
储存温度对单瓣和重瓣紫斑牡丹花粉活力及生理特性的影响
2022-02-13 09:00:16刘兴宇何丽霞
西北植物学报 2022年12期
赵 萍,唐 红*,刘兴宇,何丽霞
(1 甘肃农业大学 林学院,兰州 730070;2 甘肃农业大学 信息科学技术学院,兰州 730070;3 甘肃省牡丹工程研究中心,兰州 730046)
紫斑牡丹(Paeoniarockii)为芍药科芍药属落叶小灌木,是中国重要的观赏植物与油料作物之一,同时还具有抗寒、抗旱、耐瘠薄等特性[1]。紫斑牡丹花香,花瓣基部具紫斑,长势强盛,抗旱、抗寒、抗病、耐土壤瘠薄,具有很高的观赏及育种价值[2]。在甘肃主要分布于甘肃南部、定西、兰州等地。‘象牙白’花淡黄白色,长势强,花香,结实品质良好,是临夏、临洮等地传统品种[1]。
花粉粒的体积很小,具有很强的耐干燥性,特别适合储存,是种质资源保存的重要材料之一;杂交是牡丹培育新优品种的主要途径,特别是远距离杂交和远缘杂交,需要解决时空不遇和延长花粉寿命等问题,以便与合适的母体进行杂交[3-5]。牡丹花粉的储存和萌发率的测定有助于制定适宜的杂交方案,盖伟玲等[6]通过研究4个牡丹品种的花粉在固体培养基萌发的情况,认为培养6 h左右是计算牡丹花粉萌发率的适宜时期。紫斑牡丹单朵花寿命极短,有研究表明,单朵花从开放到衰老仅有7~10 d,因其开花时间的短暂性,花粉寿命也极为短暂,对杂交育种极为不利[7-9]。筛选适合‘象牙白’花粉储存的适宜方案是延长花粉寿命,解决远距离杂交的唯一途径。此外,重瓣牡丹的结实性能更为低下,对其育种和油用价值产生了很大的影响,制约了牡丹产业的进一步发展[6]。近年来,越来越多的研究发现,由于温度和水分的胁迫,花粉萌发率及寿命与花粉贮存期间活性氧的清除机制及膜脂过氧化水平有关,花粉保护酶活性、丙二醛和蛋白质含量保持稳定是花粉保持高活力的生理特征[10]。保护酶活性、MDA和蛋白质含量的高低可以反映花粉的健康程度,与花粉寿命及生活力具有密切关系,是花粉萌发力的直接表现[5,11]。目前,有关紫斑牡丹的研究主要集中在引种栽培[12]、遗传学[7]、油用性能[13]等方面。关于其单瓣与重瓣花花粉结构、萌发特性和储存生理机制方面的系统研究尚未见报道。因此,本研究以甘肃省兰州市榆中县官滩沟紫斑牡丹培育基地紫斑牡丹‘象牙白’花粉为材料,观测了单瓣和重瓣紫斑牡丹花粉的形态结构、不同储存条件下的寿命,及其储存期间3种保护酶活性、MDA和蛋白质的含量,以期确定‘象牙白’花粉经济、最佳的储存温度,探讨单瓣和重瓣‘象牙白’花粉寿命与花粉保护酶活性、MDA以及蛋白质含量的相关性,解析其花粉寿命在不同储存环境下降低的生理机制,为紫斑牡丹重瓣不育性和杂交育种提供理论和实验依据。
1 材料和方法
1.1 花粉采集
紫斑牡丹‘象牙白’花粉采自甘肃省兰州市榆中县官滩沟紫斑牡丹培育基地,于 2021年6月中上旬9:00-11:00,采集同一株牡丹植株上同时具有单瓣与重瓣的将开未开的紫斑牡丹花朵,按单重瓣分别标号,迅速放入冰盒,带回实验室。在实验室中将采回花朵的花萼与外轮花瓣剥去,倒扣在硫酸纸上,置于恒温培养箱内散粉,24 h后取出,一只手用镊子夹住倒置的花朵,另一只手轻弹花朵,收集散落的花粉。将收集好的花粉每个编号各分成两份,一份用于花粉萌发的测定;另一份 30 ℃ 鼓风干燥箱干燥 24 h 后,分装至 10 mL 带盖离心管中储存,写好标签,备用。
1.2 花粉储存
将1.1中所得干燥花粉分成7份,每份分装于2 mL带盖离心管中,加入1~2粒硅胶干燥剂后封口,分别存放于室温(25 ℃)、4 ℃、-20 ℃、-80 ℃下储存。从储存之日算起,分别于7、21、60、120、190 d后,分别从4个储存温度处理中取单瓣和重瓣花粉各1份(-20、-80 ℃下取出的花粉于35 ℃水浴5~10 min解冻),取出一部分花粉采用前期实验所得最适培养基[14]测定花粉萌发率,统计花粉储存寿命,另一部分用于测定生理生化指标。
1.3 观测指标及方法
1.3.1 花粉超微结构将完全干燥的花粉均匀撒在贴有双面胶的样品台上,在扫描电镜下观察拍照,记录花粉粒极轴长(P)、赤道长(E)、萌发沟长、萌发沟宽、脊宽度和网眼直径,每个样品随机测量10个以上,并计算平均值。
1.3.2 花粉萌发率和花粉寿命花粉萌发培养基由蔗糖、硼酸和琼脂配制而成。用移液管吸取所需用量加入容量瓶中,加蒸馏水定容后加热至沸腾,冷却至 30 ℃。用吸管吸取培养液,滴在载玻片上,待其凝固后,蘸取花粉均匀播种在液滴上,将播种有花粉的载玻片放在铺有吸水纸的培养皿中,置于恒温培养箱中培养,在(25±1) ℃ 、全光照下培养 6 h 后于显微镜下观察。取 4 个视野,数取花粉总数及萌发数,计算萌发率。统计时,以花粉管长度等于或大于花粉直径者为萌发。每个储存处理设 3 个重复。
花粉萌发率=萌发花粉数/观察花粉数×100%。
1.3.3 花粉可溶性蛋白质、丙二醛含量及保护酶活性准确称取0.100 g‘象牙白’花粉,放入4 ℃预冷研钵中,加入 2 mL 0.05 mol/L 磷酸缓冲液(pH 7.0),匀速研磨 30~40 s,之后将研磨液转移至10 mL离心管中,定容至刻度线。将提取液于10 000 r/min、4 ℃离心 20 min,取出上清液作为提取粗酶液,保存至4 ℃冰箱备用待测。可溶性蛋白含量采用考马斯亮蓝法、过氧化氢酶(CAT)活性采用可见光分光光度法、丙二醛(MDA)含量采用硫代巴比妥酸法、超氧化物歧化酶(SOD)活性采用氮蓝四唑法、过氧化物酶(POD)活性采用愈创木酚法,通过天津市光复科技发展有限公司提供的试剂盒测定。
1.4 数据分析
试验数据用Excel 2010、SPSS 24和Origin 2021进行作图与方差分析。
2 结果与分析
2.1 单瓣和重瓣花形态与花粉表观特征比较
紫斑牡丹‘象牙白’花粉粒为长椭圆形,两端逐渐收缩呈截形;花粉粒的3个萌发沟基本到达两端,极面观为近圆形或三角形,网眼直径大小不等,多为椭圆形、近圆形或不规则形。其中,单瓣‘象牙白’花瓣白色,花粉数多(图 1,A、B);花粉粒有3个萌发沟,是孢粉学研究的原始类型;花粉粒平均极轴长为50.98 μm(47.07~57.34 μm),平均赤道轴长为23.51 μm(21.73~26.69 μm),极轴长(P)/赤道轴长(E)≈2.18,萌发沟长44.60 μm(40.10~ 50.16 μm),萌发沟宽1.24 μm(0.71~2.19 μm),网眼直径1.39 μm(0.99~2.04 μm),脊宽度0.68 μm(0.45~0.81 μm)(图 1,C-F)。重瓣‘象牙白’花瓣为白色,花粉较少,部分雄蕊瓣化为花瓣(图 1,G、H)。花粉粒有3个萌发沟,萌发沟几乎达到两级,是孢粉学研究的原始类型;花粉粒平均极轴长为43.79 μm(39.63~48.00 μm),平均赤道轴长为23.42 μm(19.31~24.83 μm),P/E≈1.88,萌发沟长为44.60 μm(31.80~45.45 μm),萌发沟宽1.24 μm(0.87~3.10 μm),网眼直径1.39 μm(0.96~1.71 μm),脊宽度0.68 μm(0.51~0.92 μm)(图 1,I-L)。另外,‘象牙白’畸形花粉长在 10~40 μm 之间,总体观不饱满、空瘪,多为勺状、三棱状、不规则状等,单瓣和重瓣‘象牙白’畸形花粉分别约占观察花粉总数的 20.50%和 35.40%。

A-F.单瓣花;G-L.重瓣花;A、G花形态;B、H.花粉群体;C、I.赤道面观;D、J.萌发沟;E、K.极面观;F、L.萌发沟细部图1 单瓣和重瓣‘象牙白’花形态与花粉表观特征A-F. Single flower; G-L. Double flower; A,G. The flower morphology; B,H. Pollen grain; C,I. Polar morphology; D,J. Germinal furrow; E,K. Equatorial morphology; F,L. Fine parts of germinating furrowFig.1 Flower characteristics and pollen grain morphology of single-petal and double-petal ‘Xiangyabai’
2.2 储存温度对单、重瓣花花粉寿命的影响
图2显示,新鲜紫斑牡丹‘象牙白’花粉萌发率表现为单瓣花高于重瓣花,单瓣和重瓣新鲜花粉萌发率分别为83.62%和72.06%。随着储存时间的延长,单瓣和重瓣花花粉萌发率均逐渐下降,且总体呈现先快后慢的趋势。随着储存温度的降低,单瓣和重瓣花花粉萌发率均逐渐增加,且单瓣花在温度处理间的差异更大。其中,在室温条件下,花粉萌发率下降最快,这可能是较高温度下呼吸作用较强,消耗了较多的营养,导致花粉很快丧失了萌发力。在储存第21天前,花粉萌发率下降极为迅速,之后下降速度减缓;在第120天,单瓣和重瓣花花粉萌发率均降为0。即室温条件下单瓣和重瓣花花粉寿命为120 d。在4 ℃储存条件下,花粉萌发率下降呈现先快后慢的趋势,第60天为转折点,单瓣花和重瓣花花粉的半衰期分别为40和60 d,半衰期前花粉活力下降较为迅速,之后逐渐减缓。在 -20 ℃储存条件下,花粉萌发率下降呈现先缓后快的趋势,第21天为分界点;单瓣和重瓣花花粉半衰期均为120 d。在-80 ℃储存条件下,花粉萌发率下降速度最慢,单瓣花和重瓣花花粉萌发率在储存190 d后分别为76.39%和65.17%,分别降低为最开始的96.17%和90.44%。可见,储存温度极大地影响着紫斑牡丹花粉萌发率,低温有助于延长花粉的储存寿命,4种储存温度中-80 ℃最适合花粉的中长期储存,单瓣花花粉对低温的适应性高于重瓣。
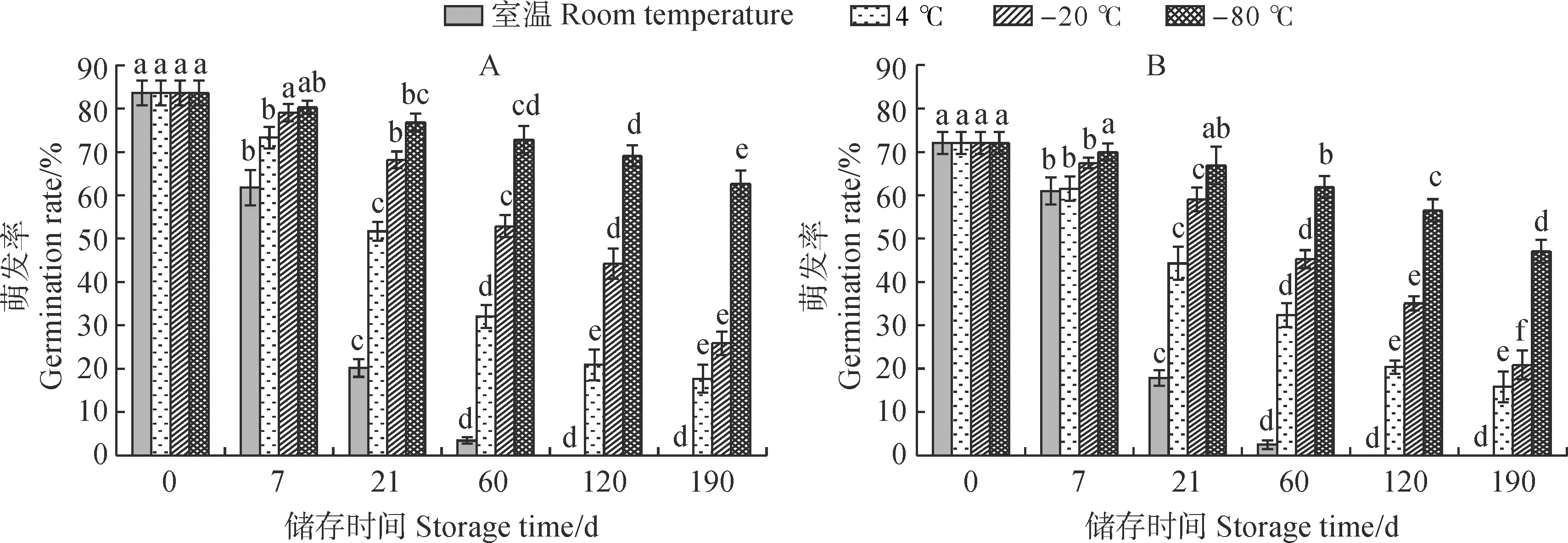
不同小写字母表示同一温度不同储存时间0.05水平差异显著性,下同图2 不同储存温度下单瓣(A)和重瓣(B)‘象牙白’花粉萌发率的变化The different normal letters mean significant difference at 0.05 level among different storage time at the same temperature, the same as belowFig.2 Pollen germination rate of single-petal (A) and double-petal (B) ‘Xiangyabai’ under different storage temperatures
2.3 储存温度对单、重瓣花花粉可溶性蛋白含量的影响
从图 3 可以看出,新鲜单瓣花花粉可溶性蛋白含量高于相应的重瓣花花粉。各温度处理下单瓣和重瓣花花粉可溶性蛋白含量均先下降后增加,并多在第21天达到最低值,之后随着储存时间的延长,花粉可溶性蛋白含量逐渐增加。在储存190 d时,单瓣和重瓣花花粉可溶性蛋白含量均随着储存温度的降低而下降。其中,单瓣和重瓣花花粉蛋白质含量在-80 ℃储存条件下均较为稳定,相较于其他温度条件波动幅度较小;在室温条件下,花粉可溶性蛋白含量变化幅度最大。单瓣花花粉可溶性蛋白含量增速在室温条件下最快,在4 ℃条件下较快,在第190天时分别已增加为初始值的3.59和1.75倍;而在-20 ℃和-80 ℃条件下增速较慢,第190天时仍低于初始值。重瓣花花粉可溶性蛋白含量在室温条件下增速最快,在4 ℃条件下增速较快,于第190天时达到最高值,分别增加为初始值的3.31和2.60倍;在-20 ℃条件下,花粉可溶性蛋白含量于第60天时最高,为初始值的3.13倍;-80 ℃条件下,可溶性蛋白含量增速最慢,第190天时为初始值的1.63倍。由此可见,单瓣花花粉可溶性蛋白含量变化在4 ℃、-20 ℃和-80 ℃条件下趋势较重瓣花更为稳定,说明单瓣花花粉受低温胁迫影响的程度更小;但在常温条件下,重瓣花花粉可溶性蛋白含量变化趋势比单瓣花花粉更为稳定。
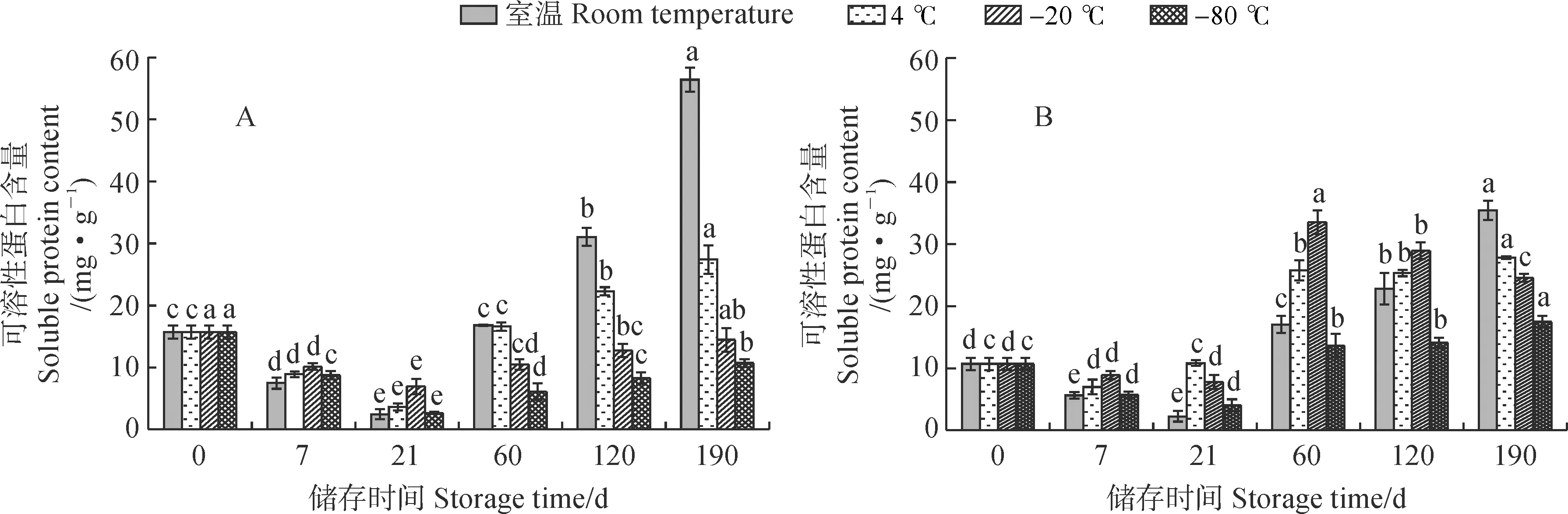
图3 不同储存温度下单瓣(A)和重瓣(B)‘象牙白’花粉可溶性蛋白含量的变化Fig.3 Soluble protein contents in pollen of single-petal (A) and double-petal (B) ‘Xiangyabai’ under different storage temperatures
2.4 不同储存条件对单重瓣花花粉 MDA 含量的影响
从图 4 可知,新鲜紫斑牡丹‘象牙白’单瓣花花粉MDA含量低于重瓣花花粉;低温储存条件下,单瓣和重瓣花花粉MDA含量均随储存时间先增加后减少,但重瓣花花粉受低温胁迫的影响较单瓣花花粉更小。其中,单瓣和重瓣花花粉MDA含量在室温条件、-20 ℃和-80 ℃下均于储存第21天达到最高峰,此时分别为初始值5.36倍和5.18倍、4倍和3.89倍、3倍和3.27倍。单瓣和重瓣花花粉MDA含量在4 ℃条件下均于第60天时达到最大,此时分别为初始值的4倍和5.06倍。单瓣和重瓣花花粉MDA含量在室温条件下变化最为显著,相较而言均在-80 ℃条件下较为稳定。
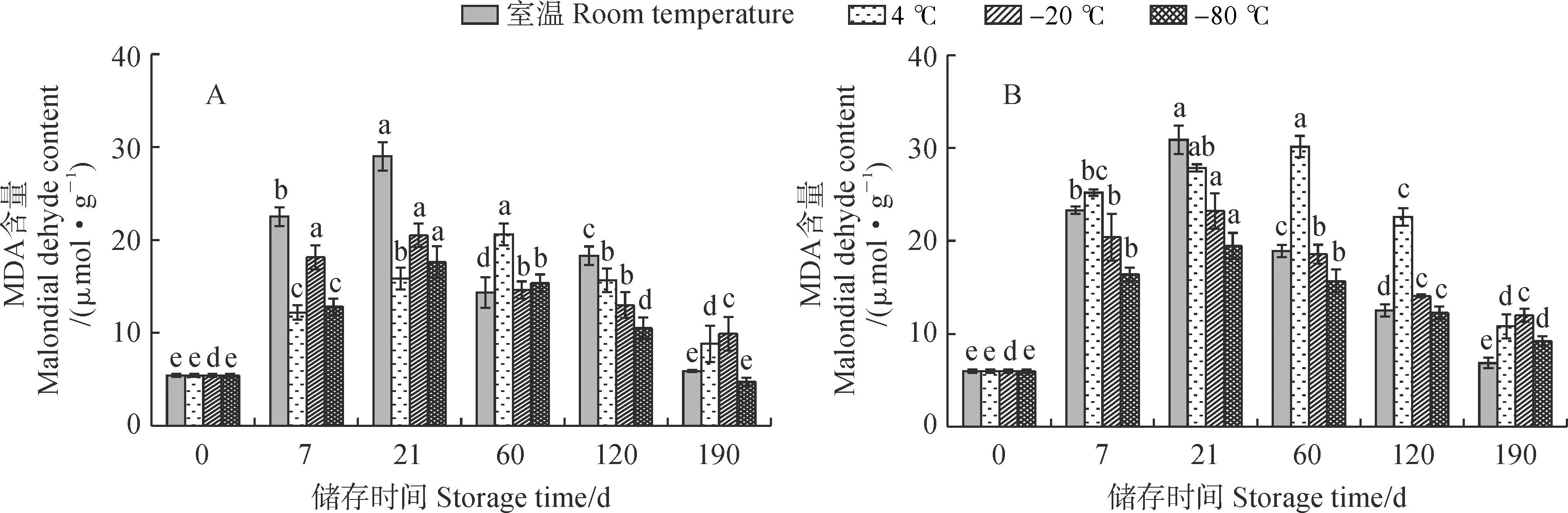
图4 不同储存温度下单瓣(A)和重瓣(B)‘象牙白’花粉MDA含量的变化Fig.4 Malondial delyde content in pollen of single-petal (A) and double-petal (B) ‘Xiangyabai’ under different storage temperatures
2.5 储存温度对单、重瓣花花粉3种保护酶活性的影响
2.5.1 SOD 活性图 5 显示,新鲜紫斑牡丹‘象牙白’单瓣花花粉SOD活性低于重瓣花花粉。在室温条件下,单瓣和重瓣‘象牙白’花粉SOD活性随着储存时间均呈现逐渐快速下降的趋势,在第120天(花粉萌发率为0)时分别降为最高值的19.80%和16.39%;相较之下,重瓣花花粉SOD活性下降较单瓣花花粉更为迅速,单瓣花和重瓣花花粉SOD活性在190 d时分别为初始值的4.9%和2.1%。在4 ℃和-20 ℃条件下,单瓣花和重瓣花花粉SOD活性均先增加后减少,在第21天时达到最高峰,这可能与花粉萌发率的急速下降有关。在-80 ℃条件下,单瓣与重瓣花花粉SOD活性均呈现先缓慢增加后逐渐下降的趋势,总体变化趋势相较平稳,在190 d后均保持在较高水平,而此时的花粉萌发率也相对较高。总体来看,重瓣花花粉SOD活性受低温胁迫的影响大于单瓣花花粉。

图5 不同储存温度下单瓣(A)和重瓣(B)‘象牙白’花粉SOD活性的变化Fig.5 Superoxide dismutase activity in pollen of single-petal (A) and double-petal (B) ‘Xiangyabai’ under different storage temperatures
2.5.2 POD活性从图 6可知,新鲜紫斑牡丹‘象牙白’单瓣花花粉POD活性低于重瓣花花粉。单瓣和重瓣‘象牙白’花粉POD活性在不同储存温度下随着时间的变化趋势不同,并在室温条件下变化趋势最显著,在-80 ℃条件下变化趋势最为平缓;花粉POD活性在室温、4 ℃和-20 ℃条件下均表现为先增加后降低,均在第21天达到最高值,在-80 ℃条件下先降低后增加,在第60天达到最低值。其中,第190天时,单瓣和重瓣花花粉POD活性在室温、4 ℃和-20 ℃条件下分别为最高值的13.62%和17.99%、23.91%和33.30%、40.43%和54.98%;单瓣和重瓣花花粉POD活性在-80 ℃储存第60天时达到最低值,此时分别降为最高值的38.13%和62.30%。可见,各温度条件下重瓣花花粉POD活性随着储存时间的变化较单瓣花花粉更为平缓,其受低温胁迫的影响较单瓣花花粉更小。
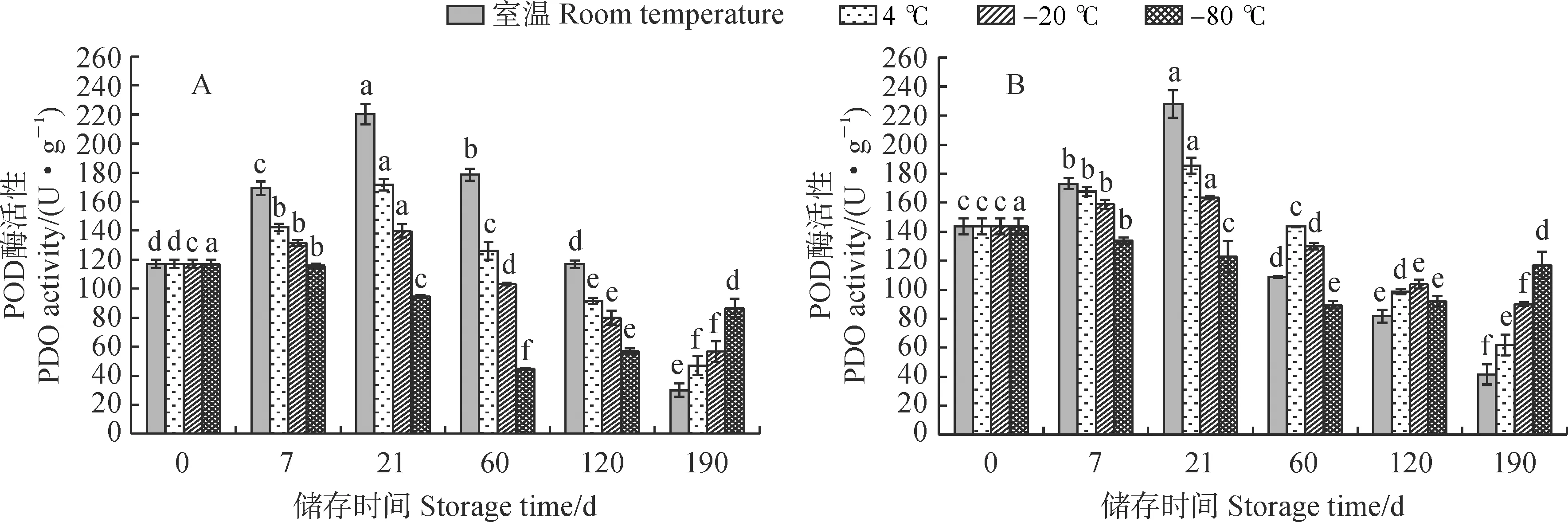
图6 不同储存温度下单瓣(A)和重瓣(B)‘象牙白’花粉POD活性的变化Fig.6 Peroxidase activity in pollen of single-petal (A) and double-petal (B) ‘Xiangyabai’ under different storage temperatures
2.5.3 CAT活性从图 7 来看,新鲜紫斑牡丹‘象牙白’单瓣花花粉CAT活性低于重瓣花花粉。各温度下单瓣和重瓣花花粉CAT活性随着储存时间的变化趋势总体相近,但不同温度下的CAT活性存在显著性差异。其中,单瓣和重瓣花花粉CAT活性均在-80 ℃条件下变化幅度最小,在第60天时分别降为最高值的60.24%和70.51%;均在室温条件下变化幅度最大,单瓣和重瓣花花粉最高值分别为最低值的5.09和4.14倍。重瓣花花粉CAT活性在低温储存过程中相较于单瓣花花粉更为稳定。单瓣花花粉CAT活性在室温、-20 ℃和-80 ℃条件下均先降低后增加,均在第60天达到最低值,而4 ℃条件下呈现增加-降低-增加的趋势。重瓣花花粉CAT活性在室温、4 ℃、-20 ℃条件下均呈现先下降后增加的趋势,在-80 ℃条件下为持续下降趋势。可见,在4种温度储存条件下,两种花粉CAT活性变化幅度均在室温条件下最大,均在-80 ℃条件下更为稳定,且单瓣花花粉CAT活性受低温胁迫的影响较重瓣更为明显。
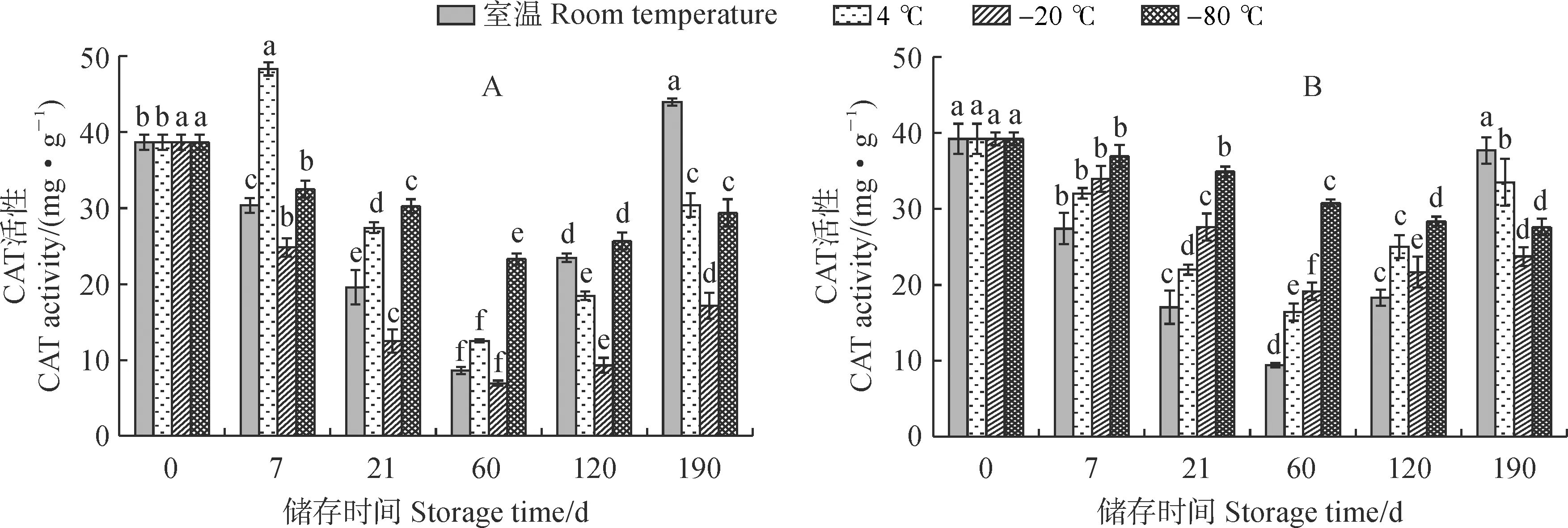
图7 不同储存温度下单瓣(A)和重瓣(B)‘象牙白’花粉CAT活性的变化Fig.7 Pollen catalase activity in pollen of single-petal (A) and double-petal (B) ‘Xiangyabai’ under different storage temperatures
3 讨 论
3.1 单瓣和重瓣花粉萌发率与储存温度的关系
室温条件下的环境温度较高且湿度较大,花粉的呼吸作用以及生理代谢活动较强,导致花粉细胞失水严重,从而使花粉的生活力降低,最终使花粉的储藏寿命缩短。而低温干燥环境条件能够抑制花粉的代谢活动,降低酶活性和呼吸作用,从而减缓花粉生活力下降的速度。也有研究认为低温、低湿以及低氧的环境能够延长花粉的储藏寿命[15]。低温储存是生物材料长期保存最有前途的方法,已经广泛应用于作物、水果、蔬菜和观赏植物[16]。花粉因体积很小而比较适用于储存,单瓣花花粉与重瓣花花粉的储存时间也不尽相同,且花粉寿命在自然条件下仅有9 d左右,适宜的储存条件可以有效延长花粉寿命,为日常杂交育种提供较高活力的花粉,满足育种工作的各种需求,因此筛选适宜的花粉储存条件具有重要的意义。研究表明,芍药属植物花粉寿命的长短受多方面因素的制约,一是基因型的控制,基因型的差异严重影响花粉的寿命;二是花粉含水量,低含水量是延长花粉寿命的有效措施[17-18]。除此之外,储存环境对于花粉的寿命也尤为重要,低温、干燥和低氧的环境可有效延长花粉寿命。本实验结果表明,经过不同储存温度处理的紫斑牡丹花粉萌发率较新鲜花粉均有不同程度的降低,室温储存的花粉寿命仅有120 d,4 ℃和-20 ℃只适用于花粉的短期保存,而-80 ℃条件下储存190 d的花粉仍有较高活性和萌发率,适用于花粉的长期保存。而单瓣与重瓣花花粉的寿命也不同,单瓣花花粉对低温的适应性高于重瓣花花粉。
3.2 花粉萌发率与其可溶性蛋白质含量的关系
蛋白质是花药发育过程中积累的营养物质,其作为细胞组成中重要的代谢物质和能源物质,对于花药和小孢子的正常发育都极为重要[19]。曾乃燕等[20]对水稻研究的结果认为低温下可溶性蛋白含量的增加,可能是由于降解速率下降或合成的加强。可溶性蛋白含量是重要的渗透调节物质,一定量的可溶性蛋白能提高细胞的保水能力,从而起到保护细胞膜的作用。本研究中紫斑牡丹单瓣和重瓣花花粉在储存第21天可溶性蛋白含量达到最低,这可能与花粉前期的活力快速下降有关,储存21 d后可溶性蛋白含量逐渐增加,温度越高增加越快,这可能与花粉对不同温度胁迫下产生了抗逆蛋白有关,抗逆蛋白一旦在植物体内形成,植物体就会尽快适应外界环境,表现出很强的抗逆性[21]。紫斑牡丹单瓣花花粉在4 ℃、-20 ℃和-80 ℃条件下受低温胁迫的程度比重瓣花更小,重瓣花花粉在常温条件下受到的胁迫程度比单瓣花更小,这可能与两类花粉本身的遗传基因有关。
3.3 花粉萌发率与丙二醛含量和抗氧化酶活性的关系
MDA是膜脂过氧化的一个重要指标,在花粉发育时期的大量积累会使细胞膜结构遭到破坏或损伤, 从而引起细胞内生理生化代谢的紊乱。MDA含量的高低代表细胞质膜透性的强度,与其是否受到伤害具有密切的相关性[22]。本研究中紫斑牡丹单瓣和重瓣花花粉MDA含量均在各温度下储存第21天之前快速上升,在第21天达到高峰,而花粉活力也在同一时间段快速下降,这说明花粉细胞膜受到严重的破坏或损伤,膜脂过氧化程度较高[23]。花粉MDA含量在室温条件下变化程度最为显著,说明在高温高湿条件下细胞受到破坏程度更大,而花粉MDA含量变化在-80 ℃储存时较为平和,花粉萌发率下降速度极慢,可能是花粉细胞内保护酶的协同作用有效地清除了多余的活性氧,重新达到氧化平衡。同时,植物花粉在利用氧的过程中会形成活性氧,这些活性氧会对植物本身产生毒害作用,从而会发生体内相关组织的生理生化代谢紊乱的现象,SOD、POD、CAT等抗氧化酶可以帮助清除植物体内积累的过剩有氧自由基,防止活性氧过度积累而造成膜质受损。研究表明,保护酶活性一定程度上是生物体机能维持的重要指标[24-25]。生物材料贮存过程中形成的活性氧引起的氧化应激是造成细胞损伤,甚至诱导细胞凋亡的主要原因之一,也可能是不同贮存环境花粉寿命差异明显的原因之一[26]。保护酶活性的稳定与花粉保存质量具有相关性,保护酶活性的大幅增加或降低可能预示着植物材料萌发率的下降[27],在本研究花粉萌发率下降最快期间,花粉细胞膜质过氧化加剧,活性氧、自由基增多,但随着时间延长、胁迫加剧,3 种保护酶不足以消除细胞内积累的活性氧,导致有害物质积累,花粉细胞死亡,花粉萌发率随之下降。-80 ℃储存条件下,紫斑牡丹花粉3种保护酶活性在不同时间阶段均保持较为平稳,花粉萌发率接近新鲜花粉萌发率,这说明-80 ℃储存环境使花粉基本处于代谢平衡状态,健康程度高,因此花粉仍然具有较高的萌发力。
4 结 论
紫斑牡丹‘象牙白’重瓣花花粉平均极轴长、平均赤道轴长、极轴长/赤道轴长均小于单瓣花花粉,其萌发沟长和宽、脊宽度和网眼直径与单瓣花花粉区别不大。紫斑牡丹花粉小粒、瘪粒和畸形是导致其萌发率较低的主要原因,不同生存环境下花粉遗传特性存在差异,仅花粉表观不足以揭示其系统演化规律。储存温度直接影响紫斑牡丹‘象牙白’花粉寿命,室温适合单瓣和重瓣花花粉1~120 d的短期储存,4 ℃、-20 ℃适合花粉较长时间的储存,-80 ℃适合花粉的长期储存。在室温、4 ℃和-20 ℃储存期间,单瓣和重瓣花粉可溶性蛋白含量显著降低后再增高,抗逆蛋白的形成是花粉萌发率下降速率减缓的主要原因;花粉MDA含量和3种抗氧化酶SOD、POD、CAT活性随着储存时间显著变化,清除活性氧自由基能力下降,细胞膜脂过氧化程度加剧,细胞损伤加重是花粉萌发率在储存期间下降的主要原因。花粉在-80 ℃储存条件下基本处于代谢平衡状态,健康程度较高,在长期储存后仍然具有较高的萌发力。重瓣花花粉受到低温胁迫的影响程度高于单瓣花花粉,在相同储存条件下相较于单瓣花花粉,重瓣花花粉细胞受损程度更高、萌发率下降速率更快。
猜你喜欢
食品与生物技术学报(2021年8期)2021-01-17 01:38:07
北京园林(2020年2期)2020-01-18 03:30:44
现代园艺(2018年1期)2018-03-15 07:56:12
现代园艺·综合版(2017年1期)2017-07-14 16:33:50
现代园艺(2017年1期)2017-07-14 04:19:32
农村百事通(2016年7期)2016-05-14 14:11:39
阅读与作文(初中版)(2016年5期)2016-05-14 12:00:55
作文评点报·中考版(2015年46期)2015-12-04 03:05:06
思维与智慧·上半月(2015年8期)2015-08-07 17:11:10
福建农业科技(2015年3期)2015-02-27 10:21:01
