
水稻OsMYB57基因表达的调控与其化感抑草作用*
2022-02-15 04:43:42杨陆可高钰杰林文雄方长旬
中国生态农业学报(中英文) 2022年2期
杨陆可,王 浩,高钰杰,严 雪,母 丹,林文雄**,方长旬,2**
(1.福建农林大学生命科学学院农业生态研究所/福建省农业生态过程与安全监控重点实验室 福州 350002;2.作物生态与分子生理学福建省高校重点实验室(福建农林大学) 福州 350002)
植物化感作用是一种普遍的生态学现象,广泛存在于农田、森林、海洋等生态系统中,农作物、林木、藻类等不同类型的植物均具有化感品种。在农田生态系统,化感水稻(L.)的根系分泌大量的酚酸类、黄酮类、二萜类等化感物质抑制周围杂草的生长。研究表明,化感水稻田间抑草率可达30%~40%,且对水稻产量和品质的影响小。利用化感水稻控制杂草也是可持续生态农业的一种发展趋势。
酚酸类化合物是一类重要的水稻化感物质。化感水稻‘PI312777’的酚酸含量极显著高于非化感水稻‘Lemont’,其中6 叶期‘PI312777’的酚酸含量达710 μg∙株。这些化感物质还能与土壤中特定的微生物相互作用,增强抑草能力。研究发现,化感水稻分泌的阿魏酸与黏细菌相互作用对稗草[(L.) Beauv.]抑制率达64.82%,显著高于单独阿魏酸(34.64%)及单独黏细菌(8.26%)的化感作用。酚酸类化合物由苯丙烷代谢途径合成,苯丙氨酸解氨酶(PAL)是该途径的关键酶。化感水稻的基因表达水平显著高于非化感水稻,这与两种水稻中基因启动子的活性密切相关。李兰兰等比较了‘PI312777’和‘Lemont’中相同基因成员的启动子活性差异,结果发现‘PI312777’中第2 染色体的第3 个成员基因()启动子的活性高于 ‘Lemont’,并推测这是导致‘PI312777’中酚酸类化合物含量高于非化感水稻‘Lemont’的原因之一; 过量表达后水稻的酚酸类化合物总量增加,抑草能力提高。此外,茉莉酸甲酯(MeJA)和水杨酸甲酯(MeSA)等小分子物质诱导可增强基因的表达,促进酚酸类化合物的积累。
植物体内酚酸等次生物质的合成与转录因子的调控作用有关。MYB 是一类能够调控次生代谢的转录因子。植物中的MYB 转录因子根据MYB 域内相邻重复序列的数量分为4 种类型,分别为1R-MYB、R2R3-MYB、3R-MYB 和4R-MYB,它们分别含有1 个、2 个、3 个和4 个MYB 重复序列。在这4种类型中,R2R3-MYB 因子是MYB 家族的主要成员,参与调控次生代谢相关基因的表达。研究表明,组成型表达能促进玉米(L.)中阿魏酸和绿原酸的积累; 在烟草(L.)、马铃薯(L.)、番茄(L.)和雪莲[(Kar.et Kir).Sch.-Bip.] 4 种植物中分别组成型表达的马铃薯、、基因和拟南芥(L.)中的基因也促使绿原酸含量的增加。葡萄(L.)的也参与了苯丙烷代谢途径。在拟南芥中,MYB11、MYB12、MYB111 等R2R3-MYB 类 型 的MYB 转录因子能够调控查耳酮异构酶、黄烷酮3-羟化酶和黄酮醇合成酶1 的基因表达,这些基因参与了类黄酮的生物合成,从而调控植物黄酮类化合物积累。利用转录激活因子VP64 增强水稻中的表达后,水稻苯丙烷途径中、、和基因也上调表达,L-苯丙氨酸含量增加,水稻对稗草的抑制率提高。对MYB57 转录调控的基因进行研究,结果显示MYB57转录调控的表达,MAPK11 可与PAL2;3相互作用从而调控水稻酚酸类化感物质的合成。在此基础上,揭示调控基因表达的转录因子能够进一步明确水稻化感作用特性形成的分子调控网络。据此,本文利用DNA-pull Down 分离了调控基因表达的转录因子,并研究其外源调控方式,认识MYB57 调控水稻化感作用的作用网络。
1 材料与方法
1.1 供试材料
供试水稻材料为国际公认的化感水稻品种 ‘PI312777’和非化感水稻品种‘Lemont’,供试受体材料为稗草。
1.2 供试水稻材料的种植
取‘PI312777’和‘Lemont’水稻种子,用25% NaClO表面消毒30 min 后清洗干净,加入灭菌水于30 ℃培养箱中浸泡16 h。倒掉无菌水,在恒温培养箱中30 ℃催芽,期间保持种子湿润。待水稻长至高度约5 cm时将其移栽至KT 板(4×8 孔)上,每板种植16 棵,后放置于培养盆(70 cm×40 cm×30 cm) 中进行水培,光照强度360 μmol∙m∙s,相对湿度80%~85%,盆的外表均匀涂黑以防止产生绿藻,每周更换新鲜配制的完全营养液。水稻长至3 叶一心时,将提前萌发好长势一致的稗草(长出2~3 叶,株高4~5 cm 左右)种植在剩余的16 孔中,稗草与水稻在盆中对称种植,对照组种植32 株水稻,继续在温室中培养。处理和对照各4 个重复。
在与稗草共培处理1 d、3 d、5 d、7 d 后,分别取样水稻的倒二片完全展开的功能叶和根系,液氮速冻,用于基因组DNA、总RNA 以及叶片可溶性总蛋白的提取。
1.3 茉莉酸甲酯处理化感水稻‘PI312777’和非化感水稻‘Lemont’
将萌发的‘PI312777’和‘Lemont’种子种植于96孔板,每板种植30 棵,并置于1 L 黑色塑料盆中培养,每周更换营养液。待长至3 叶一心时,分别添加终浓度为0.01 mmol∙L、0.02 mmol∙L、0.05 mmol∙L、0.1 mmol∙L和0.2 mmol∙L茉莉酸甲酯(MeJA)于水稻完全营养液中作为处理组,以不添加MeJA 为对照组,处理组与对照组均设4 个重复。在处理1 d、3 d、5 d 和7 d 时,分别取处理组和对照组水稻根系和倒二片完全展开的功能叶,液氮速冻,用于提取总RNA 和蛋白。
1.4 OsMYB57 基因启动子克隆及其转录调控因子分离
采用植物基因组DNA 提取试剂盒(CW0553S,北京康为世纪生物科技有限公司)提取水稻‘PI312777’和‘Lemont’的基因组DNA。采用5′端带生物素(biotin)标记的引物-promoter-F:5′-GCAGTA ACAAGCTAACAGCAGCTGG-3′与-promoter-R:5′-CTTCTTTTGCTTCCTCCCTCCTGAG-3′分别从两种水稻的基因组DNA 中扩增基因启动子用于 DNA-pull down 获得启动子的结合蛋白。DNA-pull down 根据Fang 等方法,提取‘PI312777’和‘Lemont’叶片的可溶性蛋白,使用Dynabeads kilobaseBINDER™ Kit (Invitrogen,Thermo Fisher Scientific)试剂盒收集DNA 片段中结合的蛋白质并进行SDS-PAGE,切取目标蛋白条带酶解,蛋白质鉴定在质谱仪器 Q-Exactive HF (Thermo Scientific)上进行。
1.5 qPCR 检测OsMYB57 基因启动子结合蛋白的基因表达量
采用TRIzol 法分别提取稗草共培以及茉莉酸甲酯处理1 d、3 d、5 d 和7 d 的‘PI312777’与‘Lemont’水稻叶片、根系的总 RNA 并纯化,总RNA 经Easy-Script One-Step gDNA Removal and cDNA Synthesis SuperMix 试剂盒(北京全式金生物技术有限公司)逆转录成cDNA。以MYB57 启动子结合蛋白的编码基因序列为模板设计qPCR 引物,并以肌动蛋白(actin)基因为内参基因(表1)。按照TransStart Tip Green qPCR SuperMIX 试剂盒说明书(北京全式金生物技术有限公司)配置qPCR 反应体系,qPCR 在realplex荧光定量PCR 仪(Eppendorf,Germany)中完成,根据生成的阈值(CT 值),运用2法计算基因的相对表达量。
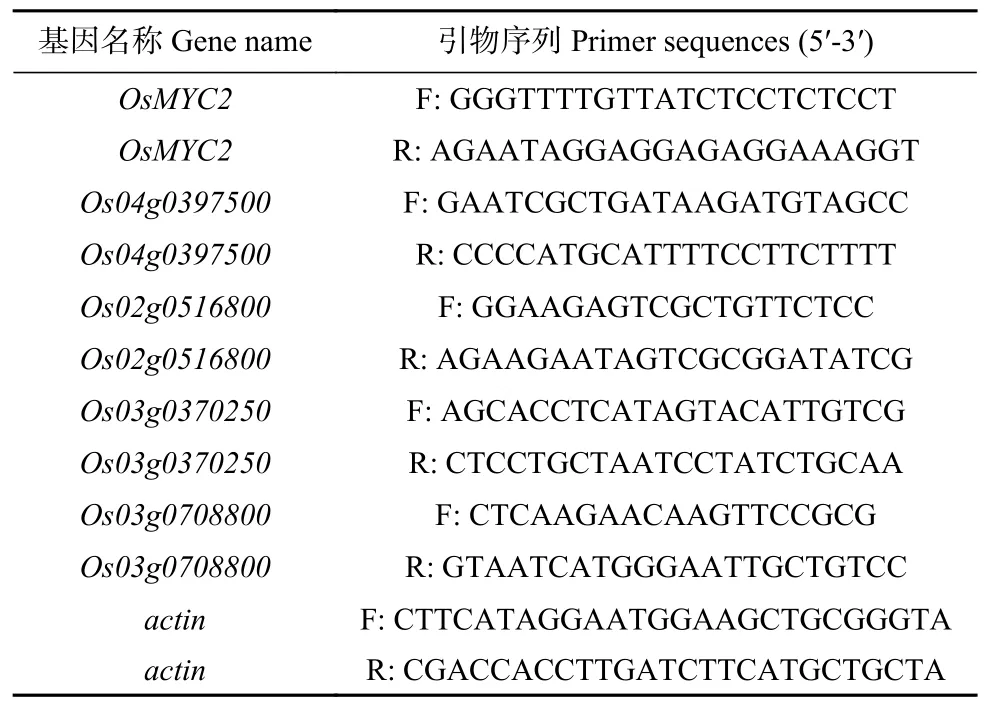
表1 本研究中采用的qPCR 引物Table 1 qPCR primers used in this study
1.6 不同浓度茉莉酸甲酯处理下水稻的蛋白表达量检测
采用TCA 丙酮法提取蛋白,并在10%/的聚丙烯酰胺凝胶电泳分离蛋白。采用湿转(80 V,2 h)的方法将蛋白条带固定于PVDF 膜上,5%脱脂牛奶封闭过夜后,分4 组与MYB57、MAPK11、PAL2;3 和Actin 的特异抗体分别孵育1.5 h。MYB57、MAPK11和PAL2;3 的二抗采用Goat Anti-Rabbit IgG (H+L),actin 的二抗使用Goat Anti-Mouse IgG (H+L),孵育1.5 h 后进行ECL (electrochemiluminescence)显色,并于ChemiDoc MP (Biorad)中进行成像。
1.7 不同浓度茉莉酸甲酯处理下水稻分泌液的抑草率评价
将稗草种子萌发,培养方式同1.3。分别将不同浓度茉莉酸甲酯处理的两种水稻的根系液进行过滤除杂后各收集1.5 L 滤液,然后将长至两叶一心的稗草分别种植于上述收集的水稻根系液中,每种处理种植25 棵稗草并添加500 mL 滤液,种植第14 d 后对各处理稗草根和叶的长度进行测定。将根茎分开杀青,烘干至恒重后测量其干重,并计算抑制率(%)。抑制率计算公式为 IR=(1−TR/CK)×100%。其中IR为抑制率(inhibitory ratio),TR 为处理组生长指标(treatment),CK 为对照组生长指标(control)。数据采用DPS 软件进行统计分析。
2 结果与分析
2.1 OsMYB57 基因启动子的转录调控因子
通过DNA-pull down 获得‘PI312777’和‘Lemont’的基因启动子所结合的蛋白,SDS-PAGE检测结果显示,‘PI312777’和‘Lemont’的基因启动子DNA 上均含有结合蛋白,对照组未经生物素标记的基因启动子DNA 上未检测到结合蛋白(图1)。
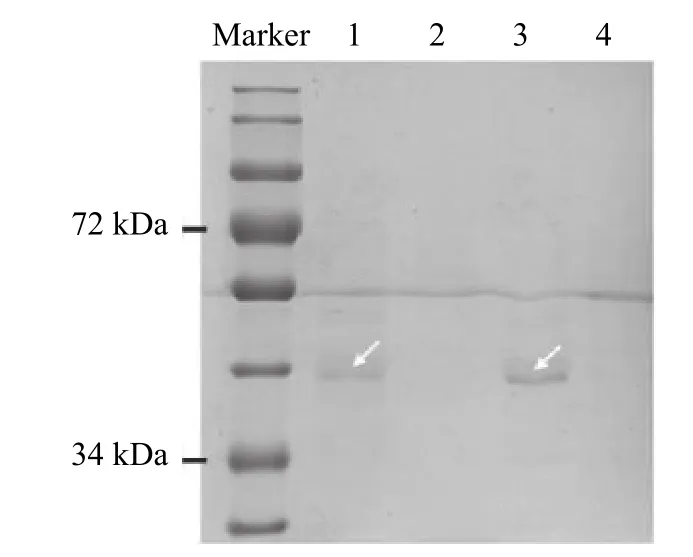
图1 ‘PI312777’和‘Lemont’水稻中OsMYB57 基因启动子的结合蛋白Fig.1 Proteins binding on the promoter of OsMYB57 from rice varieties ‘PI312777’ and ‘Lemont’
对启动子上结合的蛋白进行质谱鉴定和分析,结果从‘PI312777’水稻的基因启动子的结合蛋白中鉴定到具有结合DNA 功能的蛋白,包括转录因子basic helix-loop-helix protein 009 (基因:)、LOC_Os04g32590.1 (基因:)、LOC_Os0 2g31160.1 (基因:)、转录调节因子LOC_Os03g25430.1 (基因:)和LOC_Os03g 50110.1 (基因:); 在‘Lemont’水稻的基因启动子鉴定到具有结合DNA 功能的蛋白,包括转录因子LOC_Os04g32590.1、LOC_Os0 2g31160.1 和转录调节因子LOC_Os03g25430.1 (表2)。
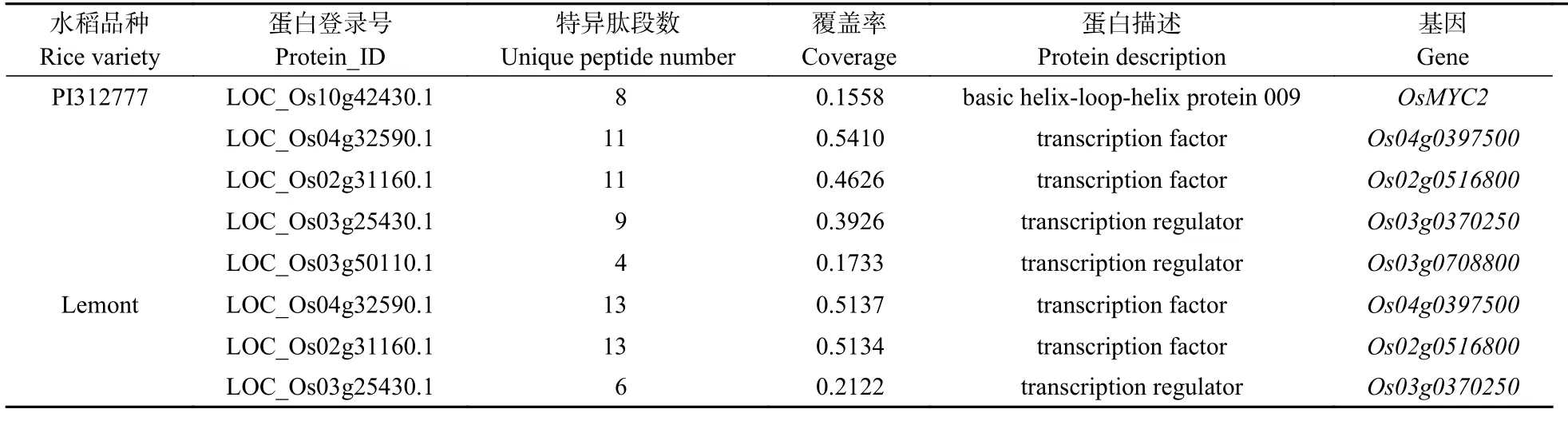
表2 ‘PI312777’和‘Lemont’水稻中OsMYB57 基因启动子上的结合蛋白鉴定结果Table 2 Identification of the proteins binding on the OsMYB57 gene promoter from rice varieties ‘PI312777’ and ‘Lemont’
2.2 OsMYB57 基因启动子转录因子的基因表达量
基因表达由生物体内反式作用因子(转录因子)作用于顺式作用原件(启动子区域)所决定。对稗草共培下的‘PI312777’和‘Lemont’中的基因启动子上结合的转录调节因子的基因表达量进行检测,qPCR 结果显示,稗草共培下的‘PI312777’水稻根系和叶片中的、、和基因表达都较其单独培养的对照组上调,具体表现为先上升后下降,且在根中的上调表达量明显高于叶。在根中,和在5 d 时上调表达量最高,分别上调100.19 倍、146.44 倍和23.98 倍; 其在相同处理下的‘Lemont’根系中分别上调4.95 倍、10.22 倍和9.36 倍。稗草胁迫下的‘Lemont’叶片中这3 个基因的上调表达倍数较大,其在共培5 d 处理组中,表达量分别上调35.11 倍、50.53 倍和40.97 倍(图2)。

图2 稗草胁迫下‘PI312777’和‘Lemont’水稻中OsMYB57 基因转录调控因子的动态表达Fig.2 Dynamic gene expression levels of transcriptional regulators of OsMYB57 in rice varieties ‘PI312777’ and ‘Lemont’ co-cultured with barnyardgrass
2.3 茉莉酸甲酯处理下水稻根系中基因的动态表达
bHLH009 是茉莉酸(JA)信号途径的调节蛋白,在JA 介导的信号转导途径中起重要作用。采用qPCR分别检测了茉莉酸甲脂(MeJA)处理1 d、3 d、5 d 和7 d ‘PI312777’和‘Lemont’根系中的较其各自对照组的表达变化。在‘PI312777’根系中,不同浓度茉莉酸甲酯处理促进了这5 个转录因子的基因表达,其中0.05 mmol∙L、0.10 mmol∙L和0.20 mmol∙L的MeJA 对基因表达的促进效果较为明显,0.05 mmol∙LMeJA 诱导7 d 的‘PI312777’中和基因的表达量为相同天数对照组的2.67 倍、5.66 倍、4.90 倍、3.18倍和3.21倍; 0.10 mmol∙LMeJA 诱导7 d处理组分别为对照组的97.39 倍、378.72 倍、159.97 倍、243.59 倍和146.94倍; 0.20 mmol∙LMeJA诱导组分别为对照组的28.36 倍、244.72 倍、54.95 倍、30.91 倍 和35.82 倍(图3a-e)。
在‘Lemont’中,MeJA 对上述5 个基因的诱导作用不及相同处理下的‘PI312777’中上调表达程度,0.01 mmol∙LMeJA 处理1 d 的‘Lemont’根系中基因上调表倍数最大,分别为对照组的39.01 倍、28.99 倍和36.69 倍; 0.20 mmol∙LMeJA 处理3 d 的‘Lemont’根系中的上调表达倍数最大,为对照组的16.33 倍和53.26 倍(图3f-j)。
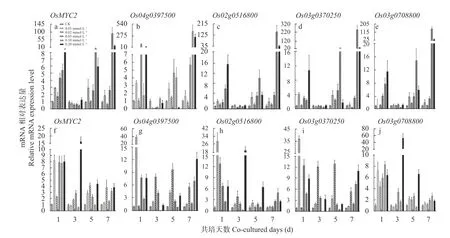
图3 不同浓度茉莉酸甲酯处理不同时间‘PI312777’和‘Lemont’水稻根系中基因的动态表达Fig.3 Dynamics of gene expression level in the root of ‘PI312777’ and ‘Lemont’ under treatments of different concentrations of methyl jasmonate
2.4 茉莉酸甲酯处理下水稻叶片中基因的动态表达
qPCR 同时检测了MeJA 诱导下的‘PI312777’和 ‘Lemont’叶片中和基因的动态表达。结果显示,叶片中的这些基因上调表达程度较其根系中低。‘PI312777’叶片中,MeJA 诱导7 d的基因上调表达程度相对较大,其中以0.02 mmol∙LMeJA 处理的效果最明显,5 个基因分别较对照组上调4.71 倍、3.33 倍、7.25 倍、3.67 倍和4.25 倍(图4a-e)。
在‘Lemont’叶 片 中,在0.10 mmol∙LMeJA 诱导5 d 的上调表达倍数最大,为对照组的4.94 倍;和基 因分 别在0.05 mmol∙L和0.20 mmol∙LMeJA 处理7 d 时的上调表达倍数最大,为对照组的14.70 倍和4.09 倍; 而0.05 mmol∙LMeJA 处理3 d 对和基因表达的诱导效果最明显,分别为对照组的18.06 倍和20.22 倍(图4f-j)。可见,MeJA 处理后,‘Lemont’水稻叶片中的这5 个基因的响应作用存在较大差异。
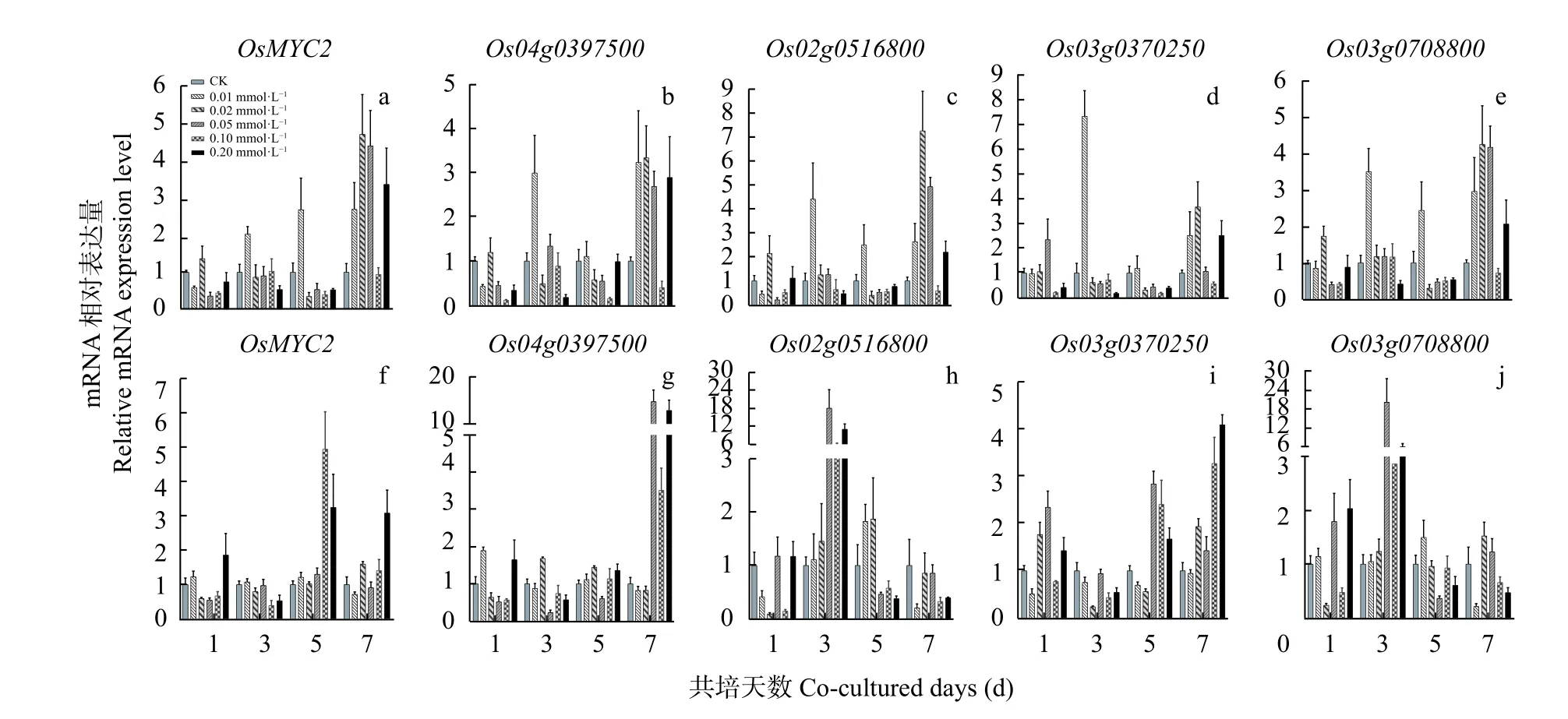
图4 不同浓度茉莉酸甲酯处理不同时间‘PI312777’和‘Lemont’水稻叶片中基因的动态表达Fig.4 Dynamics of gene expression level in the leaves of ‘PI312777’ and ‘Lemont’ under treatments of different concentrations of methyl jasmonate
2.5 茉莉酸甲酯处理下MYB57、PAL2;3 及MAPK11的蛋白表达
进一步检测了MeJA 诱导处理下,bHLH009 等转录因子调控的下游基因,以及MYB57 蛋白调控的和基因编码的蛋白表达量。0.10 mmol∙L和0.20 mmol∙LMeJA 处理下,‘Lemont’中MYB57 和MAPK11 蛋白表达量均明显提高; PAL2;3 在处理组的表达量也高于其在对照组中的表达,但不明显。在PI312777 中,0.05 mmol∙LMeJA 处理下MYB57 和MAPK11 及0.10 mmol∙LMeJA 处理下的PAL2;3 蛋白表达量较对照组明显提高。且‘PI312777’中的PAL2;3 和MYB57 的表达量均高于相同处理条件下的‘Lemont’。结果表明,MeJA 处理能够诱导提高‘PI312777’和‘Lemont’中MYB57、PAL2;3 和MAPK11 蛋白的表达量,相同浓度的MeJA 对‘PI312777’的诱导作用大于对‘Lemont’的诱导作用(图5)。
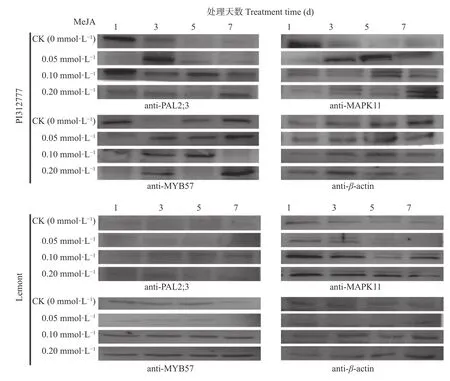
图5 不同浓度茉莉酸处理不同时间‘PI312777’和‘Lemont’水稻中MYB57、PAL2;3 和MAPK11 蛋白的表达量Fig.5 Protein expression of MYB57,PAL2;3 and MAPK11 in ‘PI312777’ and ‘Lemont’ under treatments of different concentrations of methyl jasmonate for different days
2.6 不同浓度茉莉酸甲酯处理对两水稻品种生长的影响
比较MeJA 处理下‘PI312777’和‘Lemont’的生长情况,结果显示,随着处理天数的增加,0.01 mmol∙L、0.02 mmol∙L和0.05 mmol∙L浓度的MeJA 处理能促进水稻的生长。但0.1 mmol∙L和0.2 mmol∙L浓度处理3 d 开始对两品种水稻的生长产生抑制作用,即表现为低浓度促进而高浓度抑制的生理现象,外源添加MeJA 的浓度不高于0.05 mmol∙L时,对两品种水稻的生长具有一定的促进作用(图6)。

图6 不同浓度茉莉酸甲酯(MeJA)处理不同时间 ‘PI312777’和‘Lemont’水稻的生长状况Fig.6 Plant phenotype of ‘PI312777’ and ‘Lemont’ under treatmetns of different concentrations of methyl jasmonate (MeJA) for different days
2.7 茉莉酸甲酯处理下水稻根系分泌液的抑草率
进一步测定了MeJA 对稗草生长的影响。结果显示,与 未添加MeJA 的对照组相比,0.05 mmol∙LMeJA 对稗草的根长和株高均不产生影响; 0.10 mmol∙LMeJA 对稗草的根长不产生影响,但显著抑制了稗草的株高; 0.20 mmol∙LMeJA 对稗草的根长和株高均具有显著的抑制作用(图7)。

图7 不同浓度茉莉酸甲酯(MeJA)对稗草生长的影响Fig.7 Effect of different concentrations of methyl jasmonate (MeJA) on barnyardgrass growth
随着MeJA 添加浓度的增大,‘PI312777’和‘Lemont’根系分泌液对稗草株高的抑制作用不断增强,并且都在0.20 mmol∙LMeJA 处理下抑制稗草株高最明显; 随添加MeJA 浓度的提高,增强了‘PI312777’根系分泌液对稗草根长的抑制效果,但对‘Lemont’根系分泌液抑制稗草根长则作用不明显(图8)。
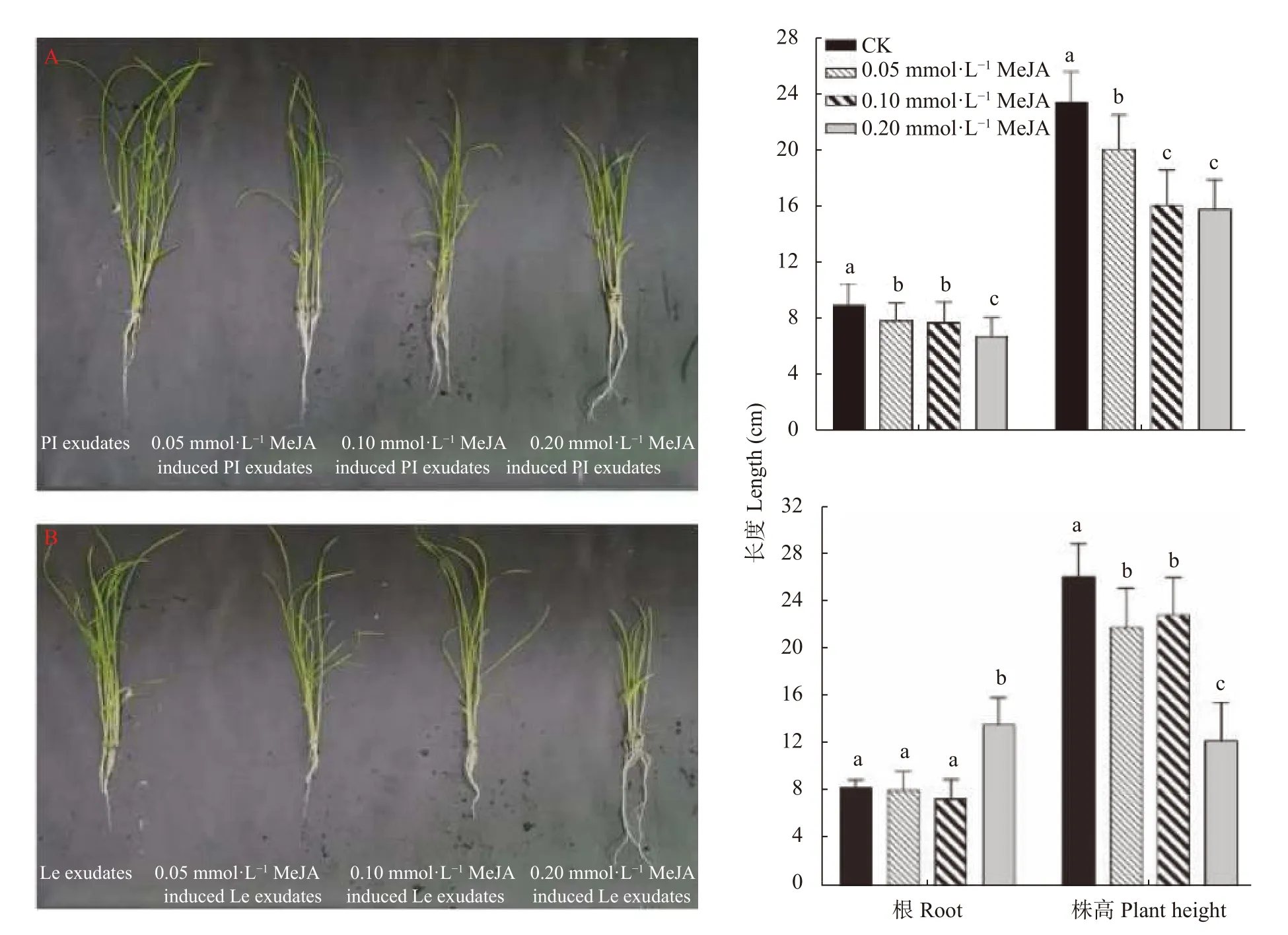
图8 不同浓度茉莉酸甲酯处理下‘PI312777’ (PI)和‘Lemont’ (Le)水稻根系分泌液对稗草生长的影响Fig.8 Effect of root exudates of ‘PI312777’ (PI) and ‘Lemont’ (Le) rice induced by different concentrations of methyl jasmonate(MeJA) on barnyardgrass growth
图9 结果可以看出,在化感水稻‘PI312777’中,0.05 mmol∙L、0.10 mmol∙L和0.20 mmol∙LMeJA处理的根系分泌液对稗草地上部分干重的抑制率为25.82%、30.52%和32.59%,表现出随添加 MeJA 浓度的提高抑制率增强的趋势; 对稗草地下部分干重的抑制率分别为17.01%、30.65 %、30.02%。在非化感水稻‘Lemont’中,上述3 种MeJA 浓度处理的根系分泌液对稗草地上部分干重的抑制率为5.30%、27.85%、35.50%,对稗草地下部分干重的抑制率分别为15.36%、15.13%、−6.44% (图9)。上述结果表明,MeJA 能够诱导增强水稻的化感作用,提高两种水稻对稗草的抑制率。
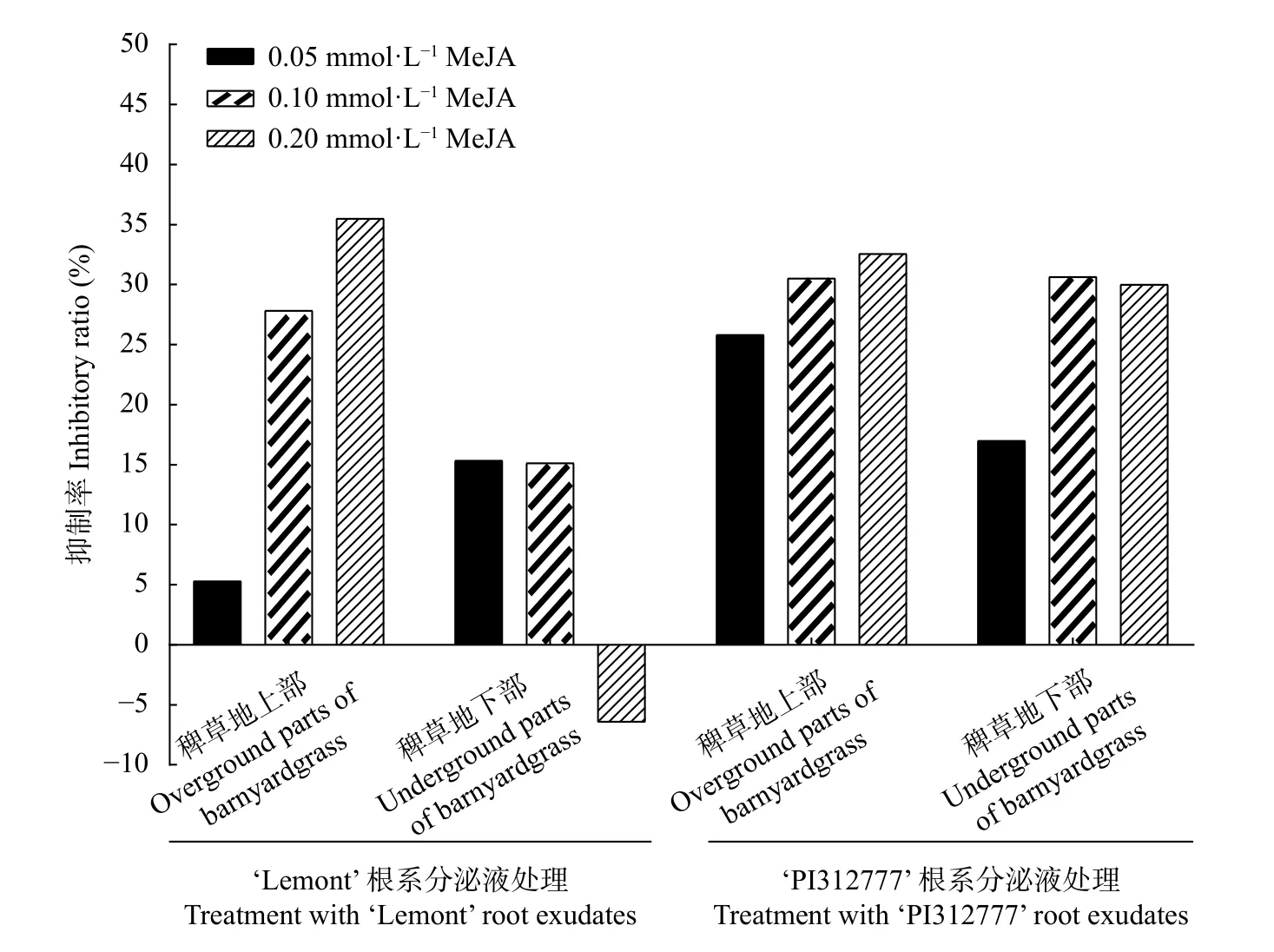
图9 不同浓度茉莉酸甲酯添加下的‘PI312777’和‘Lemont’水稻根系分泌液对稗草干重的抑制率Fig.9 Inhibitory rates of root exudates of ‘PI312777’ and ‘Lemont’ rice induced by different concentrations of methyl jasmonate(MeJA) to the dry weight of barnyardgrass
3 讨论与结论
植物化感物质合成与分泌作用是决定化感潜力的关键因素。化感水稻能够通过苯丙氨酸代谢途径合成酚酸类化合物对邻近稗草产生化感抑制作用。苯丙氨酸解氨酶是苯丙氨酸代谢途径的限速酶,其活性高低决定了水稻对酚酸类化感物质的生物合成能力。植物体内次生代谢物的合成受到多种转录因子的调控。前期研究发现增强一个R2R3-MYB 转录因子基因的表达能够使苯丙氨酸途径中上调表达;染色质免疫共沉淀结果显示MYB57 能够通过转录调控基因的表达,MAPK11 再与PAL2;3蛋白发生相互作用,间接调控水稻的苯丙烷代谢,表明了有效调控水稻的化感抑草能力。
化感作用关联基因表达的强度与其启动子上转录因子的调控密切相关。对‘PI312777’和‘Lemont’的基因启动子上的转录因子进行研究,在 ‘PI312777’的基因启动子的结合蛋白中鉴定到LOC_Os04g32590.1、LOC_Os02g31160.1、LOC_Os03g25430.1、bHLH009 等具有转录调控作用的蛋
白,在‘Lemont’的基因启动子的结合蛋白中鉴定到LOC_Os04g32590.1、LOC_Os02g31160.1、LOC_Os03g25430.1 等具有转录调控作用的蛋白。其中,bHLH009 属bHLH-TFs 转录因子家族,是茉莉酸(JA)信号途径的调节蛋白,参与多种生物以及非生物胁迫响应过程。大量研究表明,JA 能通过与其他激素或转录因子间的互作完成很多发育过程的调控。MYC 参与调控大量JA 响应基因,能够与JAZ 蛋白结合,是JAZ 抑制剂的靶点。MYC2 是第一个被报道的家族成员,它与MYC3、MYC4 和MYC5 共同作用,响应JA 并介导调控植物生理过程,如根系生长抑制、肥力以及对病原体和昆虫的抗性,MYCs 因而被认为是连结环境和植物反应的重要枢纽。稗草胁迫下,‘PI312777’和‘Lemont’水稻中的基因表达均上调,表明稗草作为环境胁迫因子能够触发水稻的JA 信号转导途径; 与‘Lemont’相比,‘PI312777’根系中基因的上调表达增加倍数更为显著,可见‘PI312777’对稗草胁迫的响应作用更加灵敏,有利于‘PI312777’化感抑草作用的形成。此外,‘PI312777’与‘Lemont’相比,其基因的上调表达倍数更高,使得bHLH009 转录调控的基因在‘PI312777’中的表达丰度也高于其在‘Lemont’中的表达,这也从一方面解释了化感水稻 ‘PI312777’中基因表达水平较高的原因。
外源添加MeJA 处理下,‘PI312777’和‘Lemont’中的基因上调表达,其在根系中的上调表达倍数高于叶片中的上调倍数,且随着MeJA 添加浓度的提高,基因上调表达倍数增加,并以0.10 mmol∙L浓度处理的诱导效果最为显著,bHLH009 转录调控的下游基因以及与MYB57 相互作用的MAPK11 的基因和蛋白表达水平也随着上调。外源MeJA 处理下及MAPK11 的表达变化表明了化感水稻中基因对JA 信号的有效响应以及JA 信号分子诱发的调控作用。MeJA 诱导也促进了水稻根系分泌化感物质,其中0.20 mmol∙L的MeJA 诱导使得‘PI312777’的化感抑草能力提高效果最为显著,但该浓度对水稻的生长也产生一定的抑制作用; 0.05 mmol∙LMeJA 对水稻生长还有一定的促进作用。Bi 等研究了外源分别施用MeJA 和水杨酸甲酯(MeSA)对水稻化感作用的影响,结果也显示MeJA 和MeSA 处理均提高了水稻PAL 酶和肉桂酸4-羟化酶(C4H)的基因表达丰度和酶活,促进了水稻酚类物质的积累,且水稻叶片提取液对稗草的抑制作用也增强。Fang等采用RNA 干扰和基因过量表达技术分别抑制和增强了‘PI312777’的JA 合成关键基因(allene oxide cyclase)表达,结果发现过表达基因使得水稻的内源JA 含量增加,、、等苯丙烷代谢相关基因上调表达,水稻中原儿茶酸、对羟基苯甲酸、阿魏酸、苯甲酸和肉桂酸等酚酸类化合物含量增加,水稻化感抑制稗草作用提高; 抑制表达则结果相反。这些结果也证实了JA 及其合成对水稻化感抑草能力的调控作用。本研究在此基础上,进一步揭示了化感水稻响应JA 的受体蛋白bHLH009 及其下游调控基因,从而促进酚酸类化感物质合成的分子机制。此外,已有的研究表明,MeJA 或JA 处理能够提高水稻抗虫性和抗机械损伤等,但对水稻的产量则具有抑制作用; 因此,如何合理喷施MeJA 提高水稻抗性并尽量降低其对水稻产量的影响,是农业生产实践中值得进一步深入探索的问题。
综上所述,基因的表达受bHLH009 转录因子的转录调控; 稗草胁迫下,‘PI312777’中基因上调表达且高于‘Lemont’,体现了‘Lemont’和‘PI312777’对稗草胁迫信号响应的灵敏性差异。bHLH009 是JA 信号途径的调节蛋白,外源添加MeJA 能够提高基因的表达,MYB57、MAPK11 和PAL2;3 蛋白的表达水平也提高,有利于促进水稻根系分泌酚酸类化感物质,提高水稻对稗草的化感抑制作用; 其中添加0.05 mmol∙LMeJA 不会抑制水稻生长,且有效提高水稻化感抑草能力。上述结果揭示了稗草胁迫下,化感水稻通过JA 途径调控MYB57 表达,促进水稻合成酚酸类化感物质抑制稗草生长的转录调控过程与机制。
猜你喜欢
云南化工(2020年11期)2021-01-14 00:50:52
文苑(2020年12期)2020-04-13 00:55:10
意林·全彩Color(2019年11期)2019-12-30 06:09:06
意林彩版(2019年11期)2019-11-22 11:49:05
现代园艺(2017年23期)2018-01-18 06:57:44
新疆农垦科技(2016年10期)2016-06-15 20:29:33
戏剧之家(2016年1期)2016-02-25 11:15:20
杂草学报(2015年2期)2016-01-04 14:57:58
化工进展(2015年3期)2015-11-11 09:07:41
中国当代医药(2015年10期)2015-03-01 02:02:39
