
白云山国家森林公园木本植物空间分布及种间关系
2022-02-07 02:24李子豪孙龙飞付前进苏晓曾曼芸王晶叶永忠袁志良陈云
河南农业大学学报 2022年6期
李子豪, 孙龙飞,付前进, 苏晓, 曾曼芸, 王晶, 叶永忠, 袁志良, 陈云,5
(1.河南农业大学生命科学学院,河南 郑州 450046; 2.河南农业职业学院,河南 郑州 451450;3.洛阳市嵩县五马寺林场, 河南 洛阳 471400; 4.河南林业科学研究院, 河南 郑州 450002;5.河南省伏牛山南北过渡带森林生态系统野外科学观测研究站,河南 洛阳 471400)
森林群落演替是研究森林群落动态变化的最主要途径,也是生态系统演替的一部分。随着演替的进行,森林群落的物种组成成分及数量会发生改变,群落的结构也随之改变[1-4]。一般来说,植物群落的演替总是向着群落结构更加稳定的方向进行的,随着演替的进程,群落的物种丰富度不断增加,群落的整体结构也更加稳定[5]。因此,探究森林群落演替规律有助于深入了解森林群落的动态变化及物种共存机制,为森林生态系统的恢复与保护提供科学依据。
种间关系是指在一定时期内的植物群落不同物种之间的相互关联,是不同种群之间相互影响、相互联系的体现,种间关系决定整个群落的主要特征和功能结构。种间关系主要分为正相关关系,中性关系以及竞争关系[6-7]。探究森林群落中不同物种之间的种间关系有利于解释森林群落的形态、结构、功能。种间关系还可以影响物种的空间分布状况[8]。随着演替的进行,群落的物种组成不断发生变化,种间关系和物种的空间分布格局也随之变化。目前已有将种间联结同演替相联系[7]及将种间联结同群落结构相联系[8]的研究,但鲜有将演替同种间关系及空间分布相结合进行的探讨。
锐齿槲栎(Quercusalienavar.acutiserrata)是壳斗科(Fagaceae)栎属(Quercus)植物,为落叶乔木[9]。针叶树种广泛分布在东北、华北、西北、西南及华南地区的高山地带,亦为森林更新、造林的重要树种。锐齿槲栎(Quercusalienavar.acutiserrata)和针叶树种均为白云山国家森林公园的优势树种。植物群落中的最优势种往往能够影响,甚至是主导整个群落的发展状况,通过对群落中优势种进行分析可以反映出整个群落的结构特点,一个区域的优势种,可被作为研究群落结构特点发展状况的代表物种。通过对比研究优势种的种间关系和空间分布格局的变化,有助于揭示该区域的森林群落整体的形态结构功能,对于正确认识群落的演替和种间关系具有重要意义[10]。
研究种群的空间分布格局是研究种群在空间尺度上的分布状况,是探究群落结构特征及物种共存机制的重要方式,植物物种的空间分布格局一直是生态学研究的热点问题[11-12]。有研究表明,马尾松和杉木作为建群早期先锋种最先入侵林地,在光竞争中占据优势,加剧群落内的光竞争,进而影响群落内的树种空间分布格局[13]。因此,从不同物种的种间关系入手,将物种的种间关系和空间分布格局相互联系,深入探讨不同演替下物种共存的维持机制是具有一定意义的。
白云山国家森林公园位于南暖温带与北亚热带的生态过渡地带,森林覆盖率高,有较大的研究价值。前人已对白云山国家森林公园主要乔木空间分布进行了研究[14],但是还未进行在不同演替下对该区域优势木本植物的空间分布进行研究。为此,根据不同的演替进程,选择人工林、择伐林、皆伐林和老龄林4个1 hm2(100 m×100 m)的永久森林动态监测样地,在前人的基础上深入探究在不同演替下优势木本植物的空间分布格局及种间关系,分析在不同演替下木本植物的空间分布格局的变化及种间关系与空间分布的相互作用,进一步研究整个植物群落结构的变化,从而为该区域生态系统的恢复和保护提供新的角度及参考。
1 研究方法
1.1 研究区域概况
白云山国家森林公园(33°34′~33°38′N,111°48′~111°52′E)位于河南省洛阳市的嵩县南部伏牛山脉,平均海拔1 500 m[15]。降水集中在每年的7—9月份,全年降水量为1 200 mm,年平均气温18 ℃,最高温度不超过26 ℃,相对湿度70%~78%[16]。白云山国家森林公园地处南暖温带与北亚热带过渡区域,植被覆盖率达98.5%[17]。
1.2 样地概况
参照CTFS(Center for Tropical Forest Science)的样地建立标准[18],根据人为干扰程度的不同,设置人工林、择伐林、皆伐林和老龄林4个1 hm2(100 m×100 m)的永久森林动态监测样地[19]。调查统计样地内胸径(diameter at breast height,DBH) ≥1 cm 木本植物,为其编号挂牌,并记录树种名称、胸径、树高及样地内坐标[14,20]。
人工林样地为皆伐后人为飞播落叶松种子并自然恢复50 a的群落。择伐林为先皆伐后自然恢复20 a之后进行择伐,再自然恢复30 a的群落。皆伐林为皆伐后自然恢复50 a的群落。老龄林为自然发展100 a都无人为干扰的群落[21]。
1.3 数据分析
采用重要值分析得出4个样地的优势种为锐齿槲栎[22]。
重要值=(相对多度/% +相对频度/% +相对胸高断面积/%)/3
本研究采用g(r)函数分析锐齿槲栎及松科优势种的空间分布格局,量化物种在各个尺度上的空间分布变化,判断最优势种锐齿槲栎与其他物种的空间关联性。
g(r)函数包括单变量和双变量分析,单变量通常用以对某一物种在某一空间内的分布状况;双变量可以用来分析两个物种之间的种间关系。本研究通过采用蒙特卡罗(Monte-Carlo)随机模拟99次产生置信度为99%的置信区间, 检验物种分布格局及关联性是否显著[23-24]。完全空间随机模型(complete space randomness,CSR)[25]通过偏离置信区间来检验物种空间分布格局呈聚集分布或均匀分布,是匀质泊松过程[26]。异质性泊松(Heterogeneous Poisson Model,HP)[25]是在消除空间及生境的异质性后物种的空间分布格局。对于单变量分析,物种的空间分布形式一般为聚集、均匀和随机3种[27]。g(r)函数值在置信区间以上时,表示该物种为聚集分布;g(r)函数值在置信区间内,表示该物种为随机分布;g(r)函数值在置信区间以下时,表示该物种为均匀分布。对于双变量分析,g(r)函数值在置信区间以上,表示两个物种之间为正相关关系;g(r)函数值在置信区间内,表示两个物种为中性关系;g(r)函数值在置信区间以下时,表示两个物种之间为负相关关系[28-29]。g(r)函数分析均通过R语言Spatstat软件包实现。
2 结果与分析
2.1 不同演替下森林群落物种的径级结构
4个不同演替的森林群落DBH≥1 cm的木本植物共89种,其中人工林41种,植株个体数1 165株,择伐林46种,植株个体数3 605株,皆伐林57种,植株个体数4 302株,老龄林51种,植株个体数2 490株。人工林、皆伐林、老龄林所有木本植物个体(DBH≥1 cm)的径级分布呈“L”型,择伐林的木本植物个体(DBH≥1 cm)的径级分布呈倒“J”型。4个林地中群落更新良好,物种较为丰富,分布紧密。
2.2 不同演替下森林群落物种的空间分布格局
单变量点格局显示(图2),人工林内的物种分布在0~23 m的尺度上为聚集分布,在23~25 m的尺度上呈随机分布。在人工林样地随着尺度不断增加,物种的聚集程度不断减弱,并最终达到随机分布。择伐林与皆伐林内物种呈聚集分布,择伐林物种的聚集程度略有波动;老龄林内的物种在0~14 m的尺度上呈聚集分布,随着尺度的增加,聚集程度不断减弱,在14~25 m尺度上呈现随机分布,略有波动变化。
采用异质性泊松模型排除生境异质性的影响后,人工林、择伐林和皆伐林在全尺度上均为随机分布,随着尺度不断增大,有均匀分布的趋势;老龄林在0~8 m的尺度上呈聚集分布,随机程度随尺度的不断增大而减小,在8~25 m的尺度上呈均匀分布(图3)。

图1 不同演替下森林群落木本植物的径级结构Fig.1 DBH of woody plants in different successional communities

注:实线为实际观测值,虚线为理论值,灰色区间为置信区间。(a)、(b)、(c)、(d)分别表示人工林、择伐林、皆伐林及老龄林样地。下同。
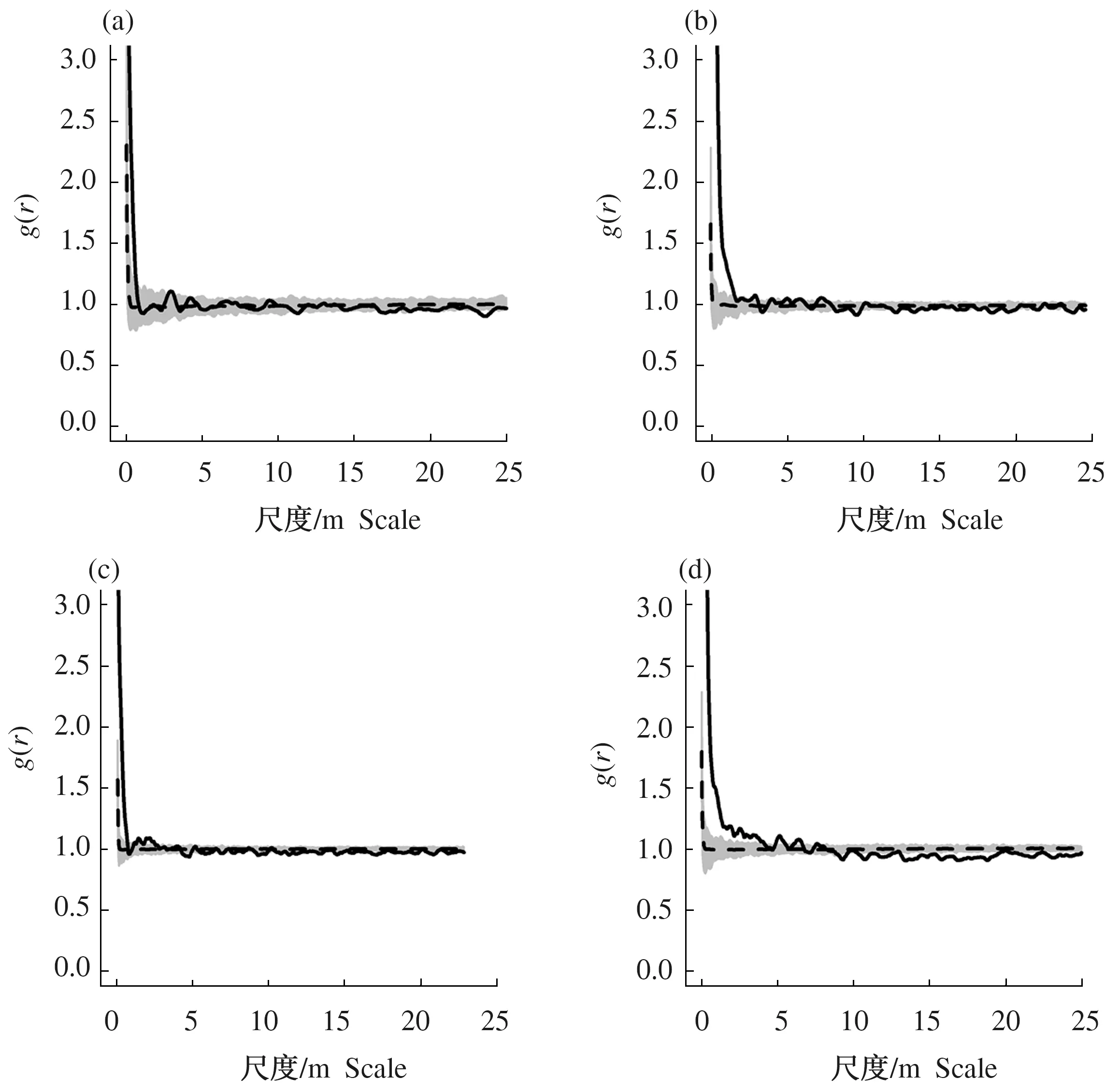
图3 不同演替下森林群落木本植物基于异质性泊松模型的空间分布格局Fig.3 Spatial distribution pattern of woody plants in different successions of forest communities based on HP
2.3 不同演替下锐齿槲栎的空间分布格局
通过完全空间随机模型和点格局分析得出(图4),人工林、择伐林、老龄林中锐齿槲栎在全尺度上呈聚集分布,随着尺度的增大,聚集程度降低;而在皆伐林中,锐齿槲栎的分布在0~10 m尺度下呈聚集分布,但聚集程度较低,在10 m尺度之后呈随机分布。


注:圆圈代表植物,圆圈的大小表示植物的胸径。下同。Note: The circle represent the plants and the size of the circles represents the DBH of the plants. The same as below.图4 不同演替下森林群落锐齿槲栎基于完全随机模型的空间分布格局Fig.4 Spatial distribution pattern of Quercus aliena var. acutiserrata community in different succession based on CSR
在排除生境异质性的影响后,由图5得出,人工林、择伐林及老龄林中的锐齿槲栎在0~5 m的尺度上为聚集分布,5~10 m的尺度上显示为随机分布,10~25 m的尺度上主要为均匀分布;而在皆伐林中,锐齿槲栎在0~4 m上为聚集分布,其余尺度上呈随机分布。
2.4 不同演替下优势针叶树种的空间分布格局
人工林的落叶松在0~1 m的尺度上为随机分布,1~25 m的尺度上呈现明显的聚集分布;择伐林中的华山松在1~2 m的尺度上显示为随机分布,其余尺度上均为随机分布;择伐林和老龄林中的华山松在全尺度上都为显著的聚集分布(图6)。
排除了生境异质性的影响后,由图7可以看出,人工林的落叶松在0~5 m的尺度呈聚集分布,5~15 m的尺度上呈随机分布,15~25 m的尺度上呈均匀分布;择伐林中的华山松在全尺度上均为随机分布;皆伐林在全尺度上呈随机分布,在小尺度范围内有聚集分布,之后有呈均匀分布的趋势;老龄林在0~21 m尺度上呈随机分布,在21~25 m尺度上呈均匀分布。

图5 不同演替下森林群落锐齿槲栎基于异质性泊松模型的空间分布Fig.5 Spatial distribution of Quercus aliena var. acutiserrata community in different succession based on HP

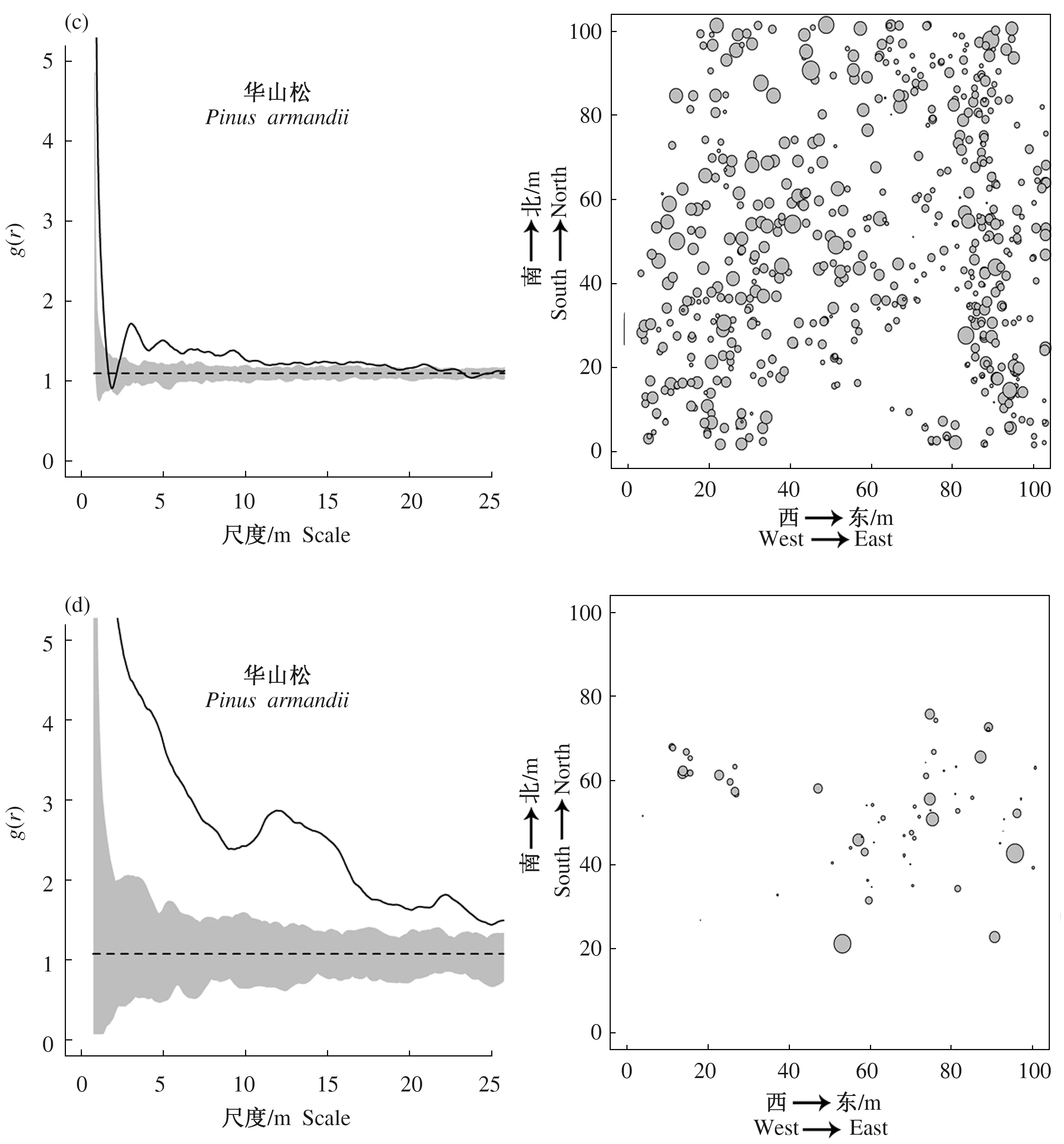
图6 不同演替下优势针叶树种基于完全随机模型的空间分布格局Fig.6 Spatial distribution pattern of dominant conifer in different successions based on CSR
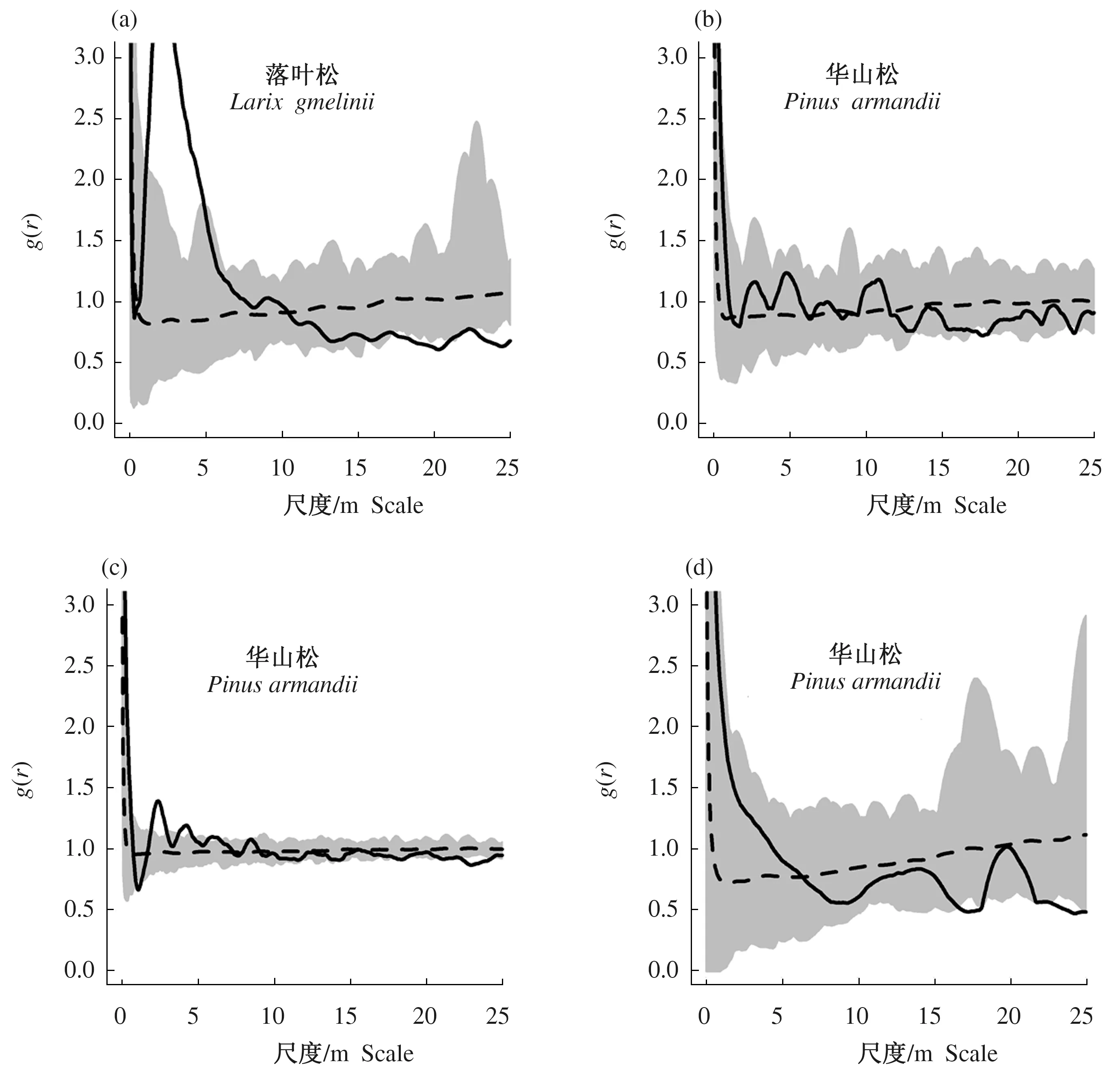
图7 不同演替下优势针叶树种基于异质性泊松模型的空间分布格局Fig.7 Spatial distribution pattern of dominant conifer under different successions based on HP
2.5 不同演替下锐齿槲栎和优势针叶树种的种间关系
种间关系分析表明(图8),人工林锐齿槲栎与落叶松、择伐林中的锐齿槲栎和华山松在全尺度上均呈负相关,其中人工林锐齿槲栎和落叶松随着尺度不断增加竞争关系越来越弱;皆伐林中锐齿槲栎和华山松在0~5 m的尺度上为负相关,5~25 m的尺度上两者无明显相关性;老龄林中锐齿槲栎和华山松在11~16 m的尺度上为正相关,其余尺度上为中性关系。
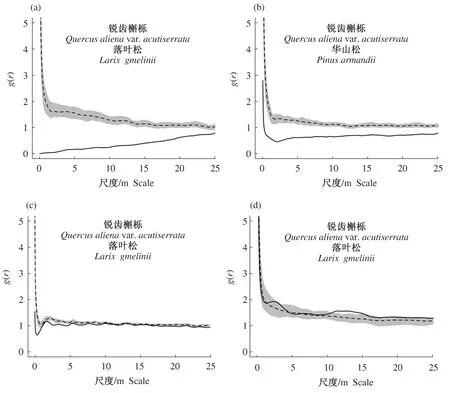
图8 不同演替下锐齿槲栎与优势针叶树种种间关系Fig.8 Interspecific relationships between Quercus aliena var. acutiserrata and dominant conifer in different successions
3 讨论与结论
4个林地木本植物小径级结构个体占比较多,择伐林木本植物个体径级结构呈现倒“J”型,其余林地木本植物个体径级结构呈“L”型,4个林地群落更新状态良好。植物群落的空间分布同尺度是息息相关的,随着空间尺度的不断变化,其空间分布特征也随之变化[12,30]。在不排除环境影响的情况下,在4个样地物种均为聚集分布;使用异质性泊松分析,排除环境的影响后,4个样地在小尺度上表现为聚集分布,但在全尺度服从于随机分布,由此可见生境异质性对于植物物种的空间分布影响显著,生境异质性使植物趋于聚集分布[31]。伴随着演替的进行,植物的空间分布也发生了变化,老龄林在排除生境异质性的干扰后,在大尺度范围内并没有呈现聚集分布,而是呈现均匀分布,原因可能是老龄林长期没有受到干扰,森林群落演替程度最高,整个森林群落发展稳定。
锐齿槲栎在4个不同演替下呈现显著的聚集分布,在排除了生境异质性后,小尺度上依然呈现聚集分布,一般认为小尺度范围内的分布格局物种其自身的生物学特性影响更为显著[12,23]。大尺度上显示为随机或均匀分布,随机或均匀分布的格局可是与自疏及他疏现象有关[12,23,32-33]。优势针叶树种在排除生境异质性后在4个不同干扰林下基本上呈现随机分布或是均匀分布,人工林小尺度范围内呈现聚集分布可能是由于种子扩散限制的原因。
植物的种间关系对植物种群及整个群落的发展有显著的影响。在不同演替阶段锐齿槲栎和优势针叶树种的种间关系有显著区别,人工林与择伐林中两者呈明显的负相关关系,在皆伐林中两者接近中性关系,老龄林中两者全尺度服从于中性关系,并且偶有正相关关系出现。伴随着演替的进行,锐齿槲栎同优势针叶树种的竞争不断减弱,由负相关到中性关系再到正相关,这与李霞等[7]研究结果一致。正相关关系的出现可能是因为经过长期的演替过程后,老龄林作为其中较高级的演替阶段,同时缺乏干扰,内部树种在经过长期演替斗争后,逐渐达到了一种平衡状态,各物种种间关系也更加稳定,生态位重叠度更高,整个森林群落也更为稳定[7,13]。
植物物种的空间分布同时也受到该物种种间关系的制约,种群之间的竞争关系导致物种存在生存压力,强烈的竞争关系会体现在优势种对于弱势种群的压迫而使其产生生存压力,从而影响整个群落的空间分布状况。随着演替的进行,锐齿槲栎和优势针叶树种两者的竞争关系不断减弱,中性关系和正相关关系增强,同时在排除生境异质性干扰后,锐齿槲栎的聚集程度不断降低,取而代之随机分布和均匀分布的程度增加。说明种间关系的变化对于植物物种的空间分布产生了影响,种间关系的缓和使锐齿槲栎分布逐渐向随机分布及均匀分布靠拢。
综上所述,不同演替下森林群落木本植物空间分布格局受到显著的生境异质性影响;随着演替的进行,物种的空间分布逐渐从聚集分布趋于随机均匀分布,物种的竞争关系减弱,种间关系逐渐从负相关关系趋于正相关关系,群落物种组成趋于稳定。通过对不同演替下木本植物空间分布及其种间关系变化的研究,揭示了不同演替下物种的空间分布与种间关系的联系与相互作用,为该区域生态系统的恢复和植物多样性保护提供新的角度及参考。
猜你喜欢
老龄科学研究(2022年9期)2022-10-12
当代水产(2022年8期)2022-09-20
河南师范大学学报(自然科学版)(2022年5期)2022-08-08
昆明医科大学学报(2022年2期)2022-03-29
食品安全导刊(2021年20期)2021-08-30
烟台果树(2021年2期)2021-07-21
烟台果树(2021年2期)2021-07-21
河南科学(2020年3期)2020-06-02
蔬菜(2018年5期)2018-05-17
体育科研(2016年2期)2016-02-28
