
谷子不同轮作模式对土壤理化性质及细菌群落的影响
2022-02-06 02:07李小霞李万星曹晋军靳鲲鹏韩文清胡丹珠黄学芳刘永忠
江苏农业学报 2022年6期
李 丹, 李小霞, 李万星, 曹晋军, 靳鲲鹏, 韩文清, 胡丹珠, 刘 鑫, 田 岗, 黄学芳, 刘永忠
(1.山西农业大学谷子研究所,山西长治046000;2.山西农业大学山西有机旱作农业研究院,山西太原030000)
连作是指同一种植物或近缘植物在同一块地上连续种植两茬或以上的现象。连作可导致土壤中的某种养分过度消耗或产生自毒作用使得作物生长发育迟缓,病虫害发生概率增加,农产品产量降低。谷子是不宜重茬的作物之一,近年来谷子种植面积不断缩减,但是由于其丰富的营养价值和保健作用,较高的经济效益,农户种植经济性较高,导致连作现象频发[1]。
已有研究结果表明,避免连作障碍的最佳防治途径是与不同作物轮作[2]。轮作可以合理利用土壤养分,调节土壤酸碱度,改善土壤结构,减轻病虫害发病率。孙倩等[3]发现,轮作能够改善土壤酶活性,轮作模式下谷茬土壤中的过氧化氢酶、脲酶、碱性磷酸酶活性提高,土壤细菌群落丰富度指数降低。蔡晨等[4]研究发现,与水稻单作相比,稻虾长期轮作可使0~20.0 cm和20.1~40.0 cm土层容重降低,非毛管孔隙度、毛管孔隙度、总空隙度提高,同时增强土壤缓冲能力,提高耕层土壤pH。熊湖等[5]研究结果表明,连作可以改变马铃薯植株形态特征(株高、茎粗、总叶面积降低,根长增加),通过轮作使土壤中速效养分转化速率加快,可以缓解连作胁迫。
本研究拟以特色杂粮谷子为研究对象,探讨不同作物与谷子轮作后对土壤的影响,包括物理结构、土壤养分、细菌多样性,筛选适用于晋东南武乡地区的最佳谷子轮作模式,以期对谷子特色产业的可持续、绿色发展提供指导。
1 材料与方法
1.1 试验地概况及试验设计
大田试验于2017-2020年在山西省长治市武乡县上司镇铺上村(E112°93′,N36°76′)进行。2018年至2020年谷子生育期内平均降水量为379.7 mm,为典型的雨养农业区。土壤为典型的红壤土,前茬为谷子,基础肥力为:有机质含量10.24 g/kg,全氮含量0.089%,速效磷含量29.14 mg/kg,速效钾含量246.35 mg/kg,碱解氮含量46.96 mg/kg,pH值为8.28。试验设置3次重复,随机区组排列,小区面积7.7 m×7.0 m,有机肥为羊粪,施入量为60 000 kg/hm2,不同小区管理方式一致。各处理设置见表1。

表1 试验设置
供试谷子为长生13,玉米为晋单73,大豆为长豆35号,高粱为晋杂22号,花生为长治县裕盛丰农业专业合作社提供的黑美人,马铃薯为晋薯16号,甘薯为山西省长治市长子县当地特有品种,玉米(糯)为晋糯10号。
1.2 土壤样品采集
用于测定土壤性质及土壤团聚体的土壤样品于谷子成熟期采集:清除地表杂物后用环刀分别采集0~10.0 cm、10.1~20.0 cm的土壤装入铝盒内带回实验室并置于室内风干,运输过程中尽量避免碰撞挤压。用于测定土壤肥力的根际土壤样品采集:于收获期采集每种作物的根际土壤,随机选取5株作物,将作物连根拔起,抖落外侧大型土块,去除石子、根系等杂物,充分混匀后过筛,随后带回实验室自然风干,备用。用于测定细菌多样性的根际土壤样品采集:于灌浆期分别采集不同种植模式的根际土壤,方法同上。每个处理样品分成2份,一份放入冻存管中,液氮保存寄送至上海派森诺生物科技有限公司用于土壤真菌多样性的检测,一份放入-80 ℃备用。
1.3 测定指标与方法
1.3.1 土壤容重及土壤水稳性团聚体测定 于谷子收获期采用环刀法在田间测定土壤容重。
土壤水稳性团聚体采用湿筛法[6-8]测定:将处理好的土壤置于团聚体分析仪套筛上部筛网上,套筛孔径依次为2.00 mm、1.00 mm、0.50 mm和0.25 mm,桶中加入蒸馏水使水没过土样,浸泡10 min后开始振荡(振荡10 min,上下移动30次,振幅3.00 cm)。振荡结束后将套筛取出,将每个筛子上面的土壤冲洗到铝盒内,静置15 min后用塑料滴管去除上清液,烘箱烘干,称量,得到粒径<0.250 mm、0.251~0.500 mm、0.501~1.000 mm、1.001~2.000 mm、>2.000 mm的水稳性团聚体质量。
1.3.2 土壤养分测定 土壤养分、酶活性的测定采用常规测定方法[9-11]。
1.3.3 土壤细菌多样性测定 根据DNeasy PowerSoil Kit(QIAGEN)试剂盒说明书对土壤总DNA进行提取,利用NanoDrop2000对提取的DNA质量浓度和纯度进行检测;对细菌16S区进行扩增。对构建好的文库进行检验、测序。对测序结果进行操作单位聚类与分析、Alpha多样性分析、Beta多样性分析及不同分类水平的群落结构分析。
1.4 数据计算与统计分析
采用Excel2000软件进行数据整理与分析,用IBM SPSS20.0软件进行单因素方差分析和主成分分析。
计算各粒级团聚体含量、各粒级团聚体比例、平均质量直径(MWD),公式[12-14]如下:
各粒级团聚体含量=各级团聚体质量/土样样品干质量×100%
(1)
各粒级团聚体比例=某级团聚体质量/团聚体总质量×100%
(2)
(3)
式中,MWD为平均质量直径;wi为第i级团聚体的质量占土壤样品干质量的质量百分数;Ri为第i级团聚体平均直径。本研究中各级团聚体平均直径分别为2.00 mm、1.50 mm、0.75 mm、0.38 mm、0.25 mm。
2 结果与分析
2.1 谷子不同轮作模式对土壤容重和水稳性团聚体的影响
2.1.1 土壤容重 图1显示,不同处理下土壤容重由大到小为:SSSi、SZGm>SZZm>SZSt、SZLb>SZSb>SZAh,SSSi处理下土壤容重为1.25 g/cm3,其他6个处理的土壤平均容重为1.19 g/cm3,较SSSi处理下的土壤容重降低了4.8%。其中SZAh、SZSb、SZZm、SZSt、SZLb处理下土壤容重较SSSi处理下的土壤容重降低了2.4%~8.8%,SZAh处理下土壤容重最低。

SSSi、SZGm、SZZm、SZSt、SZLb、SZSb、SZAh见表1。不同小写字母表示不同处理间差异显著(P<0.05)。图1 不同处理下的土壤容重Fig.1 Soil bulk density under different treatments
2.1.2 土壤水稳性团聚体的粒径分布 图2显示,0~10.0 cm和10.1~20.0 cm土层中各粒级团聚体占比变化趋势一致:占比随团聚体粒径减小而降低。在0~10.0 cm土层,各处理下粒径>2.000 mm土壤水稳性团聚体占比为42.98%~53.26%,在SZSb处理下的占比最低,在SZSt处理下的占比最高;粒径为1.001~2.000 mm的土壤水稳性团聚体占比26.72%~32.72%,在SZSt处理下的占比最低,在SZSb处理下的占比最高;粒径为0.501~1.000 mm的土壤水稳性团聚体占比9.90%~16.09%,在SZAh处理下的占比最低,在SZSb处理下的占比最高;粒径为0.251~0.500 mm的土壤水稳性团聚体占比6.16%~7.86%,在SSSi处理下的占比最低,在SZZm处理下的占比最高;粒径为0.100~0.250 mm的土壤水稳性团聚体占比0.22%~4.67%,在SSSi处理下的占比最低,在SZZm处理下的占比最高。在10.1~20.0 cm土层,各处理中粒径>2.000 mm的土壤水稳性团聚体占比为41.78%~52.55%,在SZSb处理下的占比最低,在SZLb处理下的占比最高;SZSb处理中粒径为1.001~2.000 mm、0.501~1.000 mm、0.251~0.500 mm的土壤水稳性团聚体占比均最高,分别为32.94%、15.17%、8.13%;粒径为1.001~2.000 mm的土壤水稳性团聚体在SZSt处理中占比最低,为26.61%;粒径为0.501~1.000 mm的土壤水稳性团聚体在SZLb处理中占比最低,为11.45%;粒径为0.251~0.500 mm、0.100~0.250 mm的土壤水稳性团聚体在SSSi处理中占比最低,为6.70%、0.17%;粒径为0.100~0.250 mm的土壤水稳性团聚体在SZGm处理中占比最高,为5.51%。

SSSi、SZGm、SZZm、SZSt、SZLb、SZSb、SZAh见表1。图2 不同处理土壤团聚体粒级分布Fig.2 Particle size distribution of soil aggregates under different treatments
2.1.3 土壤水稳性团聚体组成及稳定性 可用平均质量直径(MWD)反映土壤水稳性团聚体稳定性,MWD越大,代表土壤团聚体的团聚度越高,该土壤越稳定[15-16]。表2显示,同一处理10.1~20.0 cm土层的MWD均大于0~10.0 cm土层。0~10.0 cm土层中,MWD由大到小为:SZGm>SZZm>SSSi、SZSb>SZAh>SZLb、SZSt;10.1~20.0 cm土层中,MWD由大到小为:SZGm>SZZm>SZSb>SSSi>SZAh、SZSt>SZLb。在0~10.0 cm、10.1~20.0 cm分层土壤中,SZGm处理下的MWD值最大,且与其他处理差异显著,说明SZGm处理下土壤稳定性最优,SZSt、SZLb处理下土壤稳定性差,猜测块茎、块根类作物可降低土壤团聚体稳定性。
2.2 谷子不同轮作模式对土壤基础肥力的影响
2.2.1 土壤养分 表3显示,不同处理下土壤全氮含量、碱解氮含量、速效磷含量、速效钾含量、有机质含量、pH与初始土样相比均发生改变。土壤全氮、有机质含量均提高,增幅达30.3%~59.6%、96.4%~139.0%,SSSi处理下全氮、有机质含量最高,SZGm处理下全氮含量最低,SZAh、SZLb处理下有机质含量最低;不同处理下土壤均为碱性状态,SSSi、SZZm处理下土壤pH与初始土壤相比呈降低趋势;SZGm、SZLb、SZZm、SSSi处理下碱解氮含量升高,SZAh、SZSt处理下降低,SZZm处理下碱解氮含量最高,SZAh处理下碱解氮含量最低;SZGm、SSSi、SZSb、SZAh处理下速效磷含量升高,增幅为0.9%~56.5%,SZLb、SZSt、SZZm处理下速效磷含量降低,降幅为13.9%~26.2%;SSSi处理下的速效钾含量变化不明显,其他处理下的速效钾含量均明显降低,降幅为16.9%~26.8%,SZLb处理下的速效钾含量最低。

表2 不同处理土壤水稳性团聚体组成特征

表3 不同处理土壤养分含量
2.2.2 土壤酶活性 不同种植模式会影响土壤酶活性。过氧化氢酶可催化土壤中的过氧化氢分解为水和氧气,是一种与土壤呼吸作用相关的酶;同时可以分解土壤中的有害自由基,与自净能力有关。表4显示,不同处理下过氧化氢酶活性从大到小为:SZZm>SZGm>SZAh>SZSb>SSSi>SZLb>SZSt,说明SZZm处理下土壤呼吸强度高,自净能力强,SZSt处理下最差。碱性磷酸酶在磷转化方面发挥着重要作用,可以加速有机磷的脱磷速度,该酶主要由土壤细菌分泌,可将土壤中的有机磷水解为可供植物利用的无机磷[17],不同处理下碱性磷酸酶活性由大到小为:SZSb>SZGm>SZLb、SZZm>SZSt>SSSi、SZAh。
脲酶可以分解土壤中的含氮化合物,是速效氮的主要来源,脲酶活性是评价土壤酶活性和肥力的重要指标,不同处理下土壤中脲酶活性大小顺序依次为:SZZm>SSSi>SZAh>SZLb>SZGm>SZSb>SZSt,SZZm处理下脲酶活性最高,比脲酶活性最低的SZSt处理高31.0%,其他5个处理间差异不显著。
纤维素酶可由土壤真菌、细菌和放线菌分泌产生,能够降解秸秆中的纤维素,将养分释放到土壤中[18]。不同处理下土壤中纤维素酶活性由大到小为:SZSt>SZLb>SZAh>SSSi>SZZm>SZGm>SZSb,SZSt处理下纤维素酶活性最大,比酶活性最低的SZSb处理高47.8%。
转化酶又称蔗糖酶,能够促进蔗糖生成葡萄糖和果糖,可以促进土壤熟化,增加土壤中的速效养分[19]。不同处理下土壤中蔗糖酶活性由大到小为:SZLb>SZZm、SZAh>SZSb>SSSi>SZSt、SZGm,SZLb处理下蔗糖酶活性最高,比活性最低的SZGm、SZSt处理高35.3%。
蛋白酶可将土壤中蛋白质和含氮化合物分解为植物可以直接吸收利用的氨基酸或活性氮[20]。不同处理下土壤中蛋白酶活性由大到小为:SZLb>SZSt>SZZm>SSSi>SZGm>SZSb>SZAh,SZLb处理下蛋白酶活性最高,且与其他处理差异显著,比酶活性最低的SZAh处理高174.2%。

表4 不同处理土壤酶活性
2.3 谷子不同轮作模式对细菌群落结构的影响
2.3.1 土壤细菌组成 图3显示,不同处理的土壤具有相似的细菌组成,但是比例上存在一定差异。其中相对丰度大于1.00%的有6个门类,分别是Proteobacteria、Actinobacteria、Chloroflexi、Acidobacteria、Bacteroidetes和Gemmatimonadetes,6个门的丰度占总丰度的94.50%~96.70%。其中,Proteobacteria、Actinobacteria为优势菌门,在各处理中占比达64.20%~74.10%。与SSSi处理相比,SZLb、SZSt处理中Proteobacteria的相对丰度加,分别增加了2.30%、1.60%,其他处理中Proteobacteria的相对丰度降低,降低了3.60%~17.20%。与SSSi处理相比,SZSt、SZSb、SZAh、SZGm处理中Actinobacteria的相对丰度增加了3.98%~30.70%,SZLb、SZZm处理中Actinobacteria的相对丰度降低了7.50%、6.30%。另外,Nitrospirae、Verrucomicrobia、Patescibacteria、Rokubacteria是另外4个主要的门(至少在一个处理中的相对丰度大于0.50%)。
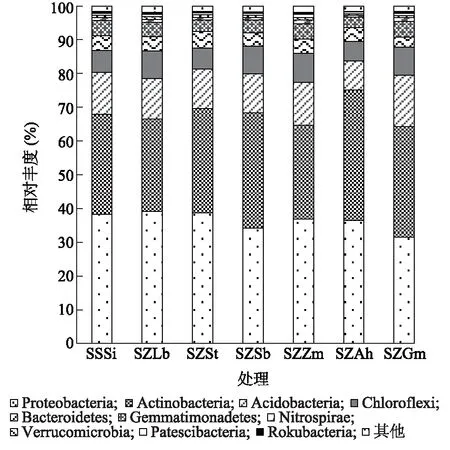
SSSi、SZGm、SZZm、SZSt、SZLb、SZSb、SZAh见表1。图3 不同处理土壤细菌门水平的相对丰度Fig.3 Relative abundance of soil bacteria at phyla level in different treatments
不同处理土壤细菌属水平群落结构分析结果(图4、图5)表明,7种轮作模式聚类图没有明显分支,说明土壤微生物结构相似,但是比例上存在差异。相对丰度大于1.00%或至少在1个处理中的相对丰度大于1.00%的优势属有27个,7个处理共有的优势属有9种,分别为Subgroup_6占比4.5%~8.1%,BIrii41占比约1.9%~3.9%,KD4-96占比1.8%~3.4%,Sphingomonas占比1.8%~3.4%,MND1占比1.3%~3.6%,Blastococcus占比1.5%~2.4%,RB41占比1.0%~2.2%,Nocardioides占比1.0%~2.1%,Haliangium占比1.0%~1.3%。TRA3-20、IMCC26256为SZGm处理特有的属,Acidibacter、CCD24为SZZm处理特有的属,Micromonospora为SZAh处理特有的属,剩余13属为2个或多个处理的共有属。聚类分析结果(图5)表明,SZLb、SZSb处理与SSSi处理距离最远,说明这2个处理对于细菌群落的影响大于其他处理,SZAh处理与SSSi处理距离最近,说明这2个处理的细菌组成最相似。

SSSi、SZGm、SZZm、SZSt、SZLb、SZSb、SZAh见表1。图4 不同处理土壤细菌属水平相对丰度的比较(前20)Fig.4 Comparison of relative abundance of soil bacteria at genus level in different treatments (top 20)

SSSi、SZGm、SZZm、SZSt、SZLb、SZSb、SZAh见表1。图5 不同轮作模式聚类分析Fig.5 Hierarchical clustering analysis of different crop rotation patterns
2.3.2 土壤细菌多样性 各处理的Chao1指数(丰富度)间没有显著差异,与SSSi处理相比,其他6个处理的Chao1指数均有所提高;与SSSi处理相比,其他6个处理的香浓指数均降低;各处理的辛普森指数间没有显著差异(表5)。综合3个指数可推断,与连作相比,轮作会改变土壤微生物群落结构:物种种类(丰富度)增多,而物种多样性降低,更利于无优势种群的生存。
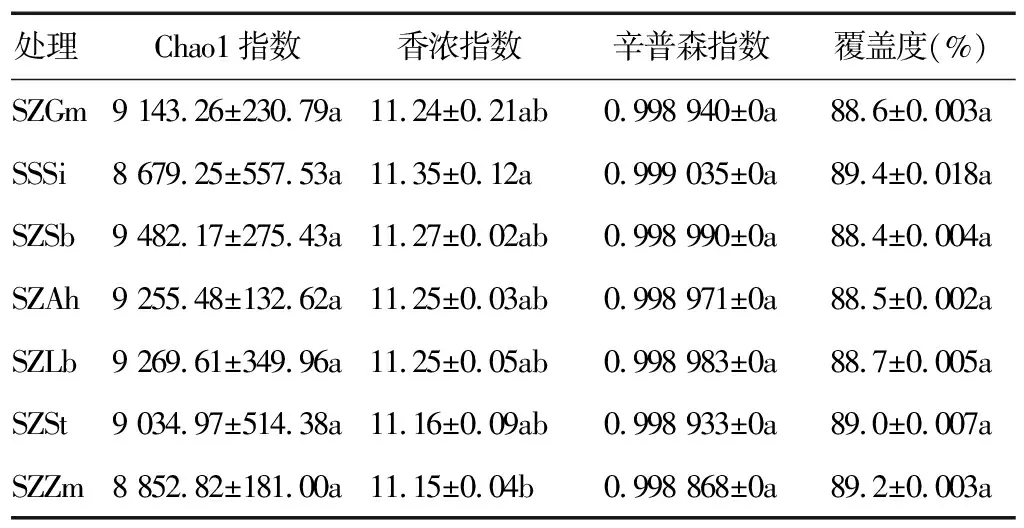
表5 不同处理土壤细菌多样性指数
2.4 土壤养分与细菌优势属的相关性分析
表6显示,蛋白酶活性是影响土壤细菌群落最主要的因子。过氧化氢酶活性与Devosia呈显著负相关(P<0.05);碱性磷酸酶活性与Gitt-GS-136呈显著正相关(P<0.05);纤维素酶活性与Altererythrobacter呈显著正相关(P<0.05);转化酶活性与Haliangium呈显著正相关(P<0.05);蛋白酶活性与Blastococcus呈极显著负相关(P<0.01),与Lysobacter呈极显著正相关(P<0.01);碱解氮含量与MND1、Nitrospira、TRA3-20、CCD24呈显著正相关(P<0.05),与Actinoplanes呈显著负相关(P<0.05);速效磷、速效钾含量与Sphingomonas呈显著正相关(P<0.05)。
2.5 谷子不同轮作模式对土壤物理、肥力指标影响的综合分析
2.5.1 土壤相关指标选取与主成分分析结果 选取具有代表性的土壤相关因子,包括:物理结构(土壤容重、10.1~20.0 cm的MWD)、土壤养分(全氮、碱解氮、速效磷、速效钾、有机质)含量及土壤酶(过氧化氢酶、碱性磷酸酶、脲酶、纤维素酶、转化酶、蛋白酶)活性,共13个指标作为主成分分析的评价指标。为使选取因子在数量级和量纲上达到一致,需对原始数据进行标准化处理,然后对土壤物理、养分指标进行相关性分析。
谷子土壤相关指标主成分分析结果(表7)表明,从13个因子中提取出5个主成分且5个主成分累积贡献率高达98.076%,即5个主成分可包含原有信息的98.076%,符合分析要求,所以这5个主成分可以代表谷子土壤相关指标进行分析。

表6 土壤肥力指标与优势属之间的相关性分析
2.5.2 不同处理谷子土壤物理、肥力综合排名 综合分析土壤理化性质等相关指标,得分最高的为SZZm处理(表8),其次为SSSi处理,得分最低的为SZSt处理。认为谷子-玉米-玉米(糯)-谷子轮作模式可以使土壤环境保持最优。
3 讨论
土壤肥力对农作物生长发育至关重要[21-26],合理轮作可以提高土壤养分含量,改善土壤结构[271]。本研究结果表明,合理的轮作可降低土壤容重,增强土壤团聚体稳定性(块根、块茎类作物除外);谷子连作降低了土壤pH,使土壤趋于酸化,这与牛倩云等[28]、徐宁等[29]的研究结果一致;谷子连作模式下,土壤中速效磷、速效钾、有机质含量均居于首位,袁洪超等[30]在当归连作研究中得到类似结论。这可能与不同种植模式作物对养分的需求不同有关。
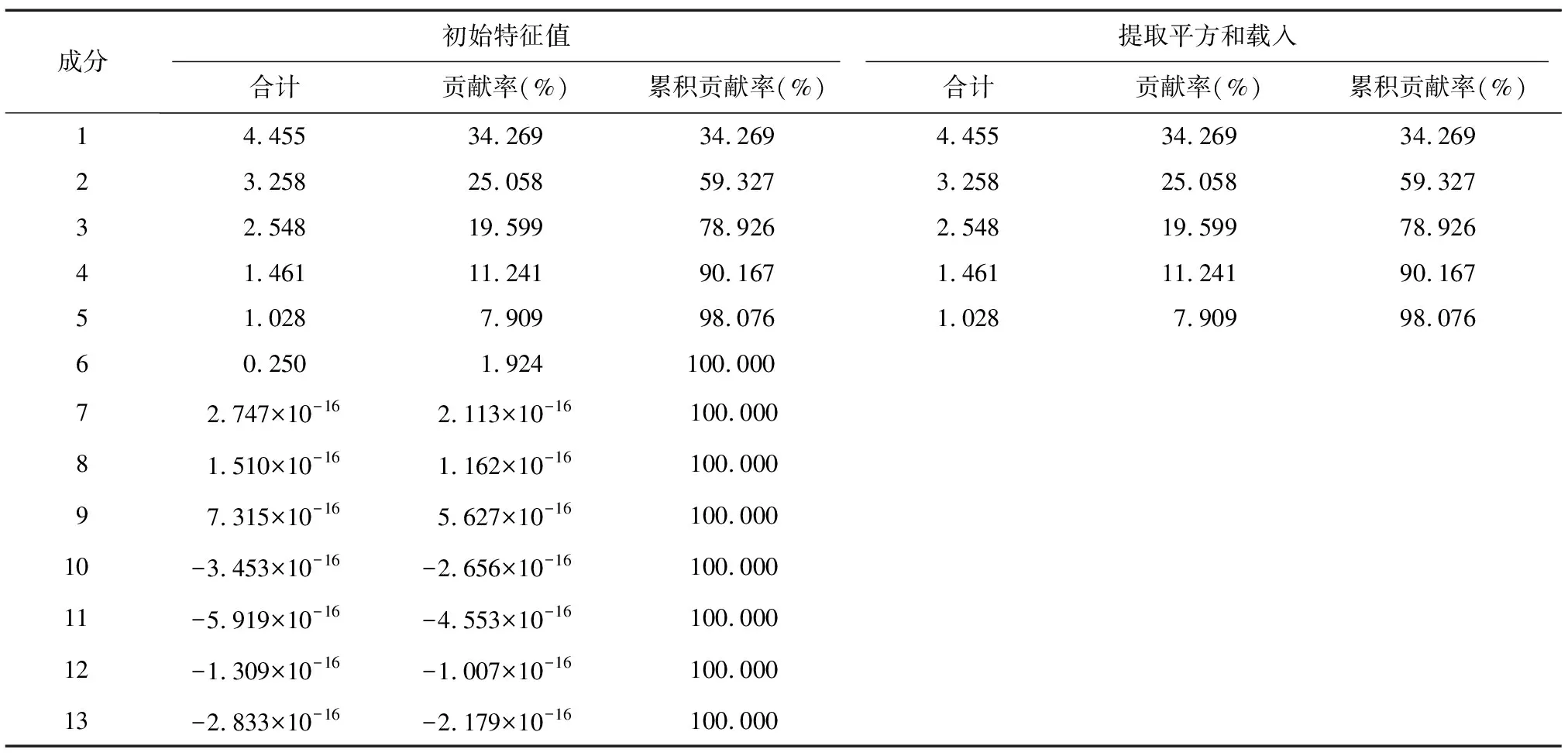
表7 主成分特征值与贡献率

表8 不同轮作模式因子得分与综合排名
土壤酶是作物根系和微生物产生的一类蛋白质代谢产物,参与土壤的各种生化反应。有研究认为,合理轮作有利于提高土壤酶活性[31-32]。本研究中,马铃薯轮作模式的过氧化氢酶、脲酶活性最低;谷子连作模式、花生轮作模式碱性磷酸酶活性最低;高粱轮作模式纤维素酶活性最低。总体来讲,合理的轮作有利于提高土壤酶活性,不合理的轮作会降低土壤酶活性。本试验中7种轮作模式的土壤酶活性没有明显规律,可能与试验地常年施用有机肥以及试验周期较短(4年)、连作障碍不明显有关。
谷子连作模式的土壤细菌丰富度指数最低,但是其多样性指数最高,推测与土壤酸化有关,该条件下更利于优势菌的生存。聚类分析结果表明,不同轮作模式细菌群落结构组成基本相同,但在比例上有所差异,推测土壤细菌对于环境变化较不敏感,前人在番茄上得到类似结论[33]。
4 结论
为了筛选出适宜晋东南地区谷子最佳的轮作作物,使土壤保持健康生态环境,本研究通过主成分分析法可知,不同作物与谷子轮作的综合得分顺序为SZZm>SSSi>SZSb>SZGm>SZLb>SZAh>SZSt。其中,轮作玉米(糯)得分最高,其次依次为谷子连作、轮作高粱、轮作大豆,且四者得分均为正值,说明轮作玉米(糯)、高粱、大豆以及连作谷子可使谷子土壤保持较健康的水平。轮作马铃薯、花生、甘薯三者得分为负值,说明这3种轮作方式降低了土壤质量。由此可知,不同轮作模式会影响土壤的质量,且长时间连作会产生连作障碍,玉米(糯)是较为适宜晋东南武乡地区避免谷子连作障碍的理想作物。
猜你喜欢
灌溉排水学报(2022年9期)2022-09-28
舰船科学技术(2022年10期)2022-06-17
全球定位系统(2022年1期)2022-04-14
农业工程学报(2022年1期)2022-03-25
今日农业(2021年11期)2021-11-27
装备制造技术(2020年3期)2020-12-25
全球定位系统(2019年4期)2019-08-30
今日农业(2019年12期)2019-08-15
中国化肥信息(2018年12期)2018-03-01
新农业(2017年24期)2018-01-30
