
黑杨派杨树种质资源的SSR 分析及鉴定
2022-02-11 07:46蔄胜军苏晓华彭儒胜吴建军
西南林业大学学报 2022年1期
刘 巍 蔄胜军 苏晓华 彭儒胜 吴建军
(1. 辽宁省杨树研究所,辽宁 营口 115213;2. 中国林业科学研究院林业研究所,北京 100091;3. 辽宁省实验林场,辽宁 抚顺 113309)
黑杨派(SectionAigeiros)杨树是杨属中最为速生的树种,为高大落叶乔木,具有极高的经济价值和遗传学价值,全世界使用的绝大多数栽培品种均源于黑杨派[1-2]。黑杨派天然种质资源存在丰富的遗传变异,是杨树育种研究重要的物质基础[3]。从20 世纪70 年代开始,我国开始有计划地引进黑杨派种质资源并对其进行研究,这些丰富的种质资源为我国杨树新品种培育奠定了种质基础,使我国的杨树育种研究得到了迅速的发展,改变了我国原有杨树栽培品种的格局[4]。
传统的形态学鉴定方法不能系统地对种质资源进行遗传学评价,分子标记技术不受组织、发育时期、季节环境限制,为杨树育种研究提供了新的手段。另外,杨树同一类的不同品种(系)间形态特征差异很小,单纯以形态学差异很难进行区分,给杨树新品种的鉴定和知识产权保护增加了困难,对林业生产影响很大,在良种的推广应用上会造成一定困难。为保证品种的特异性,利用分子标记技术开展遗传分析并构建指纹图谱信息已逐渐成为品种鉴定的重要手段及良种审定的重要信息。在众多分子标记技术中,SSR 标记具有多态性丰富、重复性好、共显性等特点,是遗传多样性研究和品种鉴定中较为理想的分子标记技术[5-7],在杨树种质资源遗传多样性研究[8-9]、品种鉴定[10-11]、指纹图谱构建[12-13]等方面广泛应用。本研究采用毛细管电泳检测技术对辽宁地区引种和自主选育的50 份黑杨派种质资源进行SSR 遗传分析,依据相似系数构建聚类图并对种质进行遗传关系分析,同时对种质的纯度进行初步鉴定,并对辽宁地区主栽的14 份种质构建指纹图谱,为杨树育种群体筛选、分类及杂交育种工作提供参考,同时为杨树新品种的鉴定和知识产权保护提供参考依据。
1 材料与方法
1.1 材料
供试的50 份黑杨派种质材料(表1)来自辽宁省杨树研究所黑杨派基因库和种质资源保存圃,包括美洲黑杨31 份,欧洲黑杨7 份,欧美杨12 份。
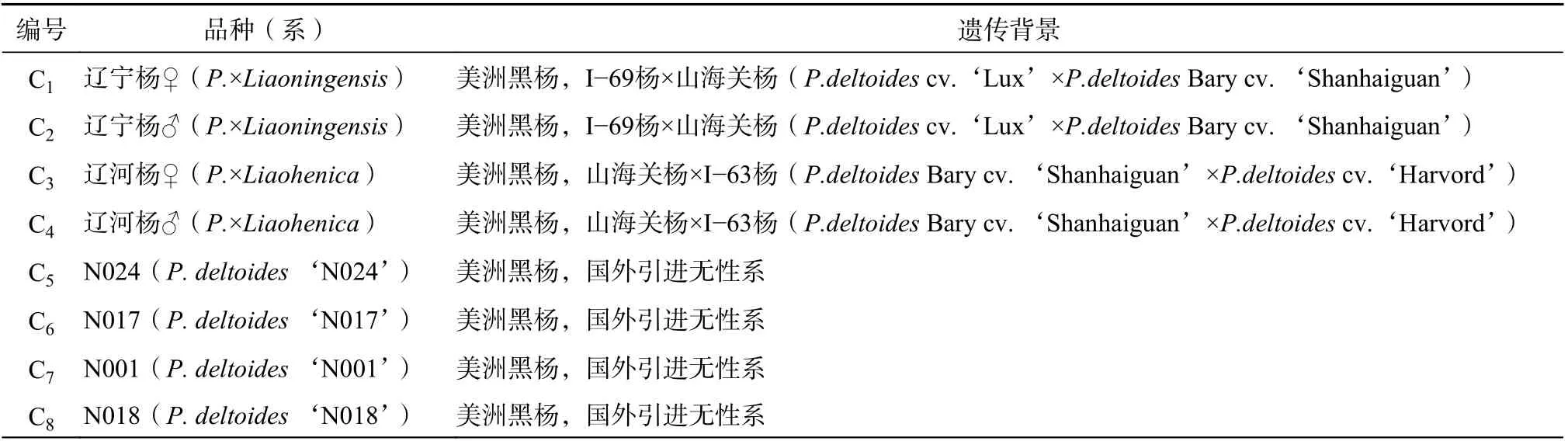
表1 试验材料Table 1 Experimental materials

续表1
1.2 方法
1.2.1 DNA 提取
2020 年春季采集大树枝条,于温室水培后采集嫩叶,采用CATB 法提取DNA[14]。
1.2.2 引物筛选
从李薇等[7]和丁明明[15]所报道的黑杨派杨树引物中选取23 对SSR 引物用于分析,引物由北京鼎国昌盛生物技术有限责任公司合成。分别从3 类杨树种质材料中各选取2 份样品通过聚丙烯酰胺凝胶电泳法进行引物筛选,最后筛选出扩增条带清晰、具有多态性和重复性好的15 对引物(表2)进行毛细管电泳检测。

表2 15 对SSR 引物信息表Table 2 Information of 15 pairs of SSR primers
1.2.3 PCR 扩增与毛细管电泳检测
采用15 对SSR 引物进行PCR 扩增,PCR 产物的毛细管电泳检测由北京鼎国昌盛生物技术有限责任公司完成。SSR-PCR 反应体系25 μL,包括模板DNA(20 ng/μL)2 μL,正、反向引物(10 μmol/L)各0.5 μL,dNTP(10 μmol/L)0.5 μL,10×PCR buffer 2.5 μL,Taq酶(5 U/μL)0.5 μL 和ddH2O18.5 μL。PCR 反应程序:95 ℃预变性5 min;95 ℃变性30 s,50 ℃退火30 s,72 ℃延伸30 s,35 个循环;最后72 ℃延伸10 min;4 ℃保存。扩增反应在Gene Amp PCR System 9600 型扩增仪上进行。采用ABI3730X Genetic Analyzer对扩增片段进行毛细管电泳检测。
1.2.4 数据分析
采用DateFormater 软件将电泳检测结果转换为“1,0”矩阵,运用POPGENE32 软件进行数据分析,计算等位基因数(Na)、有效等位基因数(Ne)、期望杂合度(He)、观测杂合度(Ho)等遗传多样性指标。用Excel 2007 计算每对引物的多态信息含量(PIC)。运用NTSYS 2.1 软件将“1,0”矩阵转换为ntsys 格式,采用软件中的SM 相似系数程序计算任意2 个品种间的相似系数,采用UPGMA 方法进行聚类分析并绘制聚类图。
2 结果与分析
2.1 SSR 引物多态性分析
利用15 对SSR 引物对参试50 份样品的基因组DNA 进行扩增,3 类黑杨派杨树种质的部分扩增结果见图1。由表3 可知,15 对引物共检测到187 个等位位点,其中多态位点187 个,多态性位点百分率为100%。每对引物检测到多态位点数8~19 个,平均12.47 个,其中引物P3 最多,为19 个,引物GCPM-2826 和引物P139 次之,为16 个,引物GCPM-2140 和引物P133 最少,为8 个。有效等位基因数(Ne)平均1.184 5 个。期望杂合度(He)为0.652 5~0.920 4,平均0.820 7,观测杂合度(Ho)为0.321 5~0.983 2,平均0.739 1。多态信息含量(PIC)为0.63~0.91,平均0.79。以上参数表明,50 份杨树种质具有丰富的遗传多样性,且根据Botstein[16]提出的理论,所筛选的15 对引物都具有高度多态性(PIC>0.5)。
图1 引物GCPM-2885 对部分杨树种质的电泳图谱Fig. 1 Electropherogram detected by GCPM-2885 for parts of poplar germplasm

表3 15 对引物在50 份杨树种质中的多态性Table 3 Polymorphisms of 15 pairs of SSR primers in 50 poplar germplasm

续表3
从分析结果可知,15 对SSR 引物中有12 对引物在17 份样品中检测到了特异性位点(表4),共检测到特异性位点36 个,每个引物检测到特异性位点数量为1~5 个。引物GCPM-2885、GCPM-2140 和P133 没有检测到特异性位点,引物P139检测到特异性位点最多,为5 个。36 个特异性位点分布于3 类杨树种质中。其中,N024、N018和N51 分别在5 对引物下都检测到特异性位点,N017 分别在3 对引物下都检测到了特异性位点。这充分表明了杨树基因组DNA 的多态性,说明了不同参试材料之间的遗传差异。

表4 SSR 引物特异性位点Table 4 Specific sites of SSR primers
2.2 50 份杨树种质的聚类分析
以50 份杨树种质的遗传相似系数为依据,采用UPGMA 法进行聚类分析,得到参试样品的遗传关系聚类分析图(图2)。50 份杨树种质相似系数为0.727 3~0.989 3,平均0.804 5,表明参试种质间存在较高的遗传相似度。聚类结果显示,当相似性系数在0.79 时,参试样品可分为4 大类。
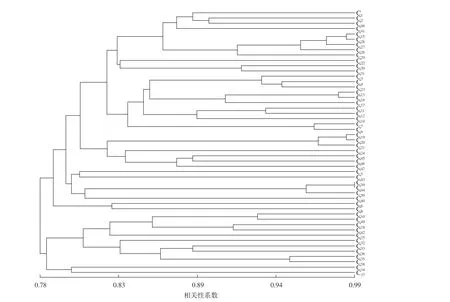
图2 基于SSR 结果的50 份杨树种质的聚类结果Fig. 2 Clustering results of 50 poplar germplasm based on SSR results
第Ⅰ大类包括36 份种质,主要以美洲黑杨类种质为主,其中5030、180-39、180-33 和180-28聚为1 个小类,相似系数为0.951 9~0.984 0;5028和2017 聚为1 个小类,相似系数达到0.978 6;N001 和N027 聚为1 个小类,相似系数达到0.962 6;178-2-19 、178-2-122 和178-2-145 聚为1 个小类,相似系数为0.962 6~0.984 0。以上几个小类主要与其均为美洲黑杨及引自相同种源有关。辽宁杨♀、辽宁杨♂和盖杨聚为1 个小类,相似系数为0.877 0~0.893 0,与其为同一父本杂交子代有关;辽河杨♀、辽河杨♂和辽育3 号杨聚为1 个小类,相似系数为0.930 5~0.941 2,这可能与其都具有山海关杨基因有关。111 杨、辽丰1 号杨和014 杨之间相似系数较大,为0.951 9~0.989 3,这3 份种质均为欧美杨遗传背景,另外,这3 个品种(系)外部形态及其相似,类似108 杨,很难区分,说明其亲缘关系较近,遗传差异较小。渤丰1 号杨、渤丰2 号杨和渤丰3 号杨聚为1 个小类,相似系数为0.866 3~0.882 4,与其为同一杂交组合选育的新品种有关。第Ⅱ大类包括N017 和N018,相似系数为0.828 9,这与其引自同1 种源有关。第Ⅲ大类包括10 份种质,主要以欧洲黑杨类种质为主,其中,N009 与中荷64 杨聚为1 个小类,相似系数为0.925 1;YN 与中辽1 号杨聚为1 个小类,相似系数为0.909 1;N13 和N54 聚为1 个小类,相似系数为0.946 5,这2 份种质均引自欧洲种源,说明其亲缘关系较近。第Ⅳ大类包括N4 和N51,相似系数为0.802 1,与其共同引自欧洲种源有关。
欧美杨是美洲黑杨与欧洲黑杨的杂交种,12 份欧美杨种质中有10 份聚到第Ⅰ大类中,包括盖杨、辽育2 号杨、渤丰1 号杨、渤丰2 号杨、渤丰3 号杨、欧美杨177、111 杨、014 杨、辽丰1 号杨和I-45 杨,说明这10 份种质与美洲黑杨亲缘关系较近;中辽1 号杨和中荷64 杨聚到第Ⅲ大类中,说明这2 份种质与欧洲黑杨的亲缘关系较近。
2.3 14 份主栽种质指纹图谱的构建
以辽宁省目前生产上主栽的速生的14 份种质为研究对象,构建指纹图谱。依据15 对引物对14 份种质的扩增结果,综合考虑多态性和引物扩增位点数,最少可以利用鉴别效率较高的4 对引物组合GCPM-2826、P139、P3 和P141 构建指纹图谱,该指纹图谱中,14 份种质均可应用特有位点或位点组合加以鉴定。
由表5 可知,引物GCPM-2826 可以区分辽宁杨♀、辽宁杨♂、辽育2 号杨、渤丰2 号杨、渤丰3 号杨、盖杨和中荷64 杨共7 份种质;引物P139 可以区分除I-45 杨、中辽1 号杨、欧美杨177 和渤丰1 号杨共4 份种质;引物P3 和引物P141 可以区分111 杨、辽丰1 号杨和014 杨共3 份种质。通过4 对高效引物即可将14 份种质完全区分开。

表5 14 份主栽种质的指纹图谱Table 5 Fingerprint map of 14 main planted germplasm
3 结论与讨论
毛细管电泳检测技术具有操作简便、成本低、污染少、精准度高等优点,与传统的聚丙烯酰胺凝胶电泳银染技术自动化和程序化更高,可以减少试验过程中人为和系统误差,尤其适合于大量样本的高通量检测[17]。王世杰等[18]用11 对SSR 引物对50 份杨树新品种进行扩增,并使用毛细管电泳检测技术对扩增结果进行检测,11 对引物共扩增出122 个DNA 片段,扩增的等位重复序列数为4~17 个,平均每对引物11.091 个,不同引物PIC值的变化范围是0.530~0.908,平均为0.803,同时对50 份样品进行了聚类分析,证实了SSR 分子标记可以有效的鉴别杨树品种,并能很好的反映品种间的亲缘关系。贾会霞等[12]采用GeXP 毛细管电泳对PCR 荧光产物进行检测,19对SSR 引物共扩增出102 条条带,多态性条带占95.10%,每个位点的等位基因数为2~11 个,平均每对引物5.37 个,并为24 份杨树种质构建了指纹图谱。本试验利用15 对SSR 引物对黑杨派3 个类别的50 份种质进行扩增,采用ABI3730 测序仪对扩增片段进行毛细管电泳检测,共检测到多态位点187 个,多态性位点百分率为100%,平均每对引物检测到多态位点数为12.47 个,平均多态信息含量为0.79,说明筛选的15 对引物具有高度的多态性。观测杂合度是随机抽取的2 个样本的等位基因不相同的概率,能够反映等位基因的丰富度的高低,杂合度越高,该位点处的基因型越多,遗传变异越丰富[19]。本试验中平均期望杂合度和平均观测杂合度分别为0.820 7 和0.739 1,相较于前人的研究具有较高的基因杂合性[19-21],表明了50 份杨树种质具有较高的遗传变异性,是遗传基础较为丰富的育种群体。
本研究中,有12 对引物在多个样品中检测到了特异性位点,共检测到特异性位点36 个,说明不同样品之间遗传差异较大。其中,N024、N018和N51 分别在5 对引物下都检测到了特异性位点,N017 分别在3 对引物下都检测到了特异性位点。通过聚类分析,可以较好地表现各种质的亲缘关系。研究表明,50 份杨树种质相似系数在0.727 3~0.989 3 之间,平均0.804 5,表明参试种质间存在较高的遗传相似性,这与参试样品均为黑杨类遗传背景有关。试验中辽宁杨♀、辽宁杨♂和盖杨聚为1 个小类,辽河杨♀、辽河杨♂和辽育3 号杨聚为1 个小类,渤丰1 号杨、渤丰2 号杨和渤丰3 号杨聚为1 个小类,以上几组聚类结果基本符合种质的系谱关系;111 杨、辽丰1 号杨和014 杨3 份种质外部形态特征及其相似,因此聚为1 个小类,说明了遗传本质差异对外部形态特征的对应反映[22]。通过试验也证实了所选的15 对SSR 引物在可以有效地对50 份杨树种质进行遗传多样性分析、种质鉴别的同时,也可以较好地研究种质间亲缘关系,为杨树种质的分类研究和核心育种群体的筛选工作提供分子基础。本研究的试验材料中,美洲黑杨和欧洲黑杨的遗传背景是根据材料来源及形态特征对其进行的初步分类,可能会存在个别种质分类不准确,而综合试验中特异性位点的检测和聚类分析的结果,初步判断31 份美洲黑杨种质中的N024、N018、N017、N009、YN 和47#可能不是纯种美洲黑杨,7 份欧洲黑杨种质中的N4 和N51 可能不是纯种欧洲黑杨,还有待于进一步验证。
目前生产上主栽的杨树新品种(系)遗传基础较狭窄,且生物学性状受生长发育阶段和环境影响较大,单纯以形态特征的差异作为品种区分标准比较困难,而且不利于新品种权的保护[12,23]。本研究采用4 对SSR 引物构建了14 份辽宁地区主栽杨树种质的指纹图谱,能够对14 份种质进行有效的鉴定,为杨树新品种的鉴定和知识产权保护提供参考依据。
对北方型美洲黑杨和欧洲黑杨种质资源遗传多样性分析、种质纯度鉴定和遗传关系进行分析,是本研究的重要研究内容。在此基础上,对种质资源进行分类,下一步将结合表型特征评价初步筛选核心育种群体,利用黑杨派树种的速生、干直、抗病虫的特性,对辽宁省乃至东北地区的乡土杨树进行遗传改良,选育速生、抗逆性强的杨树新品种,解决目前生产上栽培品种林分稳定性差和缺乏更新换代品种的问题。
猜你喜欢
分子催化(2022年1期)2022-11-02
中国农业科学(2022年16期)2022-09-19
今日农业(2022年14期)2022-09-15
今日农业(2022年13期)2022-09-15
广西植物(2022年8期)2022-09-07
中国农学通报(2022年12期)2022-06-01
作物学报(2022年4期)2022-02-10
中国种业(2022年1期)2022-01-27
中国种业(2022年1期)2022-01-27
作物学报(2022年3期)2022-01-22
