
月季TIFY基因家族的全基因组分析
2022-02-07 02:25王昊宁周佩刘燕刘梦朱晓仪石力匀栗燕逯久幸
河南农业大学学报 2022年6期
王昊宁, 周佩, 刘燕, 刘梦, 朱晓仪, 石力匀, 栗燕, 逯久幸
(1.河南农业大学风景园林与艺术学院,河南 郑州 450002;2.河南省优质花卉蔬菜种苗工程技术中心,河南 郑州 450002)
月季(RosachinensisJacq.)是全球重要的观赏花卉,具有极高的经济价值。环境方面的非生物胁迫问题限制了月季的生长发育,严重影响其花卉品质[1]。研究月季中的非生物胁迫响应基因将有助于遗传改良其胁迫耐受性,应对日益严峻的环境挑战。
TIFY蛋白含有保守的TIFY结构域,TIFY基因家族根据具体包含的额外结构域类型分为4个亚家族:TIFY亚家族仅包含TIFY结构域;PPD亚家族除TIFY结构域外,还含独特的N端PPD结构域;JAZ家族除TIFY结构域外,还包含1个CCT_2结构域;ZIM亚家族在JAZ亚家族的基础上,还包含1个ZnF_GATA结构域[2]。最近的研究表明,TIFY蛋白在调节植物干旱胁迫和盐胁迫的抗性中发挥着重要的作用[3-4]。拟南芥AtTIFY10a和AtTIFY10b以及其野生大豆的同源基因GsTIFY10a和OsTIFY10b,均为调节盐碱胁迫响应的正向因子[5]。拟南芥AtTIFY5b过表达赋予拟南芥耐旱性[6]。水稻中大多数TIFY蛋白至少参与1种非生物胁迫的响应,其中包含耐旱与耐盐,OsTIFY11a的过表达可以增强水稻对盐胁迫和脱水胁迫的耐受性[7]。棉花GaJAZ5基因的过表达减少了水分流失、气孔开口和过氧化氢的积累,提高植物的耐旱性[8]。过表达小麦TdTIFY11a的拟南芥幼苗的发芽率和生长率均高于野生型,表现出较高的盐胁迫耐受性[9]。
综上,TIFY家族的分析对月季干旱胁迫和盐胁迫的调控分子机制的研究具有重要意义。本研究通过对月季基因组的扫描,获取月季TIFY家族成员序列,对月季TIFY家族成员进行了系统进化分析、理化性质分析、基因结构分析、组织表达模式分析以及顺式元件分析。为进一步了解TIFY基因在月季干旱胁迫和盐胁迫的调控作用奠定了基础,并为月季遗传改良胁迫耐受性提供理论依据。
1 材料与方法
1.1 月季TIFY家族成员分析所需数据来源
月季基因组序列、蛋白序列库、CDS序列库来源于https://lipm-browsers.toulouse.inra.fr/pub/RchiOBHm-V2/。拟南芥TIFY家族成员蛋白序列来源于https://www.arabidopsis.org/index.jsp。水稻TIFY家族成员蛋白序列来源于https://www.ncbi.nlm.nih.gov/。拟南芥的基因组数据来源https://ngdc.cncb.ac.cn/gwh/Assembly/21820/show。水稻基因组(MH63RS3)数据来源http://rice.hzau.edu.cn/cgi-bin/rice_rs3/download_ext。月季转录组数据来源于https://www.ncbi.nlm.nih.gov/bioproject/PRJNA587482,https://www.ncbi.nlm.nih.gov/bioproject/722055。
1.2 月季TIFY家族成员全基因组鉴定与系统发育树构建
通过http://pfam.xfam.org/网站下载TIFY结构域的HMM文件(PF06 200)。以HMM文件作为种子文件,利用HMMER3.0检索月季蛋白序列库中TIFY家族成员,参数设置E<1e-5。获取的蛋白序列提交https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi检测其结构域并保存结构域完整的序列。共获取13个家族成员,根据拟南芥同源性重新命名,基因名称见表1。蛋白序列提交https://web.expasy.org/compute_pi/进行理化性质预测,提交https://wolfpsort.hgc.jp/进行亚细胞定位预测。
通过MEGA X软件[10],采用NJ法构建月季TIFY家族成员与拟南芥、水稻TIFY家族成员的系统进化树,参数设置Bootstrap=2 000。进化树结果通过Evolview线上程序https://evolgenius.info//evolview-v2/#login进行美化。
1.3 月季TIFY家族成员的染色体定位与进化分析
通过月季基因组FASTA文件与GFF文件获取染色体长度信息,基因起始中止信息。通过MCScanX程序获取TIFY基因在月季与拟南芥、水稻的共线性基因,提取具有共线性关系的月季TIFY家族成员,结果通过TBtools[11]进行可视化。
1.4 月季TIFY家族成员蛋白保守基序、蛋白结构域与基因结构分析
月季、拟南芥和水稻TIFY家族成员蛋白序列提交https://meme-suite.org/meme/进行motif预测,参数设置motif=10。蛋白序列提交https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi获取结构域信息,参数默认。月季基因组GFF文件获取月季TIFY家族成员UTR区域、内含子和外显子位置的基因结构信息。TBtools进行数据可视化。
1.5 基于RNA-seq数据构建月季TIFY家族成员组织表达热图
月季转录组原始数据通过trim_galore进行质控,参数默认。质控后clean reads通过kallisto程序[12]进行定量,参数设置b=100。定量获取的TPM表达值通过R包sleuth[13]进行结果校正,校正后的结果通过TBtools进行热图可视化。
1.6 月季TIFY家族成员上游顺式元件预测
截取月季TIFY家族成员上游2 000 bp序列提交顺式元件线上预测平台http://bioinformatics.psb.ugent.be/webtools/plantcare/html/,结果通过VB脚本进行统计,TBtools进行数据可视化。
2 结果与分析
2.1 月季TIFY家族成员的鉴定与系统发育分析
为了获取月季基因组中的TIFY基因家族成员,采取HMM和BLASTP 2种方法进行鉴定,初步鉴定结果通过CDD网站进行手动筛选包含完整TIFY结构域的蛋白序列,共鉴定13个月季TIFY家族成员。为了研究TIFY家族进化关系及对家族成员进行进一步细分,采用N-J法将月季、拟南芥和水稻的TIFY家族成员共同构建系统发育进化树,并根据基因同源性对月季TIFY家族成员进行命名,结果如图1。TIFY基因根据特定结构域分为TIFY、JAZ、ZIM和PPD 4个亚家族,13个月季TIFY家族成员中,TIFY亚家族包含1个成员,JAZ亚家族包含8个成员,ZIM亚家族包含3个成员,PPD亚家族包含1个成员。
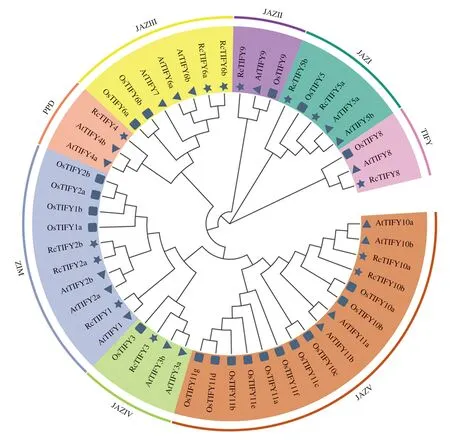
星形:月季;三角形:拟南芥;方形:水稻;RcTIFY:月季TIFY基因家族;AtTIFY:拟南芥TIFY基因家族;OsTIFY:水稻TIFY基因家族。
月季TIFY家族成员均能找到拟南芥、水稻的同源基因,但未鉴定到AtTIFY7/OsTIFY7、AtTIFY11/OsTIFY11的同源基因。这一结果可以推测这2个同源基因在月季进化过程中出现了丢失。
蛋白质序列理化性质预测表明(表1),蛋白长度123~438 aa;相对分子质量为13 813.91~46 086.01 D;等电点为4.87~9.93;RcTIFY1为稳定蛋白,其余蛋白均为不稳定蛋白。蛋白序列亚细胞定位预测结果表明,9个成员被定位在细胞核中,3个成员定位在叶绿体,1个成员定位在线粒体。
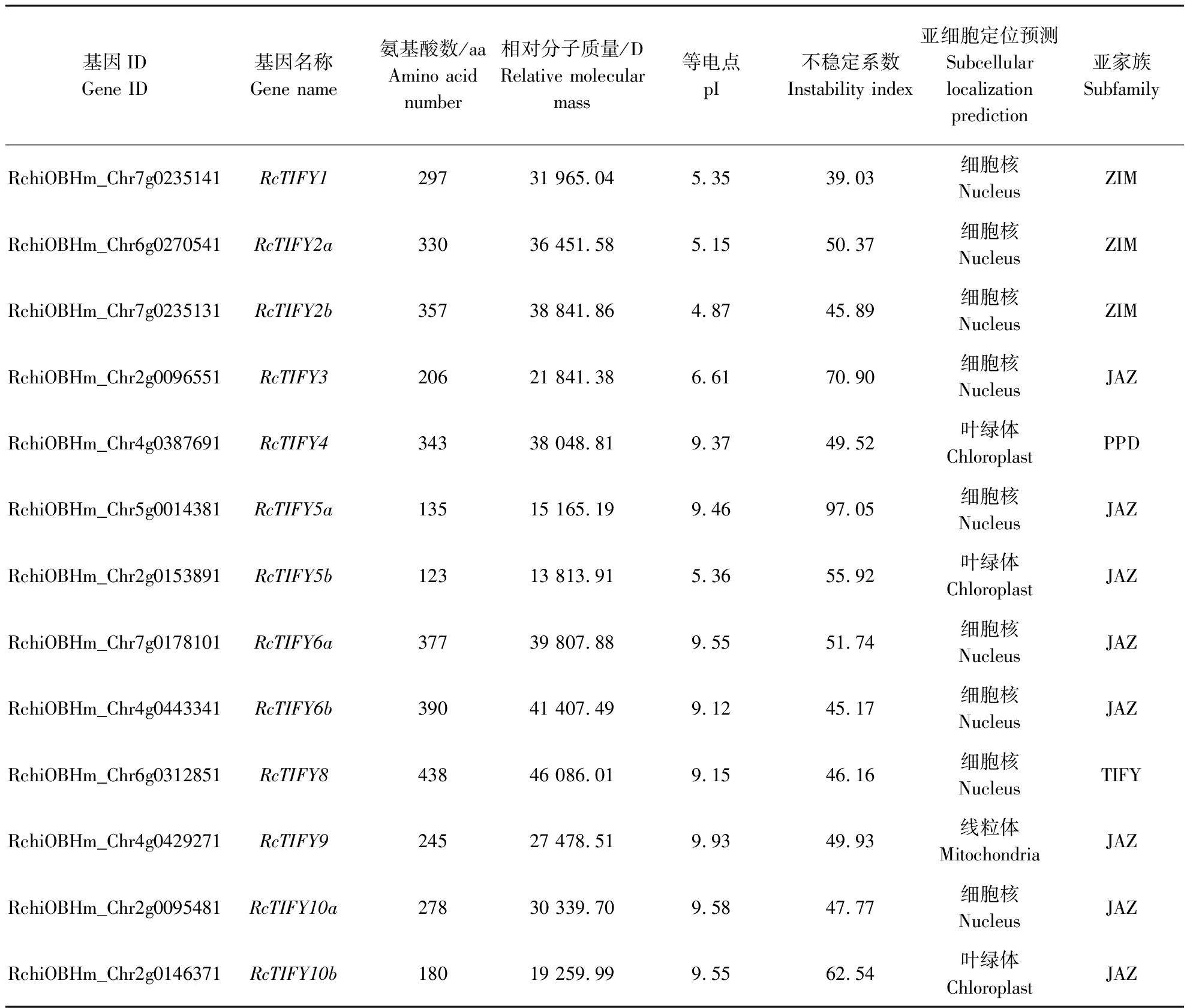
表1 月季TIFY基因家族成员信息Table 1 The information of TIFY gene family members in Rosa chinensis
2.2 月季TIFY家族成员分布与共线性分析
13个月季TIFY家族成员不均匀分布在2、4、5、6、7号染色体中(图2)。其中2号染色体拥有最多的TIFY成员(4个),5号染色体仅包含1个TIFY成员。除此之外,RcTIFY3、RcTIFY5a、RcTIFY6b、RcTIFY6a、RcTIFY8、RcTIFY10a位于相对较高的基因密度区域。
为进一步探究月季TIFY家族成员的进化,基于物种间的共线性分析构建了拟南芥与月季,水稻与月季的基因组共线性图谱。其中月季与拟南芥的TIFY家族成员的同源基因对数为12(图3-A),与水稻的为4(图3-B)。该结果表明月季与拟南芥具有更多的同源基因,也表明月季与拟南芥的分歧发生在二者的共同双子叶植物祖先与稻的单子叶植物祖先的分歧之后。

红色:基因高密度区域;蓝色:基因密度低密度区域。 Red: High gene density region; Blue: Low gene density region.图2 月季TIFY基因家族成员染色体定位Fig.2 Chromosomal localization of TIFY gene family members in Rosa chinensis

A:月季与拟南芥共线性分析,RcChr:月季染色体,AtChr:拟南芥染色体;B:月季与水稻共线性分析,RcChr:月季染色体,OsChr:水稻染色体。
2.3 月季TIFY家族成员motif序列、结构域以及基因结构分析
通过Motif序列,结构域分布以及内含子外显子的位置,可以获取月季TIFY家族成员的结构特征,结果见图4。TIFY家族的4个亚家族具有不同的结构特征,Motif1被鉴定为TIFY结构域并分布在所有基因中。Motif2为Jas结构域(CCT2结构域),该基序仅存在于JAZ和PPD亚家族成员中。Motif2/5为CCT结构域,Motif3为ZnF_GATA结构域,这2种结构域为ZIM亚家族特有。Motif7为PPD结构域,这种结构域为PDD亚家族特有。月季TIFY家族成员均包含完整的UTR区域,同一分支的成员具有相似的外显子/内含子结构。

A:RcTIFY蛋白序列保守基序,底部的刻度单位为aa,表示蛋白长度;B:RcTIFY蛋白序列结构域,底部的刻度单位为aa,表示蛋白长度;C:RcTIFY的基因结构,底部的刻度单位为bp,表示基因长度。
2.4 月季TIFY家族成员转录组表达分析
13个月季TIFY家族成员在干旱处理下叶片的表达热图如图5-A所示,RcTIFY3在干旱处理后表达量先上升,随后稳定,复水后仍不下降。RcTIFY10a、RcTIFY10b、RcTIFY9、RcTIFY5a在干旱处理60 d时表达量达到峰值。RcTIFY6a在干旱处理60、90 d时表达量显著高于0、60 d及复水后7 d。RcTIFY5b在胁迫处理后显著高表达,复水后仍不下降。
13个月季TIFY家族成员在盐胁迫处理下根的表达热图如图5-B所示,RcTIFY9在盐胁迫处理2 h表达量下降,随后表达上升。RcTIFY5b、RcTIFY3、RcTIFY10a、RcTIFY10b、RcTIFY6b、RcTIFY5a具有相似的表达模式,在盐处理24、48 h显著上调。
综上,月季TIFY家族6个成员(RcTIFY3、RcTIFY5a、RcTIFY5b、RcTIFY9、RcTIFY10a、RcTIFY10b)在干旱胁迫与盐胁迫的调节中均发挥着积极作用。

A: RcTIFY基因在干旱胁迫下叶中的表达量;LF_DR_0 d: 叶干旱处理0 d; LF_DR_30 d: 叶干旱处理30 d; LF_DR_60 d: 叶干旱处理60 d; LF_DR_90 d: 叶干旱处理90 d, LF_RW_7 d: 叶复水处理7天。B:RcTIFY基因在盐胁迫下根中的表达量;RT_SA_0 h:根盐处理0 h; RT_SA_2 h: 根盐处理2 h; RT_SA_24 h: 根盐处理24 h; RT_SA_48 h: 根盐处理48 h。
2.5 月季TIFY家族成员顺式元件分析
通过PlantCARE网站对月季TIFY家族成员上游2 000 bp序列的启动子分析表明,这些成员上游序列含有丰富的激素响应元件与非生物胁迫顺式元件。结果如图6,激素响应元件以脱落酸和茉莉酸甲酯响应元件为主。其中脱落酸ABRE响应元件是该家族最多的顺式元件,结合2.4结果,具有丰富的ABRE响应元件的RcTIFY5a、RcTIFY5b、RcTIFY6a、RcTIFY6b、RcTIFY10b均在干旱或盐胁迫下发挥着积极作用。这一结果表明,月季TIFY家族部分成员可能主要通过ABA途径影响植物对胁迫的耐受性。非生物胁迫顺式元件有干旱、盐、热激、应激、低温、创伤、缺氧胁迫元件,这一结果表明月季TIFY家族成员可能对多种胁迫均会响应。
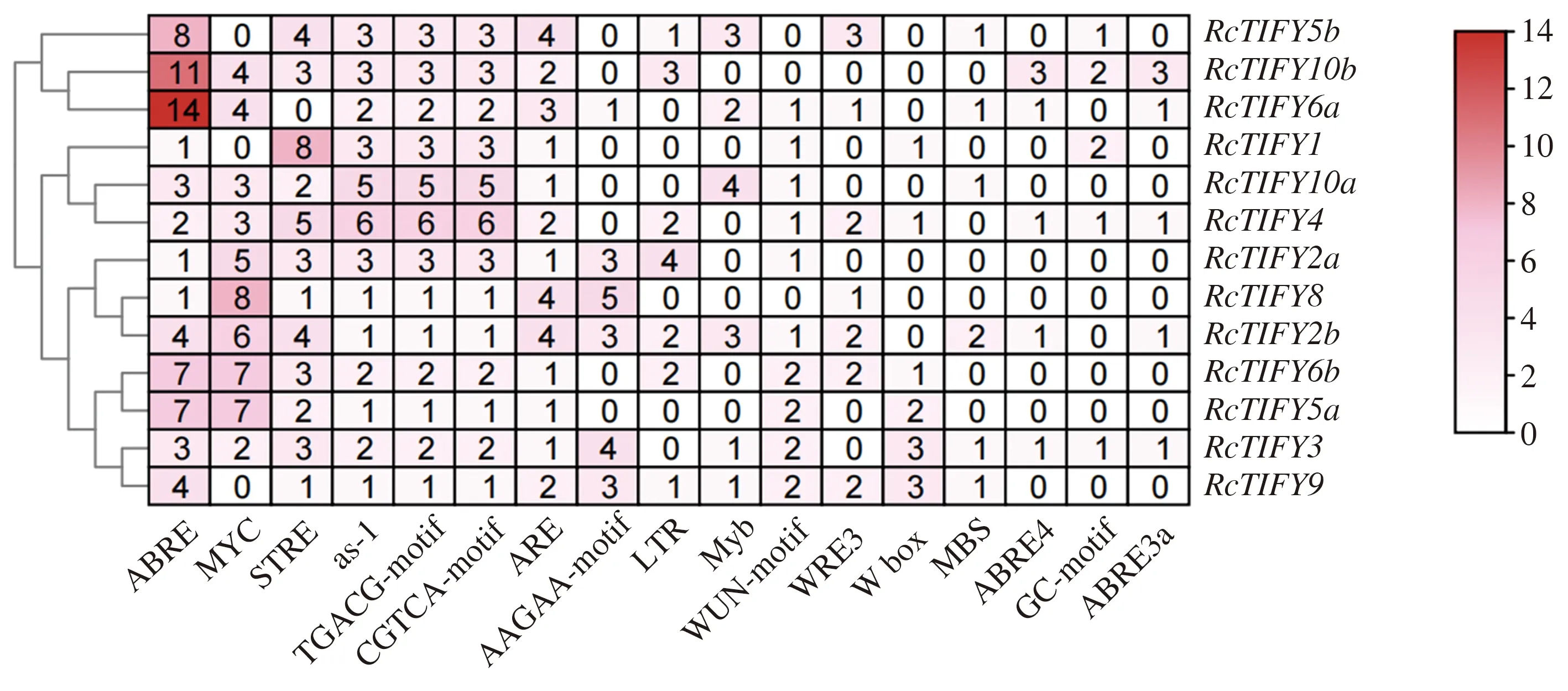
脱落酸顺式元件:ABRE、ABRE4、ABRE3a;茉莉酸甲酯顺式元件:TGACG-motif、CGTCA-motif;干旱胁迫顺式元件:MYC、MBS;盐胁迫顺式元件:Myb;热激胁迫顺式元件:STRE;应激胁迫顺式元件:as-1、ARE、WRE3、W box;低温胁迫顺式元件:LTR;创伤胁迫顺式元件:WUN-motif;缺氧胁迫顺式元件:GC-motif;图例单位为个。
3 讨论与结论
本研究共鉴定13个月季TIFY家族成员,其中TIFY亚家族1个,JAZ亚家族8个,ZIM亚家族3个,PPD亚家族1个。而其他物种TIFY家族成员数在15~54,如西瓜15个[14]、拟南芥18个[15]、水稻20个[7]、玉米30个[16]、棉花54个[17]。月季成员数较少的原因是缺少TIFY7和TIFY11的同源基因,TIFY家族出现的时间早于单子叶和双子叶植物分化的时间[18],水稻和拟南芥中均包含这2种基因,可以推测月季这2种同源基因在进化过程中出现了丢失。本研究中进化树结果显示,PPD亚家族成员仅出现在双子叶植物中。这一结果表明PPD亚家族的出现时间晚于单、双子叶植物分化的时间。本研究结果支持了单子叶植物中不存在PPD基因的假说[2]。亚细胞预测结果表明,月季TIFY家族成员主要位于细胞核,但也有位于叶绿体和线粒体的成员。这一结果与前人研究一致[19]。TIFY家族成员在植物细胞的广泛调控系统中发挥作用,以参与植物生命活动的调节。
TIFY蛋白在调节植物对不利环境条件的反应方面发挥着关键作用,6个月季TIFY家族成员在干旱胁迫后出现了上调表达,7个成员在盐胁迫后出现上调表达,推测这些月季TIFY家族成员在不利的环境条件下同样发挥着关键作用。不同物种的TIFY同源基因具有相似的生物学功能[20]。拟南芥AtTIFY10a和AtTIFY10b的敲除证明了这两个基因在盐碱反应中的积极作用[21],水稻OsTIFY10c基因被证实通过茉莉酸信号通路提高转基因烟草的耐盐性[22],月季中的同源基因RcTIFY10a、RcTIFY10b在盐胁迫24~48 h上调表达,其可能是潜在的增加盐胁迫的抗性的调控因子。拟南芥AtTIFY5b基因介导耐旱性[6],月季中其同源基因RcTIFY5a在干旱胁迫60 d显著上调表达,该基因可能是潜在的抗旱调控因子。
脱落酸(ABA)信号通路在调节各种应激反应中发挥重要作用[23],棉花GbJAZ1、大豆GmTIFY10e和GmTIFY10g通过ABA通路进行盐胁迫调节。外源性茉莉酸或其修饰产物处理可以强烈诱导JAZ亚家族基因的表达,甘蓝中大多数TIFY家族基因在外源茉莉酸甲酯(MeJA)处理后均上调[24]。在月季研究中,这2种激素对月季响应干旱胁迫与盐胁迫具有积极意义[25-27]。本研究中,这2种激素响应元件在月季TIFY家族成员中最丰富,因此推测脱落酸或茉莉酸甲酯可能是有效的提高月季TIFY家族成员积极参与月季的胁迫响应重要因素。
本研究共鉴定13个RcTIFY家族成员,并被分为4个亚家族。其中6个成员(RcTIFY3、RcTIFY5a、RcTIFY5b、RcTIFY9、RcTIFY10a、RcTIFY10b)在干旱胁迫和盐胁迫中均积极响应,转录组结果结合拟南芥与水稻的研究结果,推测RcTIFY10a、RcTIFY10b为潜在的增加盐胁迫的抗性的调控因子;RcTIFY5a是潜在的抗旱调控因子。脱落酸与茉莉酸甲酯是调节该家族成员的潜在关键植物激素。
猜你喜欢
湖北农业科学(2022年11期)2022-07-18
中国特种设备安全(2022年1期)2022-04-26
实用肿瘤学杂志(2020年4期)2020-12-08
中国核电(2017年2期)2017-08-11
上海农业学报(2017年3期)2017-04-10
浙江大学学报(工学版)(2016年2期)2016-06-05
红领巾·探索(2015年9期)2015-09-10
汽车维修与保养(2015年8期)2015-04-17
中国老年学杂志(2015年9期)2015-01-31
植物营养与肥料学报(2014年1期)2014-03-11
