
茎瘤芥BjuGAPC基因序列特征及其参与茎发育的糖酸含量调控
2022-02-06 02:07李晓燕李成双金业程魏小涵李梦瑶
江苏农业学报 2022年6期
李晓燕, 李成双, 金业程, 魏小涵, 李梦瑶
(四川农业大学园艺学院,四川成都611130)
茎瘤芥(Brassicajunceavar.tumidaTsen et Lee)属于十字花科,是芸薹属芥菜种的变种之一,也是榨菜生产的主要原料[1]。茎瘤芥的产品器官为膨大的瘤状茎,既可鲜食,又宜加工,具有极高的营养和经济价值。茎瘤芥的生长和发育过程受到外部条件、内源激素等共同调控[2-3]。已有研究结果证明,许多关键基因可以调控果实发育过程中有机酸和糖代谢的合成[4],但在茎瘤芥茎膨大过程中有关糖酸含量的调控基因以及关键基因的生物信息分析较少。因此,结合茎瘤芥茎膨大过程中糖酸含量的变化,克隆与之相关的基因并研究其表达模式与调控作用,能够给茎瘤芥的分子育种和改良茎瘤芥品种提供一定的理论依据。
在植物体内,糖不仅是能量代谢的物质来源,同时可调控植物生长发育和环境应答[5]。果蔬及其制品中糖和有机酸的种类、数量以及糖酸比会影响其风味品质。可溶性糖是淀粉合成的底物,也直接关系到变态茎的营养状况,还原糖是蔗糖、淀粉等合成过程中光合作用的必需物质。甘油醛-3-磷酸脱氢酶(GAPDH)是高等植物糖酵解和糖异生反应中的关键酶,大致可以归为2种类型,一类是NADP+-GAPDH,参与植物的卡尔文循环[6],另一类是NAD+-GAPDH,参与糖酵解和糖异生过程[7]。
过去认为GAPDH基因在所有植物组织中几乎都是高水平表达且表达量相对稳定,所以常被用作研究其他功能性基因表达的内参基因[8]。然而,新的研究结果表明,GAPDH是一种多功能酶[9],除了参与植物基础的新陈代谢外,也参与逆境胁迫下的抵御反应[10-11]。GAPDH还广泛参与植株的生长发育进程,如拟南芥GAPDH基因缺失突变体表现为根不能正常生长和花粉败育[12-13],生理型雄性不育小麦发育过程中种子在不同生长发育阶段GAPDH基因表达量存在显著差异[14],拟南芥gapc1-1、gapc2-1基因双敲除突变体的种子含油量与敲除前相比下降3%[15]。
当前已有多种植物的GAPDH基因被克隆,如拟南芥[16]、番茄[17]、水稻[18]等,但是在芥菜中还未见相关报道。本研究基于芥菜的基因组和转录组数据库[19],克隆芥菜GAPDH基因,利用生物信息学方法对其编码的蛋白质氨基酸组成、蛋白质结构及系统进化等方面进行全面预测和分析,并结合茎瘤芥不同膨大时期的糖酸含量变化,对BjuGAPC基因的表达进行相关性分析,为阐明该基因的表达在茎瘤芥茎膨大过程中的调控作用奠定基础。
1 材料与方法
1.1 试验材料
植物材料为涪杂2号茎瘤芥,采自四川农业大学(成都校区)第五教学实验楼楼顶大棚。从瘤茎发育始期(2021年12月)开始取样,横径间隔2 cm取样1次,每次取 3 个生物学重复,共取样 4次。取样完立即放入冰盒带回实验室,所取样品用清水将杂质除净,晾干水分,用液氮速冻后放于-80 ℃冰箱中保存。
1.2 试验方法
1.2.1 茎瘤芥总RNA的提取和反转录 利用十六烷基三甲基溴化铵(CTAB) 试剂盒[生工生物工程(上海)股份有限公司]提取茎瘤芥总RNA,利用TSINGKE公司研发的GoldenstarTMRT6 cDNA Synthesis Mix反转录试剂盒合成cDNA第一链。
1.2.2BjuGAPC基因的克隆 利用Primer 6.0软件设计用于克隆BjuGAPC基因编码区全长和检测该基因表达量的引物(表1)。以芥菜的cDNA 为模板进行克隆,扩增程序为:95 ℃预变性3 min;95 ℃ 30 s,53 ℃ 30 s, 72 ℃ 10 s,35 个 循 环;72 ℃ 延 伸 5 min。采用1%琼脂糖凝胶电泳检查条带大小,对大小正确的条带切胶后用DNA凝胶回收试剂盒(OMEGA公司)对PCR产物进行纯化回收,连接转化DH5α大肠杆菌感受态细胞,挑选菌落PCR检测,将筛选到的阳性菌落送至北京擎科生物科技有限公司进行测序验证。

表1 BjuGAPC基因引物信息
1.2.3BjuGAPC基因在瘤茎膨大不同时期的表达分析 以茎瘤芥不同发育阶段的cDNA为模板,以芥菜TUB基因为内参基因[20]。反应程序:95 ℃预变性3 min;95 ℃变性5 s,53 ℃ 退火30 s,70 ℃延伸10 s,35个循环;72 ℃ 延 伸 5 min。用2-△△Ct方法计算BjuGAPC基因的相对表达量。
1.2.4BjuGAPC基因的生物信息学分析 首先使用美国国家生物技术信息中心(NCBI)在线工具ORF finder查找BjuGAPC基因的开放阅读框。BjuGAPC蛋白的氨基酸组成和理化性质分析用在线分析工具ProtParam来完成。用在线分析软件NetPhos3.1Server分析磷酸化位点和激酶特异性。采用糖基化位点在线预测软件YinOYang1.2Server和NetNGlyc1.0Server对BjuGAPC基因编码氨基酸的糖基化位点进行预测分析。利用ProtScale在线工具对BjuGAPC基因编码的氨基酸序列进行亲水性和疏水性分析。利用软件PSORT Prediction分析BjuGAPC的亚细胞定位。用TMHMM在线工具预测BjuGAPC蛋白的跨膜结构域。用SignalP在线工具预测分析BjuGAPC蛋白中的信号肽。使用SOPMA和SWISS-MODEL在线工具预测蛋白质的二、三级结构。BjuGAPC基因编码蛋白质的保守结构域分析通过Pfam在线软件进行。在NCBI的蛋白质序列数据库中进行BLASTP,并用DNAMAN软件进行氨基酸多重序列比对,用MEGA7构建系统进化树[21]。
1.2.5 糖酸含量的测定及统计分析 选择长势良好且一致的试验材料,测定不同时期的糖酸含量,均进行3个生物学重复。可溶性糖、葡萄糖、蔗糖、果糖含量采用试剂盒测定,蔗糖试剂盒购自北京索莱宝有限公司,其他试剂盒购自南京建成生物工程研究所。试验数据采用Excel和 SPSS 22.0进行统计分析。
2 结果与分析
2.1 BjuGAPC基因全长cDNA的克隆
以茎瘤芥涪杂2号cDNA为模板,设计特异性引物进行PCR扩增,扩增得到大小正确的条带,将其命名为BjuGAPC基因(GenBank登录号:OM100056),见图1。通过ORF Finder分析发现,茎瘤芥GAPDH基因包含1个1 038 bp的完整开放阅读框,编码345个氨基酸以及包含1个终止密码子。
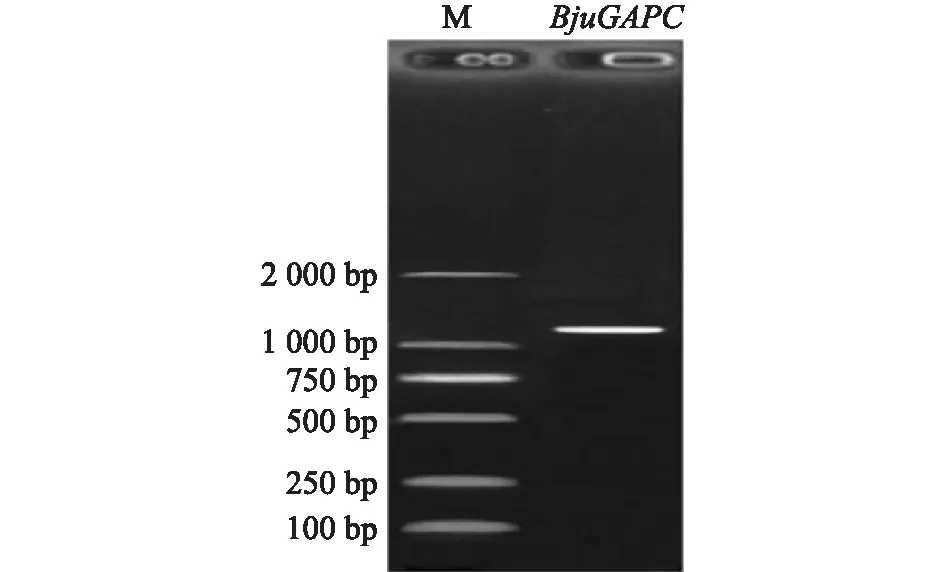
图1 BjuGAPC基因扩增电泳结果Fig.1 Electrophoretic results of BjuGAPC gene amplification
2.2 BjuGAPC蛋白的理化性质预测
利用ProtParam对BjuGAPC蛋白的理化性质进行分析,结果显示,组成BjuGAPC 蛋白的氨基酸共有20种,其中缬氨酸(Val)所占比例最高,为11.3%,半胱氨酸(Cys)所占比例最低,为0.6%(图2)。BjuGAPC 蛋白的分子式为C1 684H2 675N453O506S10,氨基酸数为 345,脂肪族氨基酸指数为 87.51,相对分子质量为3.768×104;理论等电点为 7.10,属于碱性蛋白质类;从不稳定系数为19.97和总平均亲水性为-0.161来看,BjuGAPC蛋白属于稳定、亲水性蛋白质。
2.3 BjuGAPC蛋白的糖基化位点预测和磷酸化位点预测
糖基化位点预测结果表明,BjuGAPC蛋白有7个O-GlcNAc糖基化位点(图3A)和2个N-糖基化位点(图3B),7个O-GlcNAc糖基化位点分别在第152位、287位丝氨酸 (Ser)和第191位、215位、288位、341位、344位苏氨酸(Thr)处,2个N-糖基化位点分别在153~155 aa(天冬氨酸-丝氯酸-丙氨酸,N-A-S) 和342~344 aa(天冬氨酸-组氨酸-苏氨酸,N-H-T) 处。
磷酸化位点和激酶特异性分析结果(图3C)表明,BjuGAPC蛋白存在33个潜在的磷酸化位点,包括18个丝氨酸(Ser)、11个苏氨酸(Thr)和4个酪氨酸(Tyr)磷酸化位点。根据上述磷酸化位点所对应的磷酸激酶预测结果,该蛋白质可能有10个蛋白激酶C(PKC)、5个酪蛋白激酶Ⅱ(CKⅡ)、4个周期蛋白质依赖性蛋白激酶(cdc2)、2个蛋白激酶G(PKG)、1个DNA依赖性蛋白激酶(DNAPK)、1个核糖体S6激酶(RSK)和1个酪蛋白激酶Ⅰ(CKⅠ)等7种保守的特异性蛋白激酶的结合位点。
蛋白质氨基酸序列的亲水性和疏水性分析结果(图3D)表明,该蛋白质的第340位脯氨酸(Pro)分值最低,亲水性最强;第164位亮氨酸(Leu)分值最高,疏水性最强。从总体上来看,BjuGAPC蛋白的亲水氨基酸数量稍多于疏水氨基酸,所以BjuGAPC基因编码的蛋白质最终表现为亲水性。
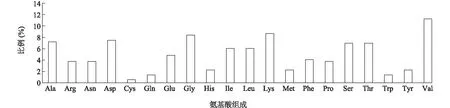
Ala:丙氨酸;Arg:精氨酸;Asn:天冬酰胺;Asp:天冬氨酸;Cys:半胱氨酸;Gln:谷氨酰胺;Glu:谷氨酸;Gly:甘氨酸;His:组氨酸;Ile:异亮氨酸;Leu:亮氨酸;Lys:赖氨酸;Met:甲硫氨酸;Phe:苯丙氨酸;Pro:脯氨酸;Ser:丝氨酸;Thr:苏氨酸;Trp:色氨酸;Tyr:酪氨酸;Val:缬氨酸。图2 BjuGAPC蛋白的氨基酸序列组成Fig.2 Amino acid sequence composition of BjuGAPC protein

A:O-GlcNAc糖基化位点预测;B:N-糖基化位点预测;C:激酶特异性预测;D:亲疏水性预测。图3 BjuGAPC蛋白的糖基化位点、磷酸化位点预测和亲疏水性预测Fig.3 Prediction of glycosylation site, phosphorylation site, hydrophilicity and hydrophobicity of BjuGAPC protein
2.4 BjuGAPC蛋白的信号肽、跨膜结构域和亚细胞定位预测
信号肽分析结果(图4A)表明,BjuGAPC基因编码的蛋白质序列中不存在已知的信号肽。跨膜结构域预测结果(图4B)表明,在整个BjuGAPC氨基酸序列中,没有发现可以与膜结合的区域或者跨膜结构,因此推测该蛋白质属于非跨膜蛋白质。结合信号肽预测分析结果可知,跨膜结构和信号肽的缺失说明BjuGAPC蛋白不是分泌蛋白质,而是一种由游离核糖体合成再进入细胞质的蛋白质。因此推测BjuGAPC蛋白可能定位于细胞质。
亚细胞定位分析结果表明,BjuGAPC基因最有可能在微体和细胞质中发挥功能,有极小可能定位于叶绿体基质和叶绿体类囊体薄膜中,这与信号肽预测结果一致。由此分析,BjuGAPC蛋白可能在细胞质中合成,在糖酸合成过程中有一定的调控作用。

A:BjuGAPC蛋白的信号肽预测;B:BjuGAPC蛋白的跨膜结构预测。图A中C代表剪切位置分值,S代表信号肽分值,Y代表综合剪切位置分值。图4 BjuGAPC蛋白的信号肽预测和跨膜结构域预测Fig.4 Prediction of signal peptide and transmembrane domain of BjuGAPC protein
2.5 BjuGAPC蛋白的二、三级结构预测
蛋白质二级结构预测结果表明,BjuGAPC蛋白的二级结构有α-螺旋、β-折叠、延伸链和无规则卷曲4种。其中构成α-螺旋(Hh)结构的有111个氨基酸,占比32.17%;构成β-折叠(Tt)结构的有22个氨基酸,占比6.38%;构成延伸链(Ee)结构的有83个氨基酸,占比24.06%;构成无规则卷曲(Cc)结构的有129个氨基酸,占比37.39%。
BjuGAPC蛋白三级结构的预测结果(图5)显示,用于建立其三级结构模型的氨基酸残基位于第5~338位,覆盖度高达96%,表明BjuGAPC蛋白能够形成同源四聚体,并且结合4个NAD+配体。
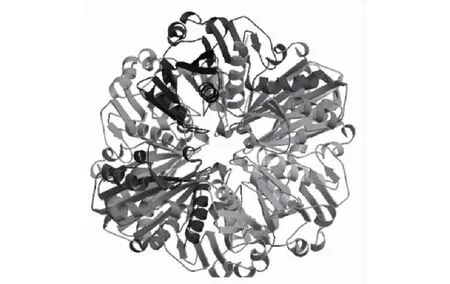
图5 BjuGAPC蛋白的三级结构预测Fig.5 Prediction of tertiary structure of BjuGAPC protein
2.6 BjuGAPC的蛋白质结构域分析和氨基酸序列的同源性分析
由蛋白质结构域分析结果(图6A)可知,BjuGAPC蛋白上有2个高度保守的超家族结构域Gp_dh_N(NAD+结合结构域)和Gp_dh_C(C端甘油醛-3-磷酸脱氢酶亚家族结构域),分别位于第6~109位和第161~318位氨基酸残基,表明BjuGAPC基因属于GAPDH基因家族。
利用BjuGAPC的氨基酸序列在NCBI上进行BLASTP,结果显示BjuGAPC蛋白的氨基酸序列与芜菁(XP_009125005.2)、油菜(XP_013742255.1)、萝卜(XP_018432846.1)、番茄(NP_001266254.2)、莴苣(XP_023733724.1)、马铃薯(NP_001275344.1)、拟南芥(NP_172801.1)的GAPDH基因编码的氨基酸序列具有高度相似性,其中与同科的芜菁、 油菜、萝卜的序列相似度高达98%以上,与其他物种的序列相似度均为92%以上。采用 DNAMAN软件将上述7个物种的GAPDH与BjuGAPC的蛋白质氨基酸序列进行多重比对,结果(图6B)显示:芥菜BjuGAPC的氨基酸序列与其他物种的GAPDH基因编码的氨基酸序列相似度极高。表明BjuGAPC同源蛋白质具有相似性和保守性,尤其表现在Gp_dh_N和Gp_dh_C两个保守结构域,因此猜测它们之间可能有着相同的生物学功能。
2.7 BjuGAPC蛋白的系统进化分析
为了探究BjuGAPC蛋白质的系统进化关系,从GenBank数据库中选取了19个物种的GAPDH蛋白氨基酸序列。在MEGA 7中用邻接法(Neighbor-joining)对20个蛋白质氨基酸序列进行分析,设置校验bootstrap=1 000,构建系统进化树(图7)。

A:BjuGAPC蛋白的结构域预测;B:BjuGAPC与其他植物GAPDH氨基酸序列比对。图中黑色方框中的区域为Gp_dh_N结构域,灰色方框中的区域为Gp_dh_C结构域。Consensus:共有序列。图6 BjuGAPC蛋白的结构域预测和同源性分析Fig.6 Domain prediction and homology analysis of BjuGAPC protein
结果表明,BjuGAPC蛋白与同为芸薹属的芜菁(Brassicarapa)的亲缘关系最近,其次是甘蓝(Brassicaoleracea)和油菜(Brassicanapus),与同为十字花科的高山南芥(Arabisalpina)、白芥(Sinapisalba)、萝卜(Raphanussativus)的亲缘性也较高,与罂粟(Papaversomniferum)、水青树(Tetracentronsinense)、松蒿(Phtheirospermumjaponicum)、博落回(Macleayacordata)等其他物种的关系较远。
2.8 不同时期瘤状茎糖酸含量变化与 BjuGAPC基因表达的关系
分别以膨大到横径为2 cm、4 cm、6 cm、8 cm(S1、S2、S3和S4时期)的茎瘤芥瘤茎为材料,测定不同时期瘤状茎糖酸含量、糖酸比。结果(表2)表明:糖酸的含量变化随着瘤状茎的发育有所不同,在发育前期茎中可溶性糖大量积累,还原糖(葡萄糖和果糖)含量较低;在发育后期,蔗糖和还原糖在茎中均有积累现象,而糖酸比整体出现先降低后升高的趋势。

图7 基于20个物种GAPDH蛋白的氨基酸序列构建的系统进化树Fig.7 Phylogenetic tree based on amino acid sequences of GAPDH proteins in 20 species
由BjuGAPC基因的转录组数据(图8A)和qRT-PCR分析得出的基因相对表达量(图8B)看出:转录组数据和qRT-PCR分析结果呈现一致趋势,BjuGAPC基因在瘤茎膨大过程中不同时期的相对表达量不同,且差异达到显著水平;随着瘤茎的逐步膨大,BjuGAPC基因的相对表达量逐渐减少,在瘤茎横径膨大到6 cm时基因的相对表达量降至最低值,之后小幅度上升。
相关性分析结果(表3)显示,可溶性糖含量与糖酸比呈显著正相关关系,可滴定酸含量与糖酸比呈显著负相关关系,基因的相对表达量与可溶性糖含量、可滴定酸含量和糖酸比均呈显著正相关关系。结果表明,BjuGAPC基因对不同时期瘤状茎糖酸含量起到一定的正向调控作用,且在横径为2 cm、4 cm时相对表达量较高,横径为6 cm、8 cm时相对表达量较低。
由此可见,BjuGAPC基因在瘤茎膨大的不同时期有明显的表达特异性,推测其在茎瘤芥茎生长发育过程中具有特定的表达模式,并在一定程度上影响了糖酸比。BjuGAPC蛋白在该过程中也发挥一定的功能,主要在瘤茎膨大前期起作用。
3 讨论与结论
为进一步了解GAPDH在茎瘤芥茎膨大过程中的作用,本试验基于芥菜的基因组数据库,克隆得到BjuGAPC基因。预测BjuGAPC蛋白相对分子质量为3.768×104,是一个具有碱性、亲水性的稳定蛋白质。GAPDH是高等植物糖酵解反应中的关键酶,能够维持植物的生命活动,按照生化特性可分为磷酸化GAPDH和非磷酸化GAPDH两大类[22]。通过分析BjuGAPC蛋白的磷酸化位点,发现该蛋白质存在33个潜在的磷酸化位点,推测其可能参与糖酵解途径,对糖酸合成起到一定的调控作用。

表2 不同采样时期茎瘤芥瘤状茎的糖酸含量

S1、S2、S3和S4时期分别指瘤状茎膨大到横径为2 cm、4 cm、6 cm、8 cm的时期。不同时期对应的柱上标有不同小写字母表示差异显著(P<0.05)。图8 BjuGAPC基因不同时期的表达丰度(A)与 qRT-PCR表达分析(B)Fig.8 Expression abundance(A) and qRT-PCR expression analysis (B)of BjuGAPC gene at different stages

表3 不同时期茎瘤芥瘤状茎糖酸含量与基因表达量间的相关性
蔗糖是植物生长发育过程中各种调控机制的信号,它还影响编码转运蛋白、贮藏蛋白和应激反应的基因的表达[23-27]。在瘤状茎发育期间,在发育前期茎中蔗糖大量积累,还原糖含量较低。在发育后期,蔗糖和还原糖在茎中均有积累现象。这说明BjuGAPC基因在前期相对表达量较多,同时由于叶片中还原糖能较快地转化为蔗糖,所以能迅速输送到茎中用于其他物质的合成,而在发育后期,由于植株机体机能逐渐衰退,BjuGAPC基因相对表达量降低,瘤状茎中其他物质合成能力减弱,合成的蔗糖减少,还原糖含量相对下降。
根据磷酸化的GAPDH在细胞中的不同定位可将其分为GAPC、GAPCp和GAPA/B三类[28-29]。试验中发现BjuGAPC的蛋白质序列中不存在已知的信号肽,并且没有任何跨膜结构或膜结合区域,说明该蛋白质不是分泌蛋白,不经过跨膜转运,而是在细胞质中直接形成,亚细胞定位预测结果也证实了这一点。因此,BjuGAPC蛋白属于GAPC,定位于细胞质中。从系统进化树可以看出,分支点越接近,物种间的亲缘性越相似,说明编码这些蛋白质的基因间的功能也具有一定的相似性,同时说明GAPDH基因家族蛋白质遗传进化比较保守。BjuGAPC蛋白之所以能行使生物学功能是因为其特定的空间结构。通过三级结构预测发现,BjuGAPC蛋白能够形成同源四聚体,并且结合4个NAD+配体。由蛋白质结构域分析结果可知,BjuGAPC的2个超家族结构域Gp_dh_N和Gp_dh_C中,前者是辅酶NAD+的结合域,后者是行使糖运输和代谢的功能域,推测BjuGAPC基因的表达产物在茎瘤芥细胞质中参与糖酵解过程,符合上述所推测的BjuGAPC的结构和功能特征。
以前的多种研究结果普遍表明,GAPDH基因在同一组织的不同生理状态下或同一生物体的不同组织中均相对稳定且高水平表达,因此可以作为内参基因来进行其他功能基因的表达差异分析。但是,本研究通过qRT-PCR技术检测发现,BjuGAPC基因在茎瘤芥茎膨大不同时期有明显的表达特异性,不同时期糖酸比测定结果显示,BjuGAPC基因相对表达量随不同时期变化与糖酸比含量呈现出一致趋势,说明BjuGAPC基因参与了茎瘤芥的生长发育过程,并且主要在瘤茎膨大前期发挥功能。GAPDH基因可以作为多物种分子生物学研究的内参基因,但是在本试验的芥菜发育过程中表达差异较大,这可能是由于物种不同导致的特异性差异所决定的。
在对BjuGAPC蛋白的磷酸化位点分析中,发现了植物抗逆相关特异性蛋白质激酶的结合位点,也可以说明GAPDH在功能上与植物的生长发育密切相关。一些研究者的转基因试验结果也表明,GAPDH缺失或过表达均会在一定程度上影响植物的代谢和生长发育[30],并且GAPDH基因的表达水平会随着内外源诱导因子的变化而变化,推测BjuGAPC蛋白中参与信号转导和激素调控等的O-GlcNAc糖基化位点和N-糖基化位点与此有关[31-32]。糖酸含量分析结果表明,BjuGAPC蛋白参与了糖代谢过程。目前,有关芥菜GAPDH功能和作用机制的相关报道还很少,本试验对BjuGAPC基因的克隆可以为分析逆境胁迫条件下BjuGAPC基因的表达模式、探讨BjuGAPC在茎瘤芥茎膨大过程中的作用奠定基础,后续还可以进一步构建植物表达载体,通过转基因等途径深入探究BjuGAPC基因在逆境胁迫和茎瘤芥茎膨大过程中的功能。此外,本研究结合不同时期糖酸比变化趋势,验证了茎瘤芥BjuGAPC基因确实对糖酸含量起到正向调控作用。
猜你喜欢
湖北农业科学(2022年11期)2022-07-18
江苏农业学报(2022年1期)2022-03-16
河北果树(2021年4期)2021-12-02
实用肿瘤学杂志(2020年4期)2020-12-08
山东农业科学(2020年1期)2020-07-04
中国洗涤用品工业(2019年4期)2019-05-11
中成药(2018年1期)2018-02-02
中成药(2017年3期)2017-05-17
天津科技大学学报(2016年1期)2016-02-28
山西果树(2014年6期)2015-03-12
