
温州蜜柑完熟栽培期间果实糖酸变化及其相关基因表达分析
2022-02-04 11:51易明亮刘德春匡柳青
江西农业大学学报 2022年1期
易明亮,肖 洒,刘德春,杨 莉,匡柳青,刘 勇*,胡 威*
(1.江西农业大学 农学院,江西 南昌 330045;2.江西省井冈蜜柚繁育与栽培技术工程中心 江西农业大学分中心,江西 南昌 330045;3.四川省合江县农业农村局,四川 泸州 646200)
【研究意义】宽皮柑橘种植方式以露地栽培为主,往往导致其成熟上市期较为集中,产品过剩,造成增产不增收现象。与露地栽培模式相比,设施完熟栽培不仅可以有效调节鲜果的成熟上市期,而且显著提高果实品质[1],从而提升产品价格使果农获得更高的经济效益。温州蜜柑果实品质主要由蔗糖及柠檬酸等决定[2],而糖酸代谢机制较为复杂,由一系列酶以及相关基因控制。因此研究设施完熟栽培过程中果实糖酸变化规律及与相关基因表达的关系,对了解完熟栽培过程中果实品质提升的分子机制,以及发展高质量优质栽培技术及选育高品质柑橘品种具有重要意义。【前人研究进展】对果实糖酸代谢规律及机制已有大量研究,主要集中在果实发育期糖酸代谢相关酶活性及酶基因的表达上。柑橘果实中蔗糖的积累主要由蔗糖磷酸合成酶(Sucrose Phosphate Synthase,SPS)及蔗糖合成酶(Sucrose Synthase,SUS)控制。研究表明CitSPSs基因对金柑果实中蔗糖的合成和积累具有重要作用[3]。Komatsu等[4]分析了温州蜜柑中3个CitSPS基因的表达模式,发现果实可食部分中CitSPS1的表达量相对较高,而CitSPS2和CitSPS3表达量则相对较低。在香蕉果实成熟过程中,蔗糖的累积与淀粉的降解与SPS基因mRNA 的富集密切相关[5]。Islam 等[6]研究了温州蜜柑基因组中6个CitSUSs基因在果实发育期间的表达模式,在不同组织中各基因的表达模式有明显差异,汁胞中CitSUS1、CitSUS2和CitSUS5基因的表达明显高于其他组织。柠檬酸是柑橘果实的主要有机酸,其含量由柠檬酸合成及降解相关酶及基因共同控制。林琼[7]的研究结果表明CitPEPCs和CitCSs基因的上调表达导致了露地栽培的椪柑柠檬酸含量显著高于温室栽培果实,而降解酶相关基因CitGAD4和CitAco3的表达水平对维持椪柑细胞内有机酸平衡具有重要作用。陈明[8]的研究结果显示CitAco3、CitGAD4/5、CitGS2的转录丰度在低酸早熟椪柑果实中均显著高于高酸的普通椪柑。【本研究切入点】柑橘糖酸积累机制的研究多集中在果实发育期或正常采收前的时期,并且设施完熟栽培的温州蜜柑前期研究主要集中在栽培技术的改进以及果实品质的变化规律上,鲜有报道在设施完熟栽培过程中果实糖酸的变化规律与相关代谢基因表达的关系。【拟解决的关键问题】本研究测定了设施完熟栽培过程中温州蜜柑‘宫川’果实糖及有机酸的含量变化,以及与糖酸相关代谢基因表达量的变化,以期明确设施完熟栽培过程中果实糖酸变化规律及与相关基因的表达关系,进一步完善完熟栽培过程中果实品质提升的分子机制。
1 材料与方法
1.1 植物材料
‘宫川’温州蜜柑(Citrus unshiuMarc.)种植于江西省吉安市新干县腊月红生态果业有限公司,树龄7 年。在2017 年11 月至2018 年1 月间,于设施完熟栽培期间,即盛花后(DPA)220,230,240,250,260,270,280 d 收集果实样品,以3 棵健康树为1 个重复,共设置3 个重复,每棵树从东、西、南、北4 个方向各取5 个果实,样品采集后带回实验室,一部分样品用于品质分析,一部分果肉样品置于液氮冷冻后保存于-80 ℃冰箱中用于后续基因表达分析试验。
1.2 测定项目及方法
1.2.1 可溶性总糖及各组分含量测定 可溶性总糖采用蒽酮比色法进行测定[9]。糖的各组分测定采用高效液相色谱(HPLC)法(日本岛津LC-20AT)进行检测,测定参照刘灵智[10]的方法。
1.2.2 可滴定酸及各有机酸组分含量测定 可滴定酸采用酸碱滴定法[9],并换算成柠檬酸含量。各有机酸含量采用高效液相色谱法,并参照林琼[7]的方法进行测定。
1.2.3 RNA 提取与反转录 果肉用液氮磨碎后采用Trizol 法提取[11]。RNA 提取质量用琼脂糖凝胶电泳进行检测,并使用Nanodrop2000微量分光光度计测定RNA 浓度。反转录采用伯乐iScript cDNA 试剂盒,得到的cDNA用于荧光定量分析。
1.2.4 基因相对表达量分析 应用Bio-Rad 的CFX96 定量PCR 仪,Ssofast EvaGreen Supermix(Bio-Rad)定量分析试剂盒对不同采样时期的果肉中所研究的基因进行定量表达分析,内参基因(Actin)和糖酸代谢相关基因的qPCR引物见表1。用2-ΔΔCt法计算基因相对表达量,每样品设3次生物学重复。

表1 实时荧光定量PCR 引物Tab.1 Primer sequences of real-time quantitative PCR
1.2.5 数据统计与分析 数据统计及整理采用Office365 Excel 软件,数据分析利用SPSS 软件进行Duncan 式方差分析以及Pearson相关性分析,最后采用OriginPro 2021软件进行图形的制作。
2 结果与分析
2.1 ‘宫川’完熟过程中果实糖酸变化
花后220 d 已是露地常规栽培‘宫川’温州蜜柑果实采收期,而设施完熟栽培的果实还需40~60 d 采收。图1A 显示完熟栽培的果实可溶性总糖含量持续上升,至花后260 d 时达到最高为13.2%,之后缓慢下降。利用高效液相色谱法分析果实中各种糖组分在不同时期的含量,结果显示蔗糖是果实中主要的糖组分,占总糖含量65%左右,其次是果糖和葡萄糖,而蔗糖含量的变化趋势也与总糖变化趋势一致,在花后260 d 达到最高,为70.38 mg/g;而果糖和葡萄糖含量在整个完熟栽培期间(花后220~280 d)相对较为稳定(图1C),因此说明温州蜜柑果实中总糖的变化主要由蔗糖含量决定。
从花后220 d 开始设施完熟栽培的温州蜜柑果实中可滴定酸的含量总体呈下降趋势,在花后280 d达到最低仅为0.57%(图1B)。利用高效液相色谱法分析了‘宫川’果实中各有机酸组分的变化情况,在花后220 d时柠檬酸含量约为6.03 mg/g,之后逐渐下降,在花后280 d达到最低,为3.67 mg/g,在整个时期中柠檬酸含量占总有机酸的65%~80%,表明柠檬酸是果实中主要有机酸,而苹果酸、奎宁酸、酒石酸含量相对较低并且在完熟过程中变化不大,因此总酸的变化受柠檬酸含量变化的影响最大。

图1 完熟栽培期间果实中糖和酸含量的变化Fig.1 Changes of sugar and acid components in fruit during delayed harvest cultivation
2.2 蔗糖合成相关基因表达分析
蔗糖磷酸合成酶(SPS)是柑橘蔗糖合成途径中一个重要的调节酶,在蔗糖合成途径中起主要作用。本研究中检测了设施完熟采收期间CitSPS1和CitSPS2基因在果肉中表达量的变化,结果表明(图2)、Cit-SPS1和CitSPS2基因表达量均在花后220~240 d表现出先上升后下降的趋势,并在花后250 d显著上升,CitSPS1在花后250~280 d 维持在相对较高的水平,而CitSPS2基因表达量则表现出先上升后下降的趋势,在花后260 d表达量达到最高值,之后表达量开始显著下降。
蔗糖合成酶基因CitSUS1、CitSUS2、CitSUS3、CitSUS4、CitSUS5以及CitSUS6在设施完熟栽培期间果肉中的表达量均进行了检测,其中CitSUS2和CitSUS5基因表达量极低未能检测出;CitSUS1基因表达量在花后220~250 d 期间表达量逐渐上升,250 d 后显著下降并维持在相对稳定的水平;而CitSUS3和Cit-SUS4基因表达量在花后220~250 d 期间也表现出上升趋势,之后保持在较高水平;CitSUS6基因在花后240 d 前表达量呈上升趋势,之后维持在较高水平,至花后280 d 时显著下降,达到最低表达水平(图2)。由此表明CitSUS3、CitSUS4以及CitSUS6基因的表达变化情况与蔗糖含量的变化情况较为相似,可能与果肉中蔗糖的积累密切相关。
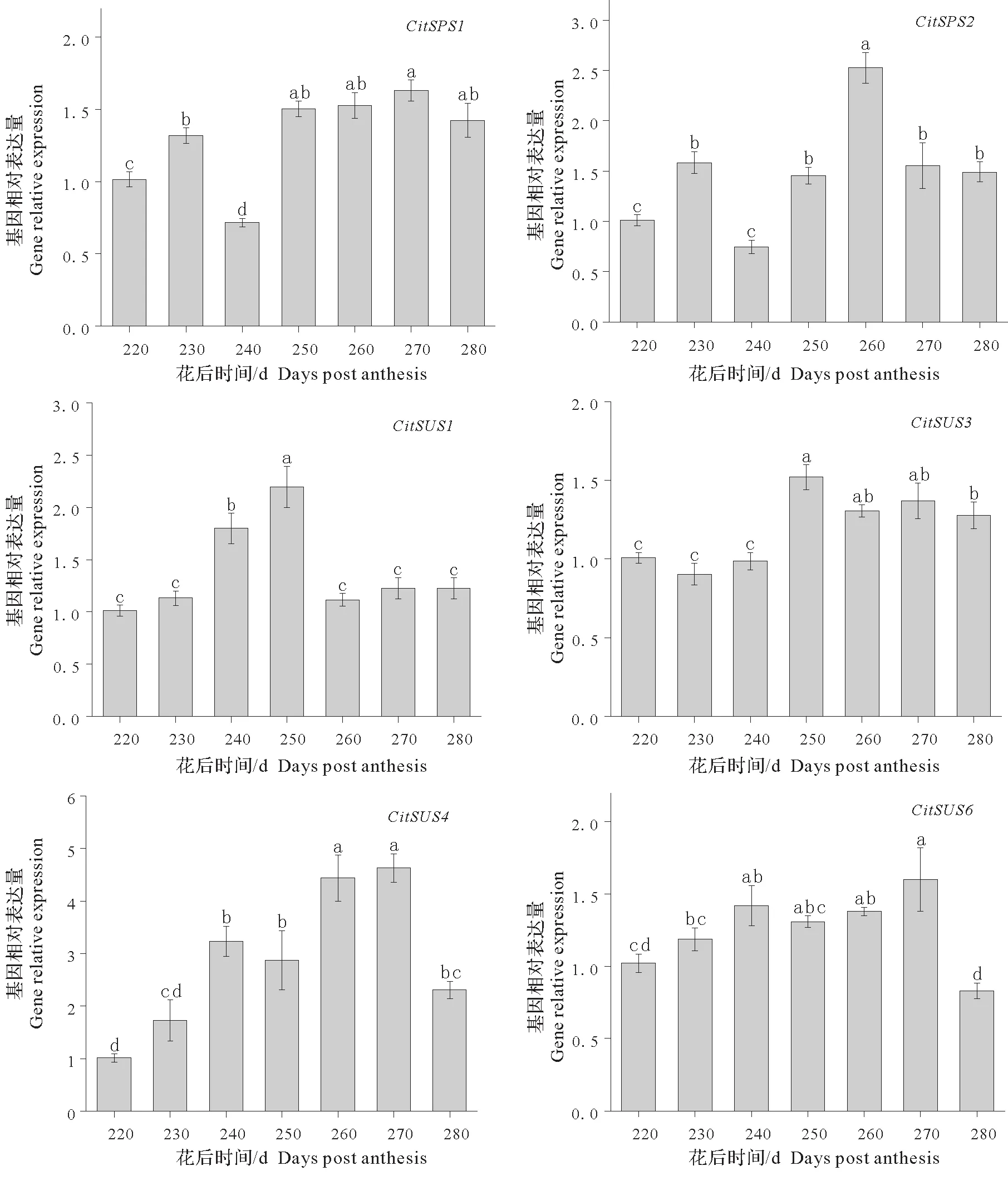
图2 果实完熟栽培期间中蔗糖合成基因的相对表达量Fig.2 Relative expression of sucrose synthesis gene in fruits during delayed harvest cultivation
2.3 柠檬酸合成相关基因表达量分析
柠檬酸磷酸烯醇式丙酮酸羧化酶基因(PEPC)和柠檬酸合酶基因(CS)均为柠檬酸合成的关键酶基因。由图3 可知果肉中CitPEPC基因表达量整体上呈先上升后下降的趋势,至花后260 d 时达到最高,260 d后则显著下降;而CitCS基因表达量在完熟栽培过程中同样呈先上升后下降趋势,其表达量在花后250 d 时达到最高,之后逐渐下降。完熟栽培过程中柠檬酸含量不断下降,而CitPEPC和CitCS基因表达量并未下降反而呈上升趋势,由此说明柠檬酸含量的下降与CitPEPC及CitCS基因并无明显关联,同时也表明完熟栽培阶段果实中柠檬酸含量虽然下降,但合成量并未减少。
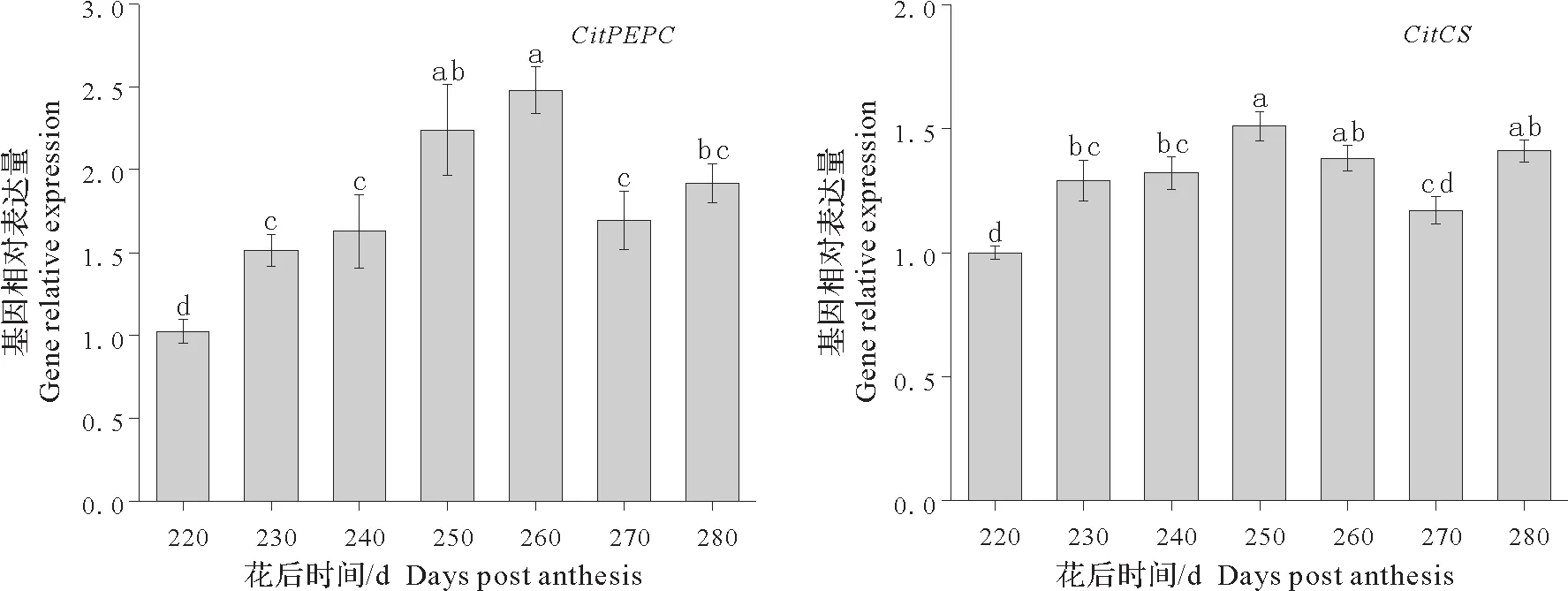
图3 果实完熟栽培期间柠檬酸合成相关基因的相对表达量Fig.3 Relative expression of citric acid synthesis gene in fruits during delayed harvest cultivation
2.4 柠檬酸降解相关基因表达分析
CitACO3、CitGAD4和CitGS2为柠檬酸代谢途径上的3 个关键酶基因,3 种基因的相对表达量变化如图4,果肉中CitACO3和CitGAD4基因表达量在整个完熟栽培过程中呈上升趋势,均在花后280 d 达到最高,并且CitGAD4基因在花后280 d时其表达活性相比于花后270 d大幅度增强。而柠檬酸含量在完熟栽培过程不断下降,说明CitACO3和CitGAD4基因的表达量增加可能对柠檬酸的下降具有关键作用。而CitGS2基因表达量变化波动较大,整体呈先上升后下降,再上升再下降的趋势,并无明显规律。

图4 完熟栽培期间果实中柠檬酸降解相关基因的相对表达量Fig.4 Relative expression of citric acid degradation gene in fruits during delayed harvest cultivation
2.5 糖酸含量与代谢相关基因表达的相关性分析
对完熟栽培的果实总糖含量及柠檬酸含量与相关代谢基因的相对表达量进行了相关性分析,其中总糖指果实中蔗糖、果糖及葡糖糖含量的总和(表2),结果表明总糖含量与CitSUS1基因表达量相关性较弱,与CitSPS1、CitSPS2、CitSUS6基因呈中等程度正相关但不显著,而与CitSUS3、CitSUS4基因呈显著正相关,相关系数分别为0.756 和0.796,说明CitSUS3、CitSUS4基因在完熟栽培果实糖的积累中起到了关键性作用。果实柠檬酸含量与合成基因CitPEPC及CitCS表达量呈负相关,但不显著;与降解基因CitGS2表达量相关性较弱,而与CitGAD4表达呈显著负相关,与CitACO3表达量呈极显著负相关关系,相关系数分别为-0.810和-0.942,由此说明完熟栽培期间柠檬酸的积累量下降是由降解基因的持续上调表达决定的,而与合成基因的表达不相关。

表2 果实糖酸含量与代谢相关基因表达的相关系数Tab.2 Correlation coefficient between total sugar and citric acid content and metabolism-related gene expression of fruit
3 讨论
糖酸含量是决定柑橘果实品质、口感、风味的主要因素。果实汁囊中积累的可溶性糖大部分为蔗糖、果糖和葡萄糖,有机酸则以柠檬酸为主,并且随着果实的发育可溶性糖的含量不断上升,有机酸则在果实发育期初期不断积累而在成熟期则不断下降。赵智中等[12]的研究表明黄岩本地早和‘宫川’温州蜜柑果实在发育期均以积累蔗糖为主,并且在整个发育期含量不断上升。曾祥国[2]分析了来自不同产区的37个不同柑橘品种果实中糖酸含量,证实总糖中以蔗糖为主而有机酸则主要为柠檬酸,其比例分别达到了总糖的58%和有机酸的85%。本研究中‘宫川’温州蜜柑设施完熟栽培期间果实中总糖含量不断增加,至花后260 d时达最高,之后缓慢下降,而有机酸含量则持续下降,并且总糖中以蔗糖含量最高,其次是果糖和葡萄糖,有机酸中则以柠檬酸含量最高。因此研究蔗糖和有机酸代谢相关基因的表达对理解完熟过程中品质变化的分子机理具有重要意义。
柑橘果实中糖的合成由一系列的酶及相关基因调控,在果实不同发育阶段不同的酶及相关基因发挥不同的调控作用。刘永忠等[13]在研究脐橙果实糖积累与蔗糖代谢酶活性时发现蔗糖磷酸合成酶SPS和蔗糖合成酶SUS 对脐橙汁胞中糖组分的调节具有重要作用,并且在果实成熟期至采收期SUS 仍保持较高的活性。王成西等[14]的研究表明NAA可显著抑制葡萄果实中VvSPS和VvSUS基因的表达,同时果实中可溶性糖含量显著降低,经相关性分析表明,可溶性糖含量的降低与VvSPS基因表达有一定相关性但并不显著,而仅在不进行任何处理的对照中与VvSUS基因的表达有显著正相关关系。此外,张雅剑[15]的研究表明,在锦橙的发育过程中CsSPS基因主要果实成熟前期的糖积累过程中表达量相对较高,而Cs-SUS基因则在果实成熟后期对糖的积累起主要作用。以上研究结果均表明在果实成熟后期糖分的积累主要由SUS基因调控,而SPS基因主要在果实发育前期对糖分的积累起主要作用。‘宫川’温州蜜柑在设施完熟栽培期间,CitSPS1和CitSPS2基因与果实中总糖的积累有一定的相关关系,但相关性并不显著,而蔗糖合成酶基因CitSUS3和CitSUS4与总糖的积累分别有显著和极显著的相关关系,说明果实完熟期间糖分的积累主要由CitSUS3和CitSUS4基因调节。
柑橘果实中柠檬酸的合成是在磷酸烯醇式丙酮酸羧化酶(PEPC)的催化下,磷酸烯醇式丙酮酸(PEP)与CO2结合生成草酰乙酸(OAA),然后在柠檬酸合酶(CS)的作用下OAA 与乙酰辅酶A 缩合成柠檬酸,因此PEPC 和CS 活性与柠檬酸的累积密切相关。Deng 等[16]通过将香橙的CjCS基因在烟草中超量表达,显著增强了CS 酶在烟草中的活性,同时柠檬酸含量相对于野生型植株显著上升。低温可以显著诱导CitPEPCs和CitCSs基因的表达使果实中柠檬酸含量上升,因此椪柑的延后采收过程中,露地栽培的果实中CitPEPCs和CitCSs基因表达量均显著高于设施栽培,并且柠檬酸含量也显著高于设施栽培的果实,因而导致果实品质明显变差[7]。但也有研究证实柑橘果实在发育成熟过程柠檬酸的积累与合成相关基因的表达并无显著的相关关系。Chen 等[17]在脐橙的研究过程中发现整个果实发育过程中CitCSs基因的表达量较为稳定,对柠檬酸的积累不起关键作用。同时高阳等[18]的研究也表明在‘靖安椪柑’的果实发育过程中CitCSs和CitPEPCs基因的表达并不是调控果实柠檬酸积累的主要原因。笔者的研究结果表明,‘宫川’温州蜜柑在设施完熟栽培过程中果实柠檬酸含量不断下降,而CitCS和CitPEPC基因的表达量并未下降,反而呈上升趋势,并且相关性分析结果显示与柠檬酸含量呈负相关关系,这也与Lin 等[19]在研究椪柑完熟栽培过程柠檬味柠檬酸合成相关基因的变化趋势相似,因此说明CitCS和CitPEPC基因并不是温州蜜柑完熟栽培过程中调控柠檬酸含量的关键基因。
柑橘果实柠檬酸降解途径中的第一个关键酶是顺乌头酸酶(ACO),其催化柠檬酸转化为异柠檬酸,之后通过一系列生化反应生成谷氨酸盐后经γ-氨基丁酸(GABA)途径和谷氨酰胺途径降解,而谷氨酸脱羧酶(GAD)和谷氨酰胺合成酶(GS)则分别是这两条代谢途径上的关键酶。Terol等[20]研究了橘、甜橙、柠檬等柑橘种类中多个品种果实发育过程有机酸的变化规律及CcACOs基因的表达规律,证实在除柠檬外的大部分品种中CcACOs基因的表达能促进果实中有机酸的降解。并且CitACO2/ACO3的表达量增加是‘靖安椪柑’果实发育阶段后期柠檬酸下降的重要因素之一[18]。另外在椪柑叶片及果实中瞬时表达CitACO3基因及调控CitACO3基因的转录因子基因CitNAC62和CitWRKY1时,叶片及果实中的柠檬酸含量均显著降低[21]。前人研究认为柑橘中柠檬酸降解过程中GABA 途径占主导地位[22],之后Lin等[19,23]证实在椪柑的果实成熟过程中以及在低温诱导下柠檬酸的降解主要与CitACO3、CitGAD4基因的表达密切相关,说明柠檬酸主要是通过GABA 途径进行降解;另外Chen 等[24]以及Sheng 等[25]的研究也表明椪柑以及HB 柚与椪柑的杂交种果实在采后贮藏过程中其柠檬酸的降解也主要是通过GABA 途径进行。‘宫川’在设施完熟栽培过程中果实柠檬酸降解分别与CitACO3以及CitGAD4基因的表达呈极显著和显著的负相关关系,与CitGS2基因的表达相关性较弱,而CitACO3和CitGAD4基因是GABA 途径上的关键酶基因,由此说明柠檬酸也主要是通过GABA 途径进行降解,CitACO3和CitGAD4基因的表达决定了果实中柠檬酸的含量。
4 结论
本研究明确了‘宫川’温州蜜柑在设施完熟栽培期间果实糖酸的变化规律,果实中蔗糖为主要的糖组分而柠檬酸为主要有机酸,随着时间推移总糖含量不断升高后缓慢下降,而柠檬酸积累则不断减少。总糖的变化与蔗糖合成酶基因CitSUS3和CitSUS4的表达呈显著正相关,总糖含量由CitSUS3和CitSUS4基因的表达决定;而柠檬酸的积累与柠檬酸合成基因CitPEPC、CitCS的表达无直接的关系,主要受柠檬酸降解基因CitACO3、CitGAD4表达的影响,果实柠檬酸降解主要通过GABA途径进行。
致谢:江西省教育厅科学技术研究项目(GJJ170292)同时对本研究给予了资助,谨致谢意!
猜你喜欢
江西水产科技(2022年2期)2022-05-17
现代仪器与医疗(2021年1期)2021-06-09
食品界(2018年8期)2018-09-03
妇女之友(2017年8期)2017-10-24
科技与创新(2016年22期)2017-03-30
科学与财富(2016年34期)2017-03-23
江苏农业科学(2016年7期)2016-10-20
科技视界(2016年20期)2016-09-29
长江蔬菜·学术版(2014年12期)2015-01-08
科技与创新(2014年17期)2014-10-22
