
桃Pp4CL2基因的克隆及抗寒功能验证
2022-02-13 10:06李小兰郝兰兰
西北植物学报 2022年12期
李小兰,郝兰兰,张 帆,王 鸿*
(1 甘肃省农业科学院 林果花卉研究所, 兰州 730070;2 甘肃农业大学 园艺学院,兰州 730070)
苯丙烷生物合成途径是植物主要的次级代谢途径,在植物中普遍存在[1]。由苯丙氨酸作为起始底物,经苯丙氨酸脱氢酶(phenylalanin ammonia-lyase, PAL)、肉桂酸-4-羟基化酶(cinnamate-4-hydroxylase, C4H)以及4-香豆酸辅酶A连接酶(4-coumarate CoA ligase, 4CL)催化生成4-香豆酸辅酶A后,进入不同的分支途径合成类黄酮、木质素、色素类等不同的多酚类次生代谢物。木质素作为细胞壁的重要组成部分,在植物应对生物和非生物胁迫中提供机械支持,其含量与植物应对环境胁迫的抗性成正比[2-3]。在植物处于逆境时,应激合成的类黄酮可以清除植物体内由于代谢紊乱超量积累的氧自由基,维持细胞稳态[4]。这些重要的次生代谢物是植物应对环境胁迫的重要途径之一[5-6],在植物生长发育以及逆境胁迫的应答过程中发挥重要作用[7-8]。
4-香豆酸辅酶A连接酶(4CL)是植物苯丙烷生物合成途径中的最后一个关键酶,处于苯丙烷代谢途径的终端位置。大多数植物中4CL基因是以基因家族的形式出现的,如拟南芥中有3个4CL基因(At4CL1、At4CL2、At4CL3)[9],大豆(Gm4CL1、Gm4CL2、Gm4CL3、Gm4CL4)[10]和杂种杨中均有4个4CL基因(Ptd4CL1、Ptd4CL2、Ptd4CL3、Ptd4CL4)[11]。不同家族成员在植物中的作用也不尽相同[12],4CL基因在植物中的作用主要分为两类:第Ⅰ类参与类黄酮的生物合成,第Ⅱ类参与木质素的生物合成。Cao等[13]通过对库尔勒香梨4CL基因家族的研究发现,只有同时具有保守结构域LPYSSGTTGLPK和催化活化中心GEICIRG的4CL基因才能够调控木质素的合成,而不同时具有这两种保守结构域的家族基因参与黄酮类物质的合成。
目前,关于4CL基因的研究大多集中在模式作物中,在桃中尚没有关于4CL基因的研究报道。低温是影响桃树生长发育和地理分布的重要环境因子。本研究采用RT-PCR方法从‘丁家坝李光桃’中克隆了一个4CL(Pp4CL2)基因,通过农杆菌介导法获得转Pp4CL2的拟南芥和烟草,并对其抗寒功能进行初步分析,以期为桃抗寒新品种的培育提供候选基因。
1 材料和方法
1.1 试验材料
本研究材料为抗寒型桃品种‘丁家坝李光桃’,取自甘肃省农业科学院桃园。转基因材料使用红花大茎子烟草(NicotianatabacumL.)组培无菌苗和哥伦比亚生态型拟南芥(Arabidopsisthalianaecotype Columbia),均为本实验室已有材料。
1.2 试验方法
1.2.1 RNA提取及Pp4CL2基因克隆称取桃叶片0.1 g,参照植物总RNA提取试剂盒(Tiangen,北京)操作说明提取总RNA,以提取的总RNA为模板合成cDNA第一链。从课题组前期获得的‘丁家坝李光桃’抗寒转录组数据中筛选Pp4CL2候选基因的转录本,并通过NCBI进行比对,获得Pp4CL2基因全长CDS序列。使用DNAMAN软件设计特异性引物(表1),以获得的cDNA为模板进行PCR扩增。反应程序为:94 ℃预变性5 min;94 ℃变性30 s,58 ℃退火30 s,72 ℃延伸90 s,32个循环;72 ℃延伸10 min。PCR产物用2%琼脂糖凝胶进行电泳并回收目的条带,连接pMD19-T克隆载体,转化大肠杆菌并挑选阳性克隆进行测序。
1.2.2Pp4CL2生物信息学分析理化性质利用在线网站ExPASy(http://web.expasy.org/protparam/)进行分析;采用SOPMA(https://npsa-prabi.ibcp.fr/cgibin/npsa_automat.pl?page=npsa_sopma.html)对编码蛋白的二级结构进行预测;运用SMART在线网站(http://smart.emblheidelberg.de/)预测蛋白结构域;亚细胞定位使用Plant-mPLoc(http://www.csbio.sjtu.edu.cn/bioinf/plant-multi/)进行预测,从NCBI数据库中(https://blast.ncbi.nlm.nih.gov)搜索并下载同源氨基酸序列,运用DNAMAN软件进行同源氨基酸序列比对分析;通过MEGA 7.0软件,采用Neighbor-Joining法建立系统树。
1.2.3 植物表达载体构建及农杆菌转化参照曹冬梅等[14]的方法进行表达载体的构建。将测序鉴定的重组质粒pMD19-T-Pp4CL2和载体pRI101用KpnⅠ和SmaⅠ进行双酶切反应,在37 ℃的条件下反应1~1.5 h后用2%琼脂糖凝胶电泳检测条带大小并切胶回收。连接目的基因片段与载体,转化大肠杆菌。采用冻融法进行农杆菌LBA4404转化,在28 ℃恒温培养箱倒置暗培养48 h左右,挑选单克隆在LB液体培养基上培养,菌液做PCR鉴定。
1.2.4 转基因烟草的获得及鉴定参照郭爱霞[15]的方法进行农杆菌介导的烟草遗传转化。将烟草组培苗叶片减去主脉及叶柄,剪成0.5 cm×0.5 cm大小的小方块,浸泡在无菌水中备用;将叶片夹入已活化好的农杆菌箘液中振荡侵染20 min;取出叶片用无菌滤纸吸干,叶片正面朝上平铺于MS培养基上;黑暗预培养3 d后,将叶片转移到含250 mg/mL头孢霉素和100 mg/mL卡那霉素的烟草分化培养基上。培养条件为温度25℃,光照强度6000 Lx,光周期16 h/8 h。当芽长到约1 cm左右时,剪下继代到生根培养基,每14 d继代1次,60 d左右获得T1代转基因植株。依据试剂盒说明书(Tiangen 北京)提取转基因烟草叶片的DNA,用实时荧光定量PCR进行检测。
1.2.5 转基因拟南芥的获得参照Hu等[16]的方法通过花序浸染法进行拟南芥的遗传转化。得到T0代拟南芥种子,用75%乙醇处理5 min,2.6%次氯酸钠处理10 min,去离子水冲洗3次后点播到含卡那霉素的培养基上,用PCR筛选出抗性植株。经过连续3代的筛选,获得T3代纯合的转基因植株。
1.2.6 实时荧光定量PCR设计Pp4CL2基因特异引物(表1),以Actin为内参基因[17],不同时间低温处理的cDNA为模板,依据SYBR Premix Ex TaqTM Ⅱ试剂盒(TaKaRa,大连)说明书,使用Light Cycler®96 Instrument PCR仪(Roche, 瑞士)进行PCR反应。反应体系为20 μL:2×SYBRRPremixExTaqTMⅡ 10 μL,10 μmol/L上/下游引物各1 μL,cDNA模板2 μL,ddH2O 6 μL。试验结束后,采用2-ΔΔCT法计算基因相对表达量,并用SPSS软件进行差异显著性分析。
1.2.7 转基因烟草和拟南芥的低温处理和相关指标测定将野生型拟南芥和T3代纯合种经过消毒和春化处理,播种到MS培养基上,15 d后将幼苗转移到基质中进行培养。基质中生长1个月以后同野生型烟草和获得的T1代转基因烟草一同转移到培养箱进行4 ℃低温诱导,分别在低温处理0和12 h时,选取一组材料采集叶片,立即用液氮速冻,-80 ℃保存备用。采用硫代巴比妥酸(TBA)反应[18]测定丙二醛含量(MDA);磺基水杨酸—酸性茚三酮法测定[19]游离脯氨酸含量(Pro);考马斯亮蓝G-250染色法测定可溶性蛋白含量[20];蒽酮比色法测定可溶性糖含量[21];使用Yun等描述的方法测定相对电导率[22];过氧化物酶(POD)和超氧化物歧化酶(SOD)含量使用索莱宝POD和SOD试剂盒,根据说明书进行测定。数据采用Excel工作表和SPSS 20.0进行整理分析。
2 结果与分析
2.1 Pp4CL2基因克隆及表达载体的构建
以‘丁家坝李光桃’叶片的cDNA为模板,利用特异引物(表1)扩增出1 500 bp左右的目的片段(图1, A),将纯化回收的目的片段连接到pMD19-T克隆载体,挑选阳性克隆进行测序后发现,该目的片段与桃基因组数据库中的4CL2序列片段完全一致,大小为1 635 bp,为桃的Pp4CL2基因(登录号:LOC18792923)。使用ExPASy在线软件分析Pp4CL2基因的理化性质可知:Pp4CL2基因编码544个氨基酸残基;分子式为C2696H4292N708O788S22;相对分子质量为59.94 kD;理论等电点为5.83;正负电荷数分别为58、68。不稳定系数为30.32,为稳定类蛋白;脂肪系数为99.65;亲水性为-0.009,属于疏水性蛋白。在NCBI网站对其特定结构域进行分析,发现其含有4CL基因家族保守结构域。

表1 引物序列
对测序鉴定的含Pp4CL2的pMD19-T-Pp4CL2重组质粒和原核表达载体pRI101用KpnⅠ和SmaⅠ进行双酶切反应,将含有Pp4CL2编码区的1 635 bp片段插入表达载体pRI101,成功构建过表达载体pRI101-Pp4CL2(图1, B)。
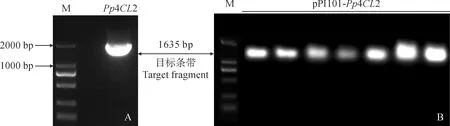
图1 Pp4CL2(A)和pRI101-Pp4CL2(B)的PCR扩增结果Fig.1 The PCR amplification results of Pp4CL2(A) and pRI101-Pp4CL2(B)
2.2 Pp4CL2生物信息学分析
通过在线软件对Pp4CL2蛋白二级结构进行预测。结果显示(图2):Pp4CL2蛋白由4种状态的二级结构组成,其中α螺旋占30.51%、β-折叠占7.35%、不规则卷曲及延伸链分别占41.54%和20.59%。利用New PLACE网站预测Pp4CL2前2000 bp序列中的顺式作用元件。结果表明(表2):该序列含有许多与抗逆性相关的顺式作用元件,如:低温响应元件(LTR)、光响应元件(GA-motif)和水杨酸响应元件(TCA-element)等。说明该基因可以对外界多种信号做出响应,参与植物的生长发育和器官形成过程。

蓝色. α螺旋;紫色. 不规则卷曲;红色. 延伸链;绿色. β-折叠图2 Pp4CL2蛋白二级结构预测Blue. Alpha helix; Purple. Random coil;Red. Extended strand; Green. Beta turnFig.2 Secondary structure of Pp4CL2

表2 Pp4CL2顺式作用元件分析
通过NCBI对Pp4CL2蛋白序列进行同源性检索并通过DNAMAN软件对同源序列进行比对。结果表明(图3),Pp4CL2与杏(Prunusarmeniaca)、欧洲甜樱桃(Prunusavium)和梅(Prunusmume)的相似性最高,分别为99.08%、97.98%和96.14%,与其他物种的4CL2蛋白在C端有较高的保守性,而在N端具有一定的差异,说明4CL2基因在进化过程中具有一定的保守性。为了进一步分析Pp4CL2与检索到的其他物种4CL2的亲缘关系,通过MEGA 5.0构建系统发育树。结果显示(图4):桃与杏(Prunusarmeniaca,KAH0993361.1)、梅(Prunusmume,XP_008223734.1)和欧洲甜樱桃(Prunusavium,XP_021810058.1)聚在一个亚家族,亲缘关系最近,与其他物种亲缘关系较远。

图3 4CL2蛋白多序列比对Fig.3 Multi-sequence alignment of 4CL2 protein

图4 Pp4CL2及同源蛋白进化分析Fig.4 Phylogenetic analysis of Pp4CL2 and homologous proteins
2.3 转Pp4CL2基因烟草和拟南芥的获得
将成功构建的pRI101-Pp4CL2过量表达载体通过冻融法转化农杆菌LBA4404。采用农杆菌菌液侵染烟草叶盘并置于黑暗预培养3 d,随后将叶片转移到含头孢霉素和卡那霉素的烟草分化培养基上进行培养。叶片分化产生不定芽后,剪下继代到生根培养基,培养60 d左右时对其进行阳性鉴定,获得转基因烟草(图5)。

A.农杆菌侵染烟草叶盘; B.阳性不定芽产生; C.阳性不定芽培养; D.植株再生;图5 Pp4CL2基因遗传转化烟草A. Agrobacterium infect leaf disc of tobacco; B. Generation of positive adventitious buds; C. Culture of positive adventitious buds; D. Plant regenerationFig.5 Genetic transformation of Pp4CL2 gene into tobacco
采用农杆菌菌液侵染拟南芥花序,获得T0代拟南芥种子。对T0代种子进行消毒后点播在含卡那霉素的培养板上,待15 d左右移栽到基质中置于光照培养箱中进行培养并进行PCR阳性鉴定。经过连续3代的筛选,获得T3代纯合的转基因拟南芥(图6)。

A.农杆菌侵染拟南芥花序; B.拟南芥种子点播;C.抗性拟南芥培养;D.抗性拟南芥移栽图6 Pp4CL2基因遗传转化拟南芥A. Agrobacterium infect inflorescence of Arabidopsis; B. Arabidopsis seed on-demand; C. Resistance A. thaliana culture; D. Resistance A. thaliana transplantFig.6 Genetic transformation of Pp4CL2 gene into A. thaliana
2.4 转基因拟南芥和烟草对低温胁迫的响应
选取长势良好的野生型烟草(WT)、拟南芥(WT),以及转基因烟草(#1、#2、#3)、拟南芥(#1、#2、#3)置于4 ℃培养箱处理0和12 h。通过qRT-PCR测定Pp4CL2基因在不同时间4 ℃低温处理下不同株系中的相对表达量。结果(图7)表明:4 ℃处理0和12 h后,拟南芥(图7, A)和烟草(图7, B)的3个转基因株系中Pp4CL2基因的表达量均显著高于其野生型的表达量,且处理12 h与处理0 h的表达量相比,转基因株系的表达量上调较野生型株系明显。不同的转基因株系在相同时间的低温处理下,Pp4CL2基因的表达量变化差异不大。
2.5 低温胁迫下转基因拟南芥的表型及生理指标测定
选择长势相近的野生型和转基因株系的一月生拟南芥幼苗,置于4 ℃培养箱中冷训化24 h后降到-4 ℃处理4 h,观察表型。结果(图8, A)表明:不同株系拟南芥在4 ℃处理4 h之后与处理之前(25 ℃)对比均出现了不同程度的失水萎蔫现象,野生型株系相较于转基因株系失水萎蔫程度更严重。

图8 -4 ℃处理下拟南芥(A)和烟草(B)表型Fig.8 Phenotype of A. thaliana (A) and tobacco (B) treated at -4 ℃
将野生型拟南芥株系和转基因拟南芥株系经4 ℃处理0 和12 h后,取叶片测定其低温胁迫下的生理生化指标。结果(图9)表明:4 ℃低温处理0 h时,转基因株系和野生型株系的相对电导率和丙二醛含量差异不大。4 ℃低温处理12 h后,转基因株系中的相对电导率和丙二醛含量显著低于野生型株系(图9, A~B),说明转基因拟南芥在低温胁迫下受损伤程度较轻,抗寒能力较强。转基因植株体内的渗透调节物质经4 ℃处理12 h以后,含量显著高于野生型株系(P<0.05),说明转基因能增加植物脯氨酸、可溶性蛋白和可溶性糖含量的积累(图9, C~E)。4 ℃处理下转基因株系的过氧化物酶和超氧化物歧化酶的活性显著高于野生型株系,说明低温胁迫下转基因株系的抗氧化能力高,抗寒性强(图9, F、G)。

图9 4 ℃低温胁迫下野生型拟南芥和转Pp4CL2拟南芥的生理生化指标Fig.9 Physiological and biochemical indicators of wild and transgenic Arabidopsis thaliana under 4 ℃ low temperature stress
2.6 低温胁迫下转基因烟草的表型及生理指标测定
选择长势相近的野生型烟草和过表达Pp4CL2基因的烟草组培苗置于4 ℃培养箱中进行冷训化24 h后降到-4 ℃处理2 h,观察表型(图8, B)。结果表明:-4 ℃处理2 h后野生型烟草出现了严重的失水现象,叶片萎蔫卷曲。转基因烟草相比于野生型烟草受害程度较轻,说明转基因烟草比野生型烟草对低温更具有耐受性。
将野生型和转基因烟草株系置于培养箱中进行4 ℃低温处理,处理0和12 h后进行叶片取样并测定生理生化指标,探究其低温胁迫下的生理生化变化(图10)。结果表明:与野生型株系相比,转基因株系在4 ℃低温胁迫下的相对电导率和丙二醛含量较低,说明过表达Pp4CL2株系的细胞结构不易被破坏,抗寒能力强(图10, A、B)。4 ℃低温处理0 h时的脯氨酸、可溶性蛋白和可溶性糖等渗透调节物质的含量差异不大,但经4 ℃胁迫12 h后转基因烟草的含量显著高于野生型烟草,说明转基因可以增加植株的脯氨酸、可溶性蛋白和可溶性糖的含量(图10, C~E)。转基因烟草在4 ℃低温胁迫12 h时,过氧化物酶活性和超氧化物歧化酶活性显著高于野生型烟草,说明转基因烟草的抗氧化能力高于野生型烟草(图10, F、G)。
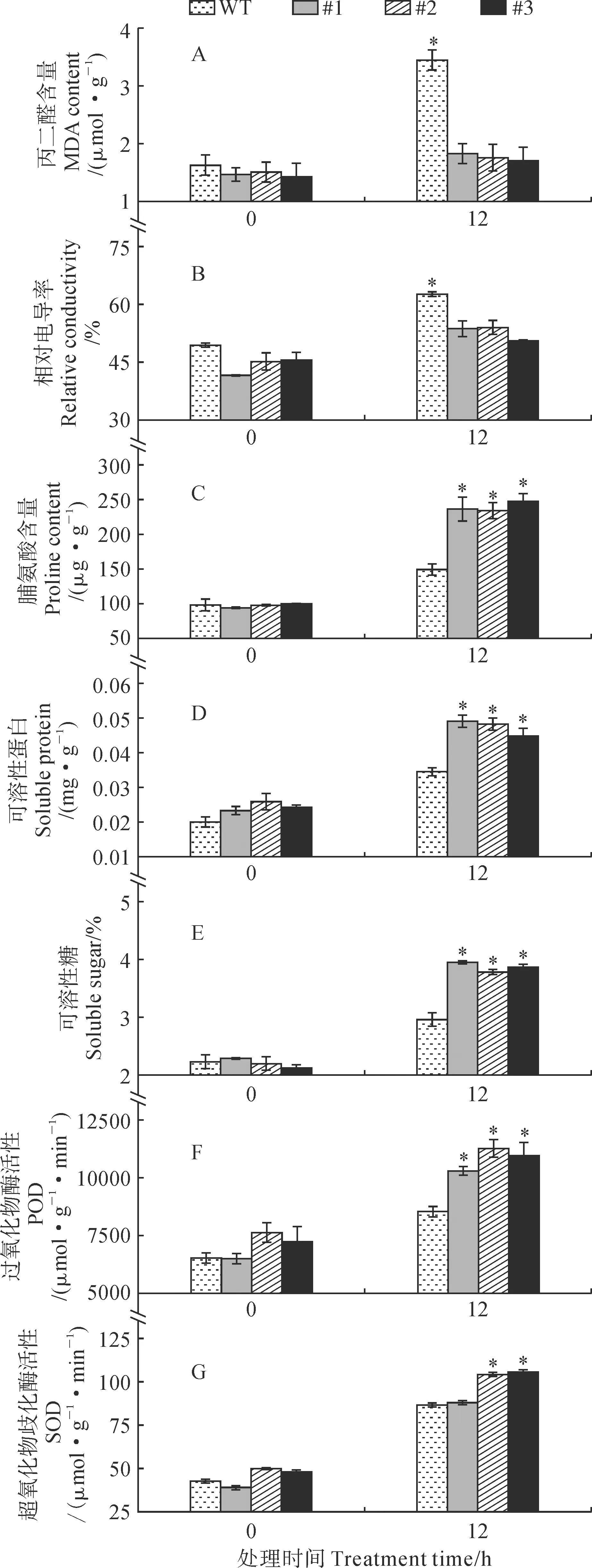
图10 4 ℃低温胁迫下烟草的生理生化指标Fig.10 Physiological and biochemical indicators of tobacco under 4 ℃ low temperature stress
3 讨 论
近年来,针对4CL基因家族的研究已成为热点。自1987年,Lozoya等[23]首次从欧芹(Petroselinumcrispum)中克隆出2个4CL基因以来,水稻(Oryzasativa)[24]、拟南芥(Arabidopsisthaliana)[25]、烟草(Nicotianatabacum)[26]和毛白杨(populustomentosaAcarr)[27]等约40多种植物的4CL基因被相继克隆出来,但在桃中尚没有关于4CL基因研究与克隆的报道。本研究根据前期获得的桃响应低温胁迫的转录组数据,结合RT-PCR技术克隆得到一个桃4CL转录因子家族的4CL2基因,命名为Pp4CL2。序列分析表明:Pp4CL2编码544个氨基酸残基,与其他物种4CL基因的氨基酸序列具有较高的一致性,显示了Pp4CL2在进化过程中的保守性。保守基序分析表明:Pp4CL2含有4CL家族保守结构域中的LPYSSGTTGLPK和GEICIRG序列,属于第Ⅱ类4CL基因,参与植物体中木质素的生物合成。系统进化分析表明:Pp4CL2与杏的4CL基因亲缘关系最近,同源性最高。顺式作用元件分析表明:Pp4CL2前2 000 bp序列中含有许多与抗逆性相关的顺式作用元件,说明该基因可以响应植物体受到的外界刺激,参与植物的生长发育以及逆境应答过程。
前人对不同物种中4CL基因的克隆及功能验证方面的研究很多,但大都集中在4CL基因对不同物种生长发育方面[28-30]的作用研究,鲜有关于4CL基因在植物逆境胁迫方面响应的研究。前人通过对干旱和低温胁迫下云烟203幼苗[31]和银杏叶片中[32]的多酚物质含量及其代谢相关酶活性的研究表明,4CL酶活受低温诱导。但其仅仅关注低温胁迫下的酶活,并没有对编码蛋白的4CL基因进行研究和功能验证。低温是影响桃树生长发育以及地理分布的关键因素,为了进一步研究Pp4CL2在桃低温响应过程中的功能,我们构建了Pp4CL2基因的过表达载体,并运用农杆菌介导法遗传转化拟南芥和烟草,通过对低温处理下转Pp4CL2基因型和野生型拟南芥和烟草的相对表达量、表型和生理生化指标的测定表明,低温胁迫下转基因拟南芥和烟草与野生型拟南芥和烟草相比Pp4CL2的相对表达量高,受冷害程度轻,具有更高的渗透调节物质含量和抗氧化酶活性,对低温具有更强的耐受性。
猜你喜欢
学与玩(2022年10期)2022-11-23
今日农业(2022年3期)2022-06-05
亚热带植物科学(2022年1期)2022-05-17
四川农业科技(2019年5期)2019-07-01
上海农业学报(2017年3期)2017-04-10
浙江柑橘(2016年1期)2016-03-11
创新科技(2015年1期)2015-12-24
红领巾·探索(2015年9期)2015-09-10
天然产物研究与开发(2014年6期)2014-04-27
植物营养与肥料学报(2014年1期)2014-03-11
