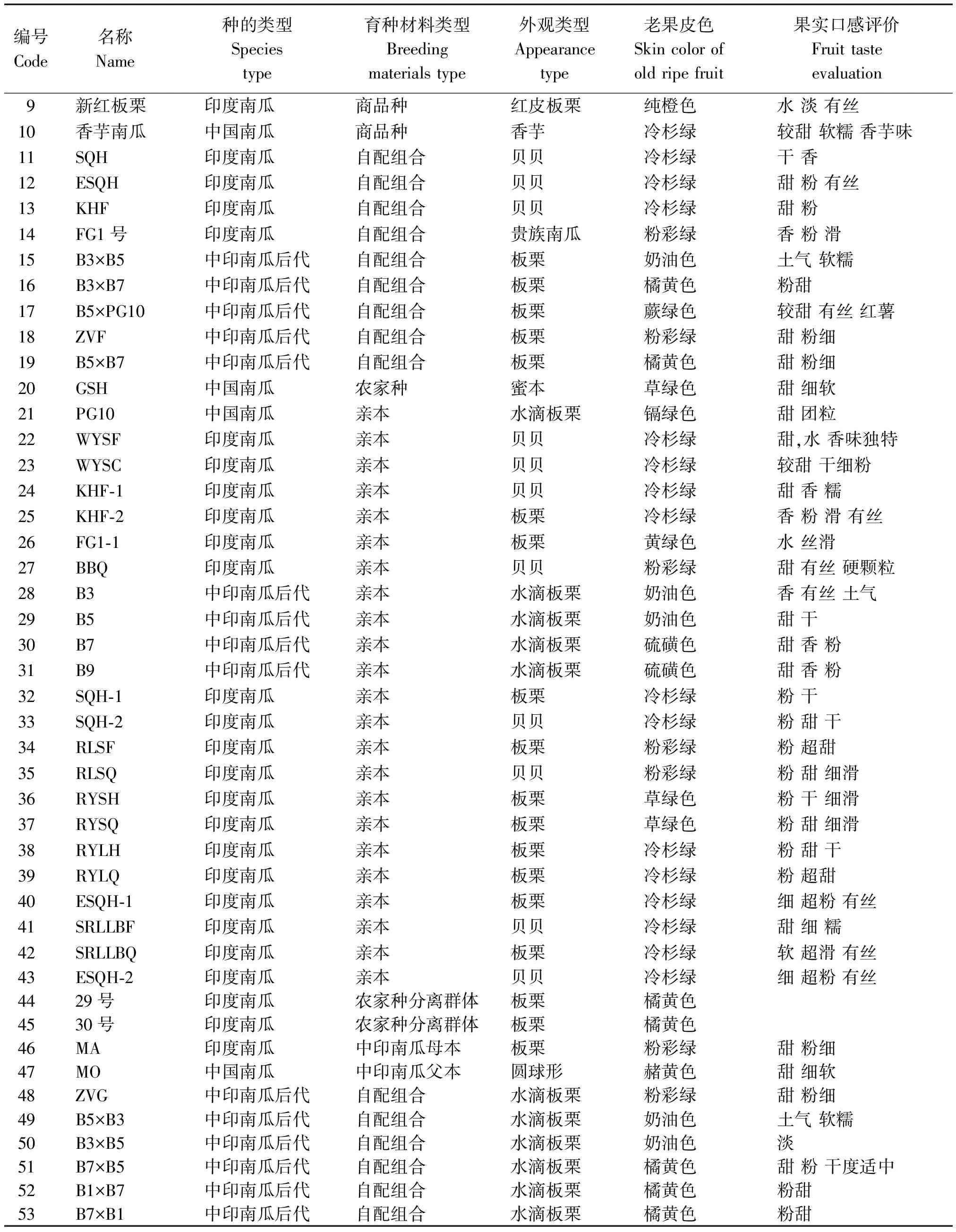
表1(续)
1.2 试验方法
1.2.1 植物学性状调查 参照蔡宝炎[28]和褚盼盼[29]制定的南瓜植物学性状调查表,结合本试验制定南瓜形态学性状调查内容和赋值标准(表2)。分别在植株伸蔓期、开花期、结果期和采收期,调查植株和果实的30个表型性状。需要特别说明调查时期和记载标准的表型性状见表3,其中,果实口感评价,按照尹玲等[30]报道的南瓜感官评价方法进行。
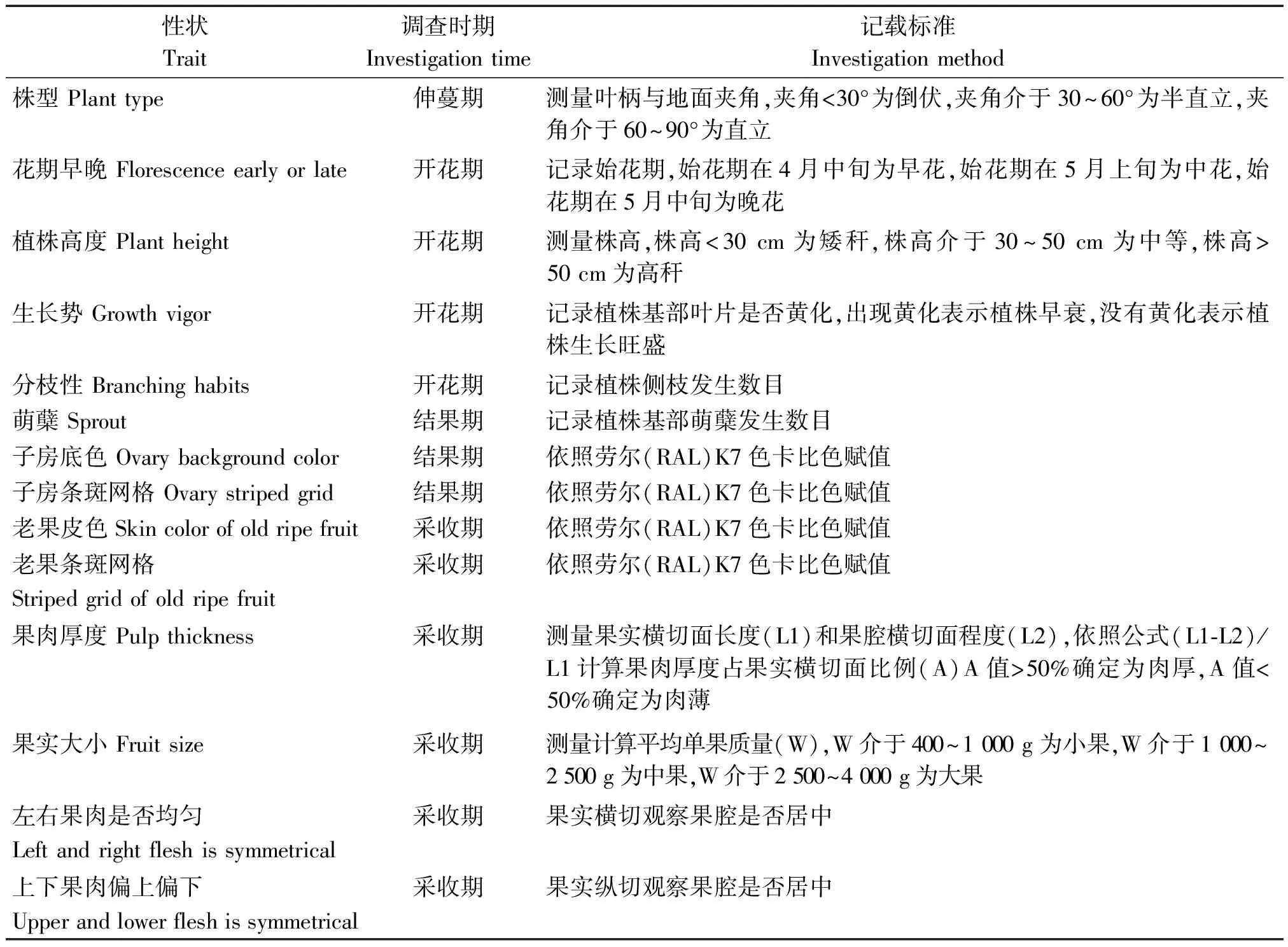
表3 部分南瓜形态学性状调查时期和记载标准Tab.3 Investigation time and method of partial pumpkins morphological traits
1.2.2 遗传多样性分析 利用孔凡洲等[31]通过Excel建立的生物多样性指数计算软件,计算30个统计性状的Berger-Parker指数(d)、Margalef指数(dMa)、和Shannon指数(He′)。其中Berger-Parker指数用来表示遗传多样性的优势度,Margalef 指数用来表示丰富度,Shannon 指数用来测定群落多样性的高低。3个指数数值越大,表明群体多样性越丰富。
1.2.3 表型聚类和主成分分析 利用SPSS 19.0软件[23],采用聚类功能,各样本间的距离用卡方距离进行度量,对30组表型数据进行聚类。利用SPSS 19.0软件,对53份南瓜材料的30个统计性状进行主成分分析,根据累计贡献率≥85%原则,筛选主成分因子。在每个主成分因子中,30个统计性状的特征向量排序不同,每个因子中特征性向量绝对值较大的1~2个性状,是该主成分因子代表的主要特征。利用这些主成分因子对53份南瓜材料进行二次表型聚类分析。如果二次表型聚类与分子聚类具有较高符合度,则这些主成分因子可作为简化表型性状统计的指标。
1.2.4 SSR分子标记扩增与聚类图绘制 在植株伸蔓期,取茎尖幼叶,液氮储运。利用EasyPure®Plant Genomic DNA Kit试剂盒(北京全式金生物技术有限公司)提取样品DNA。利用K5600超微量分光光度计(北京凯奥科技发展公司)和0.8%琼脂糖凝胶电泳检测DNA质量和浓度。参照Gong等[32]报道,委托生工生物工程(上海)股份有限公司合成南瓜SSR引物。选择在前期南瓜SSR标记扩增试验中,带型清晰、多态性稳定的引物23对,进行标记扩增。
参照宋慧等[33]的报道,采用康为世纪生物科技有限公司2×Taq MasterMix(Day)PCR混合液,按照说明书调整反应体系为10 μL:5 μL 2×Taq MasterMix(Day)、0.4 μL 引物(10 mmol/L)、1 μL DNA(10 μg/μL),加ddH2O至10 μL。PCR扩增程序为:94 ℃ 2 min;94 ℃ 30 s,60 ℃ 30 s,72 ℃ 10 s,35个循环;72 ℃ 2 min。PCR产物使用5% Yeasen高分辨率琼脂糖凝胶(上海翊圣生物科技有限公司)电泳。上样量为1.1 μL,电压90 V,电流100 mA,时间3 h。凝胶成像系统观察扩增产物。
根据琼脂糖凝胶电泳结果,统计条带清晰、样品间有多态性的位点,相同迁移位点扩增出条带的记为“1”,无条带记为“0”,建立0和1矩阵。计算标记多态信息量,多态性比例=(多态性位点/扩增位点)×100%。利用NTSYS-pc2.10e软件处理数据,绘制聚类图。
2 结果与分析
2.1 表型结果
2.1.1 南瓜形态学性状的遗传多样性 图1展示了3种南瓜测试组合30个形态学性状的3个遗传多样性指数,结果显示,3种南瓜组合分别为10个南瓜商品种、10个商品种+28份骨干亲本以及10个商品种+28份骨干亲本+15份中印南瓜杂交种后代。结果显示,随着育种材料的加入,供试南瓜的3个遗传多样性指数均增加。He′的增幅较小(均值2.09~3.72),d和dMa的增幅明显(均值6.84~31.42和2.77~9.92)。结果表明,供试10份南瓜商品种之间的表型已经存在不同之处,商品种的选择具有代表性;骨干亲本和中印南瓜杂交种后代的加入,在株型、生长势和分枝分蘖等生长习性,以及果实皮色、果型等多方面使得商品种群体多态性丰富度提高。由于增加的材料均以选育食用南瓜为目标,因此,在果实口感、果肉厚度等方面的性状与商品种育种目标一致,测试材料在这几方面的多态性丰富度变化不大。
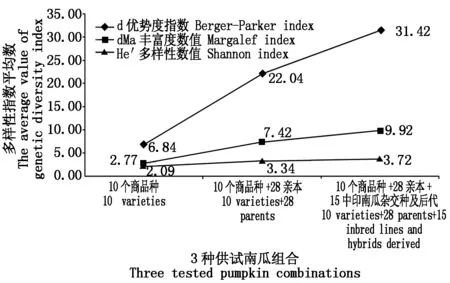
图1 30个形态学性状的3个遗传多样性指数在3种供试南瓜组合中的平均数的变化Fig.1 The changes of three genetic diversity index average value in three tested pumpkin combinations
2.1.2 53份南瓜材料表型聚类分析 采用SPSS软件,根据30个形态学性状在53份南瓜材料间的不同表现进行聚类,结果见图2。在距离22的位置,将供试材料聚为三大类群。第一类包含20份南瓜材料,主要是中印南瓜远缘杂交的亲本及后代组合;第二类有30份南瓜材料,包含印度南瓜商品种及所有印度南瓜骨干亲本;第三类包含3份中国南瓜(10、20、1)。这3个类群中,中印南瓜远缘杂交材料与印度南瓜遗传距离相近。中印南瓜远缘杂交材料拉近了中国南瓜和印度南瓜的遗传距离,有望成为中印南瓜种间杂交的育种桥。
在遗传距离20的位置,可以将第二类群细分为2个亚群。一个亚群是以2-锦栗为代表的印度南瓜
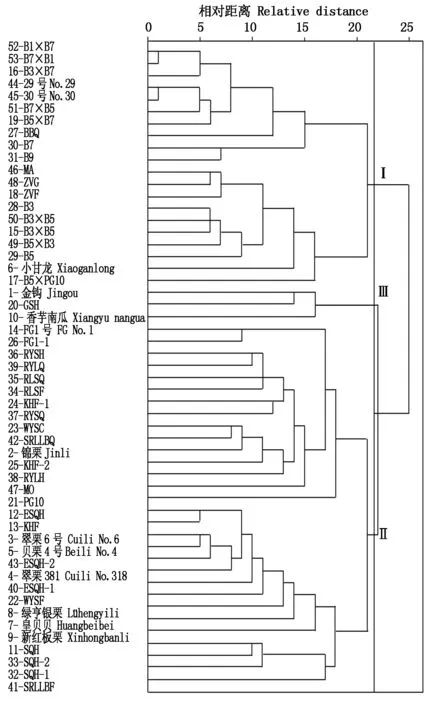
图2 53份南瓜材料形态学性状聚类Fig.2 Dendrogram of 53 pumpkins derived from clustering analysis based on phenotypic traits
亲本;另一个亚群是以3-翠栗6号、5-贝栗4号、4-翠栗318、8-绿亨银栗和9-新红板栗等为代表的印度南瓜亲本。由此可见,供试骨干亲本与市售品种遗传距离较接近,对于改良同一类型南瓜品种有一定作用,但是培育突破性品种,难度较大。
2.2 分子聚类结果
2.2.1 差异引物的多态性分析 试验共使用23对SSR标记,扩增61个位点,平均每个引物扩增2.7个条带。筛选到20个SSR标记,在53份南瓜材料中扩增出55个多态性位点,多态性比例为90%(图3)。
2.2.2 53份南瓜材料分子聚类分析 NTSYS软件通过计算53份材料间的遗传相似系数绘制聚类图(图4)。材料间遗传相似系数在0.520~1.000,以0.680为阈值,可以将供试材料分作3个类群,与南瓜传统分类的3个种对应,分别为中国南瓜(1、10、17、20、21、47)、美洲南瓜(7)和印度南瓜(剩余材料)。这3个种间,中国南瓜与美洲南瓜亲缘关系最近,印度南瓜与中国南瓜和美洲南瓜亲缘关系均较远。

M.Marker(Marker L 50~500 bp)。
在阈值为0.840处,中国南瓜类群可以细分为3个亚群(图4)。第1个亚群包括2个供试中国南瓜商品种,1-金钩和10-香芋南瓜,与骨干亲本21-PG10,三者相似系数为0.958~0.982,亲缘关系近。第2个亚群包含亲本20-GSH和47-MO,其中47-MO是中印南瓜远缘杂交种的母本,第2个亚群与第1个亚群的相似系数为0.88,亲缘关系较远。第3个亚群是17-B5×PG10,这是中印南瓜远缘杂种自交系B5与中国南瓜亲本PG10配置的杂交组合。从聚类结果来看,17-B5×PG10与47-MO相近,与21-PG10较远,但是17-B5×PG10未与MO或者PG10聚在一起,表明利用中印南瓜远缘杂交后代与中国南瓜杂交,会在遗传组成上创造较大变异,产生与现有中国南瓜材料有差别的新组合。
在阈值为0.840处,印度瓜类群被细分为2个亚群(图4),第一亚群包含31份南瓜材料,主要是印度南瓜商品种和印度南瓜骨干亲本;第二亚群包含15份材料,主要是中印南瓜远缘杂交自交系及其父本印度南瓜MA以及自交系自配杂交组合。2个亚群在相似系数0.890的位置被明显分开。在第一亚群中,供试的印度南瓜商品种,2-锦栗和9-新红板栗、3-翠栗6号和4-翠栗318,两两差异不大;6-小甘龙、5-贝栗4号和8-绿亨银栗,亲缘关系较远。印度南瓜骨干亲本23、35、39和41为姊妹系,相似度太高,没有区分开,它们与2-锦栗和9-新红板栗相似。14、32和26与小甘龙近缘;22、27、33、34、38、40、43、36、11和24与5-贝栗4号相似;42、37和45与8-绿亨银栗相似。对于与市售品种相似的亲本,应尽量选择不同类型、遗传相似系数大的亲本相互配组,最大程度地利用杂种优势,改良现有品种。在第二亚群中,中印南瓜远缘杂交自交系29-B5和28-B3与父本印度南瓜MA聚在一起;自交系30-B7和31-B9聚在另外一个亚群中,凡与B7杂交的组合,都被聚在这里。
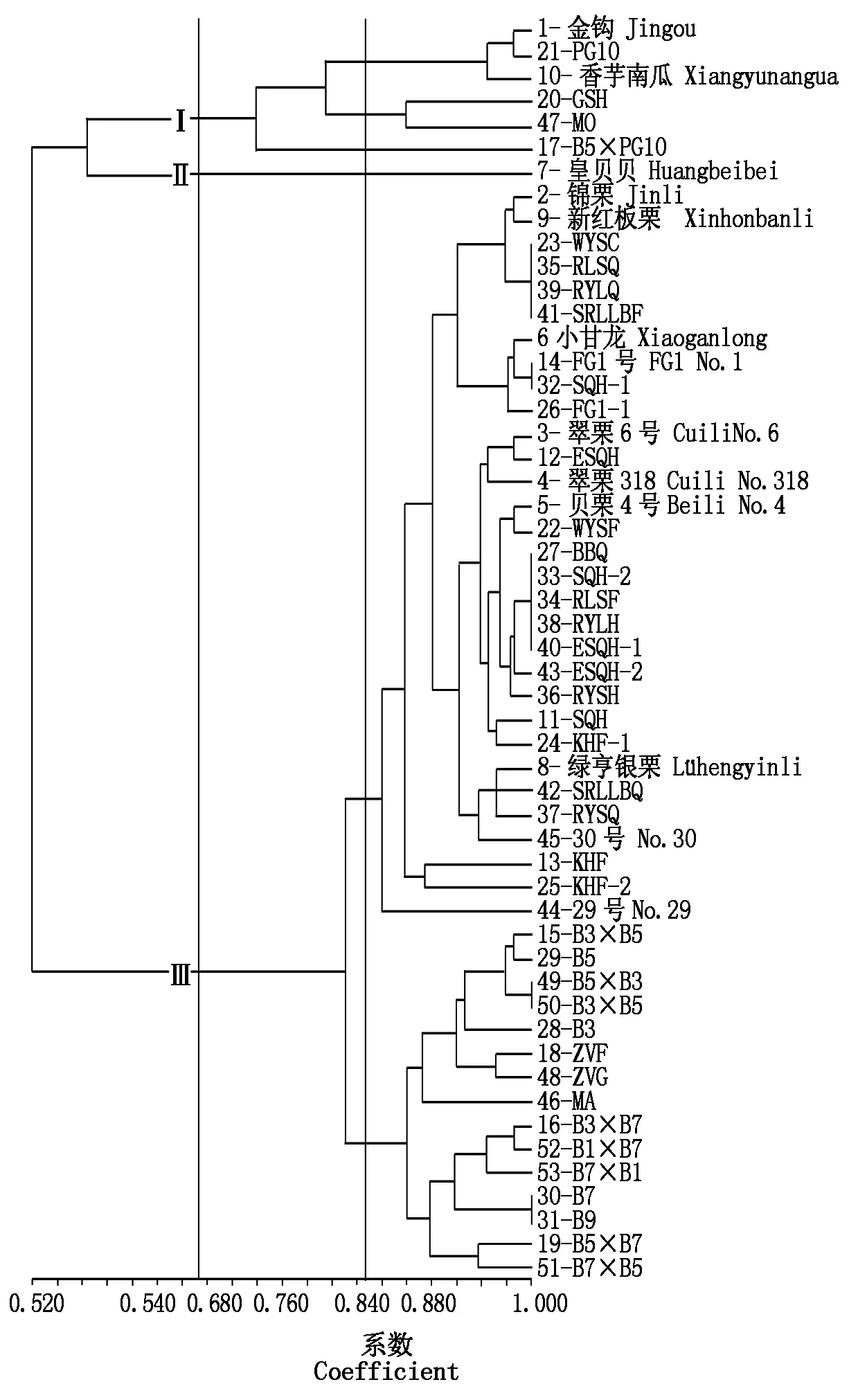
图4 53份南瓜材料SSR分子标记聚类Fig.4 Dendrogram of 53 pumpkins derived from clustering analysis based on SSR molecular markers
2.3 分子聚类与表型聚类的结果比较
53份南瓜材料的分子聚类结果和表型聚类结果有相同之处。首先,2种聚类方法都将中印南瓜远缘杂交后代及组合聚为一类,将印度南瓜商品种及印度南瓜骨干亲本聚为一类,中国南瓜类型与这两类被显著地区分开。根据这个聚类结果,结合30个形态学性状分析发现,中国南瓜类群表现出果型特异性,如长条蜜本、长条香芋等;中印南瓜远缘杂交后代及组合类群表现为果实皮色特异性,如粉彩绿、橘黄色等;印度南瓜商品种和骨干亲本表现为果实大小和果皮颜色的特异性。表明三者之间无论在分子层面还是表型层面都具有显著差异。通过南瓜属种间杂交,增加了现有南瓜品种和育种材料的遗传多样性。其次,2种聚类方法都将供试印度南瓜骨干亲本与印度南瓜商品种进行了比对归类。比如,3-翠栗6号、4-翠栗318和5-贝栗4号,在表型聚类和分子聚类中都比较相近。这些结果都表明,供试印度南瓜商品种和印度南瓜骨干亲本相似度较高,同质化严重,对于培育特色品种缺乏异质基因。此外,对于中印南瓜远缘杂交后代及组合的聚类,2种聚类方法得到结果非常一致,即B7和B9以及与B7杂交的组合都被聚在一起,B3和B5及其杂交组合被聚为一类。B3、B5、B7和B9是中印南瓜杂交种不同株系的自交系,B3和B5相近,B7和B9相近,在育种实践中,2种类型2个株系择其一使用即可。同时,分子聚类与表型聚类的结果也存在不同之处。首先表型聚类不能把美洲南瓜7-皇贝贝和中国南瓜47-MO与印度南瓜区分开;其次表型聚类和分子聚类对骨干亲本与南瓜商品种的归类也不完全相同。
2.4 表型性状主成分分析
根据53份南瓜材料30个表型数据进行主成分分析,前6个主成分累计贡献率为88.17%,30个复杂的表型性状简化为6个主要因子(表4)。第一主成分特征值为15.98,贡献率为36.09%,其特征向量凝聚的生物学信息主要是子房和老果皮色。第二主成分特征值为9.58,贡献率为18.56%,其性状特征值较大的性状为子房和老果的条斑网格。第三主成分特征值为5.22,贡献率为12.42%,果型指数的特征值较大。第四主成分特征值为3.33,贡献率为8.05%,老果皮色和条斑网格是该主成分的特征,综合了第一和第二主成分的老果表皮的生物学信息。第五主成分特征值为2.54,贡献率为7.73%,果实口感评价的特征值较大。第六主成分特征值为1.29,贡献率为5.32%,其性状特征值较大的性状为分枝和萌蘖习性。由此可见,本研究涉及的30个表型性状中,幼果和成熟果实的皮色、果实表面的条斑网格、果型指数、口感和植株分枝萌蘖等性状是区别53份材料的最主要的特征。

表4 53份南瓜材料30个表型数据的主成分分析结果Tab.4 The results of principal component analysis derived from 30 morphological traits of 53 pumpkins
2.5 利用主要因子对53份南瓜材料进行二次聚类
利用主要因子对53份南瓜材料进行二次聚类的结果见图5。在距离15.5的位置,供试材料被聚为8大类群。其中第Ⅰ类、第Ⅱ类和第Ⅵ类包含所有的中印南瓜远缘杂交后代(16份),B7和B9被单独聚在第Ⅱ类。第Ⅲ类、第Ⅳ类和第Ⅴ类包含所有的印度南瓜骨干亲本及商品种。5份中国南瓜(1、10、20、21、47)和1份老果皮色为草绿色、肉质水质丝滑的印度南瓜FG1-1,被聚类到第Ⅶ类。17-B5×PG10被单独聚在第Ⅷ类。
二次聚类的结果与第一次利用30个表型性状进行聚类的结果相比更加准确,与分子聚类结果符合度更高。首先,第一次表型聚类的第Ⅱ类“印度南瓜骨干亲本”中,包含中国南瓜47-MO,二次聚类将它分离出来,与中国南瓜聚在一起,更符合分子聚类结果(图4)。其次,两次聚类结果中16份中印南瓜远缘杂交后代,分布基本相同,但是二次聚类将B7和B9单独聚为第Ⅱ类,将B5×PG10单独聚为第Ⅷ类,与分子聚类结果更吻合。此外,二次聚类将印度南瓜骨干亲本及商品种分为第Ⅲ类、第Ⅳ类和第Ⅴ类,这三类与第一次表型聚类的第Ⅱ类的2个亚类分布不同。与分子聚类结果更符合的是二次聚类结果。主要表现在分子聚类将2、9、23、35、39聚为一类,这一类与其他印度南瓜材料遗传距离较远;这个结果与二次聚类将37、38、24、35、42、25、27、9、23、36、39、2、14聚为第Ⅳ类,与其他材料(第Ⅲ和Ⅴ类)分开的结果吻合。而第一次表型聚类结果中2、9、23、35、39被分散到第Ⅱ类的2个亚群,区分不明显(图2)。因此,利用主要因子对53份南瓜材料进行二次聚类,获得了与分子聚类具有较高符合度的结果,这些主成分因子可作为简化表型性状统计的指标。
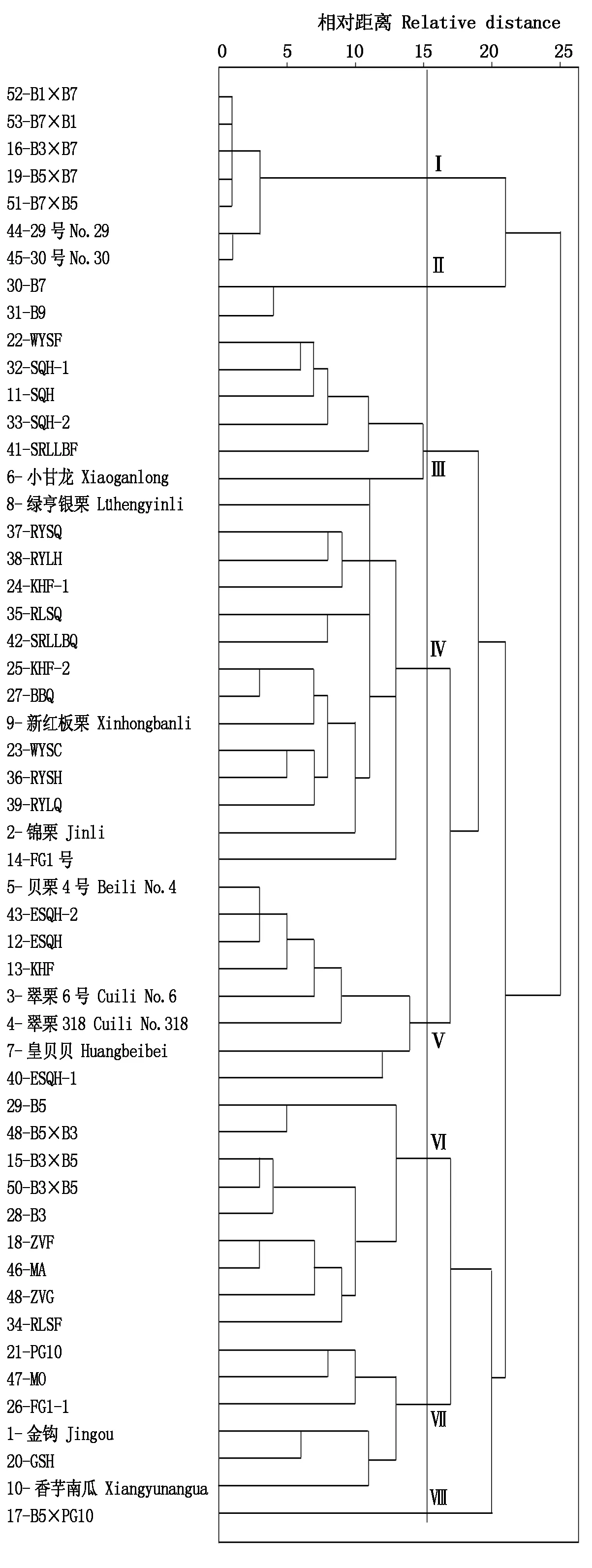
图5 基于6个主成分因子的53份南瓜材料形态学性状聚类Fig.5 Dendrogram of 53 pumpkins derived from clustering analysis based on the 6 principal component factors
3 结论与讨论
本研究在选择食用南瓜商品种时,主要是针对老熟果蒸煮食用的南瓜品种。对美洲南瓜类型,只选择食用方式和表型特征与此类南瓜相似的7-皇贝贝,而没有选择长条笋瓜和圆形笋瓜的品种。主要是考虑到长条或者圆形笋瓜,以食用嫰果为主,且笋瓜材料种间遗传组成差异较大[34],笋瓜与中国南瓜和印度南瓜差异较大[35],因此,增加笋瓜类型,会使供试材料生物多样性指数和遗传相似系数异常,不利于本研究数据分析。
供试材料的分子聚类与第一次利用30个表型性状进行聚类的结果不完全统一的主要原因,可能是由于试验中使用的分子标记,虽然是在前期SSR扩增试验中具有稳定多态性的引物,但是数目有限,不能完全代表供试材料广泛的遗传背景[36],同时,这些分子标记与30个表型性状也不能完全对应[37],因此导致二者的聚类结果不能统一。主成分分析被广泛应用于作物表型性状的核心指标筛选[38],提高表型聚类与分子聚类的吻合度[39],是现代育种的重要方法。从30个表型性状的主成分分析中,根据贡献率得到6个主成分,包括子房和成熟果实的皮色、果实表面的条斑网格、果型指数、口感和植株分枝萌蘖等8个性状。利用这8个性状对53份南瓜材料进行二次表型聚类,得到了与分子聚类符合度更高的结果。其原因可能在于,不同表型性状对主成分因子贡献率不同[38]。剔除叶色、株型、株高、花期等在供试材料间表现较为一致、贡献率较低的表型特征,减少表型聚类的干扰因素,突出贡献率较大的主成分因子,最终获得较为准确的聚类结果。这样,一方面解决了表型聚类与分子聚类结果不统一的问题,另一方面为今后食用南瓜性状调查简化了调查指标,避免了性状间相互作用造成的重复选择。
本研究的3次聚类结果均表明,现有印度南瓜品种及育种材料在表型和分子层面亲缘关系较近,存在同质化。周秦等[14]对26份南瓜种质进行遗传多样性分析,也发现同样问题,并且指出,同质型南瓜亲本或者品种即便再组合,也还会和原始亲本聚在一起。高斌等[40]在对北方花生育成品种以及骨干亲本进行分子标记鉴定和系谱分析时,发现相同的花生骨干亲本在不同地区被反复利用,导致花生品种间相似度升高。研究指出,加大相对封闭地区的南瓜材料引种,以及利用种间杂交,能够拓宽南瓜遗传多样性[41]。3次聚类都将中印南瓜种间杂交自交系及其新配组合聚为一类,与供试印度南瓜商品种及骨干亲本明显区分开。这与王瑞等[24]得到的结果相同。他们在研究95份南瓜资源亲缘关系的时候发现,远缘杂交中间类型被聚在一起,形成一个分枝。这些都显示了种间杂交对增加南瓜遗传多样性的有效性。通过种间杂种,为培育特色南瓜新品种提供异质基因,推陈出新。供试材料中,18-ZVF和48-ZVG是利用中印南瓜远缘杂种自交系与骨干亲本自配杂交组合,植株耐热,果肉粉香,淀粉含量高;在参加2021年新疆昌吉中国园艺学会南瓜研究分会南瓜新品种展示会时,受到承办方的好评。
猜你喜欢
西南农业学报(2022年5期)2022-06-06
中国糖料(2021年3期)2021-07-13
阅读与作文(小学高年级版)(2019年5期)2019-05-27
热带农业科技(2019年1期)2019-01-14
中国果业信息(2019年11期)2019-01-05
现代园艺(2017年21期)2018-01-03
中国康复理论与实践(2015年10期)2015-12-24
医学研究杂志(2015年5期)2015-06-10
现代检验医学杂志(2015年5期)2015-02-06
