
足球运动对运动员源定位及脑功能连接的影响研究
2022-02-03 02:27李垂坤张业廷魏翠兰李慧军袁琼嘉
中国体育科技 2022年11期
李垂坤,张业廷,魏翠兰,赵 祥,李慧军,李 佳,袁琼嘉
静息态是大脑在不执行具体认知任务、保持安静、放松、清醒的状态。静息态下大脑消耗的能量约占人体总耗能的20%,任务态下约为25%,增加约5%,说明大脑消耗的能量大部分用于维持静息态的神经元活动。大脑在静息态下,不同脑区的神经元也在协同工作,自发性的神经活动形成同步、结构化的功能网络(Beckmann et al.,2005; Fox et al., 2007)。静息态下的脑功能连接反映了大脑内在、固有的活动模式,是组成所有认知、感知和行为活动的核心,显示了神经集合兴奋性的持续和有组织的变化(季淑梅 等,2020;Roth et al., 2014)。
在运动协调和运动表现的背景下,静息态脑功能连接用于监测注意力和感觉-运动大脑区域的相互关联性,作为衡量体育活动准备状态和效率的指标。功能磁共振成像(functional magnetic resonance imaging, fMRI)研究发现,注意力相关的脑功能连接和感觉-运动脑功能连接的调节,揭示了运动引起的静息态脑功能连接变化(Schmitt et al., 2019)。
文献已证实,运动可以改善认知水平,诱导神经可塑性发生(Zilidou et al., 2018)。在大脑结构层面,优秀运动员的脑解剖在左楔前叶、左顶下小叶和右额叶3个区域的皮质厚度显著增加(Wei et al., 2020)。铁人三项健将级运动员的内侧前额叶皮层、中央前回和中央后回,以及岛叶皮质厚度增加(Wood et al., 2016)。运动员在右侧感觉运动区、边缘叶和小脑前叶表现出更大的灰质体积(Gao et al., 2019)。运动员在左楔前叶和右/左岛叶之间的脑功能连接增加(Kim et al., 2021)。不同运动训练对内在功能连接的精确定位不同。研究发现,职业羽毛球运动员在额叶和左顶上小叶之间的功能连接发生了改变(Jolles et al., 2013;Takeuchi et al., 2013)。另一项研究发现,职业篮球运动员在动作预期过程中,顶下小叶和额下回的活动更活跃(Wu et al., 2013)。这些改变是由于运动训练引起的,促进了相关区域之间的连接性,使大脑的结构和功能更具适应性。
脑电(electroencephalography, EEG)的静息态脑功能连接分析具有重要的科研价值,任何身体活动干预措施都可能导致皮质连接性的改变(Flodin et al., 2017;Jonas‐son et al., 2017)。基于体素的脑成像理论,了解足球运动训练对脑区之间相互作用,有助于更全面、深入地了解脑的可塑性以及训练对脑功能、脑连接的影响。进一步从脑功能连接的层面了解足球运动训练对脑神经机制的影响,有助于在未来的体育教育及运动干预中设计更有针对性的措施,从而提高足球运动训练效果。为此,本研究采用电磁断层成像(standardized low-resolution brain elec‐tromagnetic tomography analysis, sLORETA)在源水平上重构EEG信号,计算相应延迟指数(phase lag index, PLI)构建脑功能连接。分析静息态下各频段功率谱、源定位及脑功能连接的特征,探究足球运动训练对运动员静息态下与视觉注意和视觉运动协调加工整合相关脑区的影响。
1 研究对象与方法
1.1 研究对象
本研究招募成都足球学院国家二级及二级以上足球运动员和非专业运动员,比例为1∶1,总人数为72人。纳入被试标准:1)无严重躯体疾病,无脑创伤及神经系统疾病病史,且目前精神状况良好,无药物和酒精依赖史;2)右利手;3)视力或校正视力正常,无色盲和色弱;4)智力与焦虑水平正常。本研究的内容经过成都体育学院伦理委员会(成体伦理〔2020〕22号)审批,所有被试签署知情同意书。其中,5名被试的脑电数据质量较差,伪迹超过30%,未纳入统计。最终,本研究以67名学生(男生49名,女生18名)作为研究对象,年龄范围为17~22岁。依据训练年限和运动员等级分为:运动组33人(男27人,女6人),训练年限均为6年以上,国家二级及二级以上运动员;对照组34人(男22人,女12人),对足球运动有一定的了解,无运动等级的体育专业学生,且半年以上未进行大强度运动。
1.2 研究方法
1.2.1 静息态EEG数据采集
采用德国BP公司的actiCHamp 64通道脑电系统,被试保持安静、放松状态,始终注视正前方60 cm处显示器屏幕中央视平线处的“+”标记,尽量减少眨眼和眼动,采集5 min连续无间隔的睁眼静息态脑电信号。
1.2.2 静息态EEG数据预处理
基于Matlab开发的工具箱,使用EEGLAB脑电数据处理与分析软件对数据进行预处理,利用EEGLAB中默认的Standard-10-5-cap385文件进行电极定位;采用全脑平均参考,同时恢复FCz电极的通道信息;删除眼电等无用电极;采用0.5~100.0 Hz波段过滤脑电波(Mantini et al.,2007);每2 000个数据点分为1段;去除眨眼、头动、肌电等伪迹,并剔除电压幅值绝对值大于100 μV的数据段(Onton et al., 2006)。
1.2.3 静息态EEG功率谱密度
利用EEGLAB的STUDY模块计算各个频段功率谱,通过快速傅里叶变换(fast Fourier transform,FFT)计算功率谱(蒋园园 等, 2020;García-Monge et al., 2020)。将脑电信号频带分为 δ(0.5~4.0 Hz)、θ(4~8 Hz)、α(8~14 Hz)、β(14~30 Hz)、γ(30~48 Hz)5 个频段。功率谱密度采用相对功率谱密度(power spectral density,PSD),以消除个体差异的影响(Gray, 1994)。统计分析采用SPSS 18.0,进行独立样本t检验,显著性水平P<0.05。使用Graph‐Pad Prism 8和Adobe Illustrator CS 5软件进行图片制作。
1.2.4 静息态EEG功率源定位
采用sLORETA进行溯源分析(Pascual-Marqui et al.,2002a, 2002b)。假设大脑是一个由脑内组织、颅骨、头皮3层组织构成的头模型,基于线性和叠加原理,在配准的Talairach脑模板下,通过估计最大电流密度准确定位独立源的三维坐标,并以蒙特利尔神经病学研究所(Montreal Neurological Institute,MNI)152坐标系(5 mm分辨率)展现溯源结果(Mazziotta et al., 2001)。脑组织被数字化标定为6 239个立体像素,每个像素都对应1个三维坐标,同时也会有1个电流密度值。
对EEG信号经过预处理后,将头皮脑电电极MNI坐标转换为 Talairach 坐标(Talairach et al., 1988),用 Talai‐rach坐标创建转换矩阵。根据分段的txt文件计算EEG频段的交叉谱生成相对应的sLORETA文件。通过sLORE‐TA软件进行基于非参数映射法的体素随机检验(5 000次置换),对每个成分用log(fRatio)统计量评估组间在每个频段内皮层振荡的源定位差异,显著性水平P<0.05。最后生成的脑源定位图像显示出布罗德曼分区(Brodmann area,BA)和其对应的MNI坐标值(杨剑 等, 2015)。使用GraphPad Prism 8和Adobe Illustrator CS 5软件进行图片制作。
1.2.5 构建静息态EEG脑功能网络
脑功能网络的构建是利用图论的分析方法,将复杂的大脑分割为许多不同的独立脑区,以电极点作为网络节点,节点是脑区,不同电极点之间的相互关系作为边,边是脑区间的物理连接或功能相关性。计算PLI测量不同大脑区域之间的相位同步(Vinck et al., 2011)。基于相位的测量连通性的量度,不易受到容积传导效应的影响。利用希尔伯特变换(Hilbert transform),确定每个时间点的瞬时相位。计算60个电极对之间的PLI,生成一个连接矩阵,将所有PLI值进行平均,获得每个被试的连接矩阵。为了测量整体连接强度,将所有电极对的PLI取平均值,在每个时间点进行独立样本t检验,比较组间的差异PLI平均值,使用具有显著差异的时间点来识别用于进一步分析的窗口。基于网络的统计分析(network-based statis‐tics,NBS)检验两组被试间在每个频段下60个电极对之间的功能连接强度差异,显著性水平P<0.05。使用Graph‐Pad Prism 8和Adobe Illustrator CS 5软件进行图片制作。
2 结果
2.1 相对功率谱密度结果
足球运动训练后,睁眼静息态相对功率谱密度,与对照组相比,运动组在θ、α、β和γ频段上均有增高(表1)。其中,在β和γ频段上增高显著(P<0.05),在α频段上增高极显著(P<0.01)。在α频段上,与对照组相比,运动组的脑地形图表现出较大面积的活跃程度(图1)。
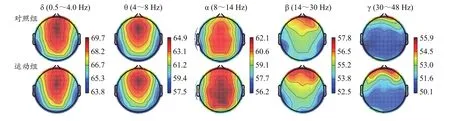
图1 静息态EEG各频段功率谱密度的脑地形图Figure 1. Brain Topographic Map of Power Spectral Density of Each Frequency Band of Resting EEG

表1 静息态EEG各频段相对功率谱密度统计结果Table 1 Statistical Results of Relative Power Spectral Density of Each Frequency Band of Resting EEG M±SD
2.2 静息态sLORETA溯源结果
全频段和各频段脑激活差异的sLORETA统计结果如图2~图7所示。
2.2.1 全频段睁眼静息态sLORETA溯源结果
在全频段中神经电活动的脑区变化不一,运动组在右侧大脑颞叶的颞中回(BA21)、MNI(65,-40,-15)和左侧大脑额叶的额下回(BA46)、MNI(-35,35,15)表现出了更高的活动性,而在左侧大脑顶叶的顶下小叶(BA40)、MNI(-40,-40,55)、颞叶的颞中回(BA37)、MNI(-40,-53,0)和枕叶的枕下回(BA19)、MNI(-40,-75,-10)表现出了较低的活动性(图2)。
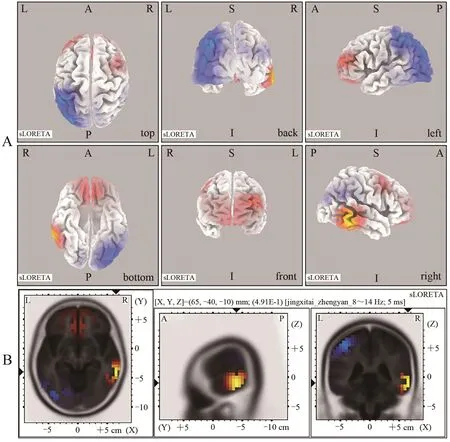
图2 运动组与对照组全频段范围内睁眼静息态下脑激活差异的sLORETA图Figure 2. sLORETA Plots of Differences in Brain Activation in Resting State with Open Eyes in the Full Frequency Band between Exercise Group and Control Group
2.2.2 各频段睁眼静息态sLORETA溯源结果
与对照组相比,运动组在δ频段上左侧大脑枕叶的梭状回(BA19)、MNI(-40,-75,-20)和顶叶的顶下小叶(BA40)、MNI(-35,-45,55)表现出活动性降低,在右侧大脑颞叶的颞中回(BA21)、MNI(65,-40,-15)和左侧大脑额叶的额中回(BA11)、MNI(-40,35,-15)表现出活动性略有升高(图3)。

图3 运动组与对照组δ频段范围内睁眼静息态脑激活差异的sLORETA图Figure 3. sLORETA Plots of Differences in Brain Activation in Resting State with Open Eyes in the δ Band between Exercise Group and Control Group
与对照组相比,运动组在θ频段上左侧大脑额叶的额中回(BA10)、MNI(-30,40,25)、右侧大脑颞叶的颞中回(BA21)、MNI(60,-40,5)表现出活动性较强,在左侧大脑枕叶的枕中回(BA19)、MNI(-30,-95,15)表现出活动性略降低(图4)。
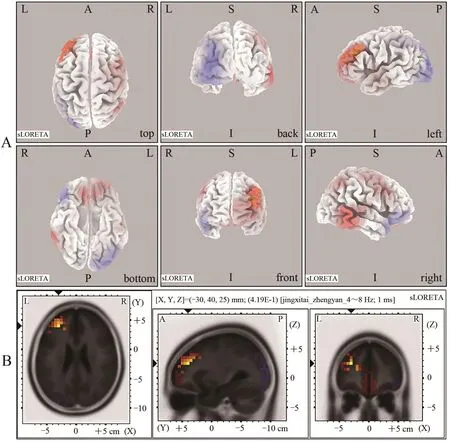
图4 运动组与对照组θ频段范围内睁眼静息态脑激活差异的sLORETA图Figure 4. sLORETA Plots of Differences in Brain Activation in Resting State with Open Eyes in the θ Band between Exercise Group and Control Group
运动组在α频段上大脑激活区域最大,与对照组相比,右侧大脑顶叶的顶下小叶(BA40)、MNI(40,-50,50)、颞叶的颞中回(BA22)、MNI(50,-40,10)、额叶的中央前回(BA4)、MNI(40,-20,50),以及左侧大脑额叶的中央前回(BA4)、MNI(-10,-35,70)表现出活动性增强(图5)。

图5 运动组与对照组α频段范围内睁眼静息态脑激活差异的sLORETA图Figure 5. sLORETA Plots of Differences in Brain Activation in Resting State with Open Eyes in the α Band between Exercise Group and Control Group
与对照组相比,运动组在β频段上左侧大脑额叶的额内侧回(BA9)、MNI(-20,40,20)、额上回(BA10),MNI(-25,45,25)、额中回(BA10)、MNI(-30,40,25)、右侧大脑额叶的额内侧回(BA9)、MNI(5,50,25)表现出活动性增强(图6)。
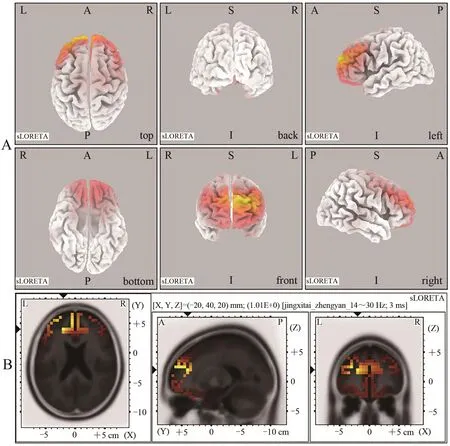
图6 运动组与对照组β频段范围内睁眼静息态脑激活差异的sLORETA图Figure 6. sLORETA Plots of Differences in Brain Activation in Resting State with Open Eyes in the β Band between Exercise Group and Control Group
与对照组相比,运动组在γ频段上左侧大脑额叶的额内侧回(BA9)、MNI(-20,40,20)、额上回(BA10)、MNI(-25,45,25)、右侧大脑额叶的额内侧回(BA9)、MNI(5,50,25)、额上回(BA10)、MNI(25,45,25)表现出活动性增强(图7)。

图7 运动组与对照组γ频段范围内睁眼静息态脑激活差异的sLORETA图Figure 7. sLORETA Plots of Differences in Brain Activation in Resting State with Open Eyes in the γ Band between Exercise Group and Control Group
2.3 静息态脑功能连接网络结果
把由每个被试得到的0、1矩阵相加,利用NBS工具箱的PLI指标筛选连接网络。在P=0.01时,阈值为2.4。阈值处理后的两组被试平均PLI连接矩阵如图8~图13所示。
本研究显示,在1 770个导联对中,与对照组相比,运动组全频段有101个连接特征呈极显著性差异(P<0.01);α频段有92个连接呈显著性差异(P<0.05);β频段有68个连接呈极显著性差异(P<0.01);δ、θ和γ频段各电极对连接无显著性差异(表2)。从图11~图12可以看出,运动组在α和β频段中的连接强度比对照组高。说明通过运动训练后,相比于对照组,运动组电极对之间的认知同步关系更紧密,大脑神经活动更复杂,活跃性更强。

表2 静息态EEG全频段和各频段脑功能连接统计结果Table 2 Statistical Results of Brain Functional Connectivity in the Whole Frequency Band and Each Frequency Band of Resting EEG
为了更直观地反映大脑网络分布情况,将连接矩阵转换为拓扑图(图8B、图9B、图10B、图11B、图12B、图13B)。两组被试的功能网络拓扑图的连线均遍布整个大脑,但有明显差异:运动组的网络结构较强,对照组相对较弱。全频段范围内静息态网络连接图显示,两组被试间差异的脑功能连接主要集中在额叶、颞叶以及枕叶等脑区(图8B)。α频段范围内,两组被试间差异的脑功能连接主要集中在额叶、颞叶以及枕叶等脑区,左半球的连接强度略高于右半球(图11B)。β频段范围内,两组被试间差异的脑功能连接主要集中在额叶和顶叶等脑区(图12B)。δ频段(图9B)、θ频段(图10B)和γ频段(图13B)的各电极对连接无显著性差异。
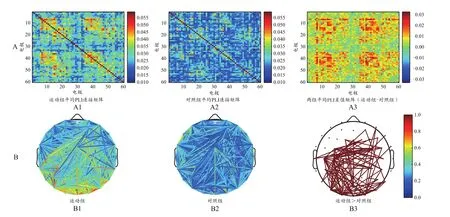
图8 全频段范围内睁眼静息态的总平均PLI连接矩阵(A)与脑功能网络分布(B)Figure 8. Total Mean PLI Connectivity Matrix (A) and Brain Functional Network Distribution (B)of Open Eyes Resting State in the Full Frequency Range
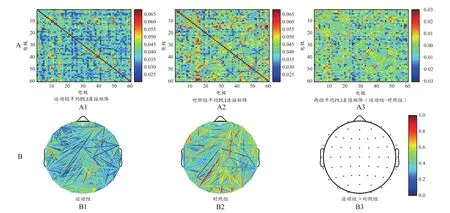
图9 δ频段范围内睁眼静息态的总平均PLI连接矩阵(A)与脑功能网络分布(B)Figure 9. Total Mean PLI Connectivity Matrix (A) and Brain Functional Network Distribution (B) of Open Eyes Resting States in the δ Range
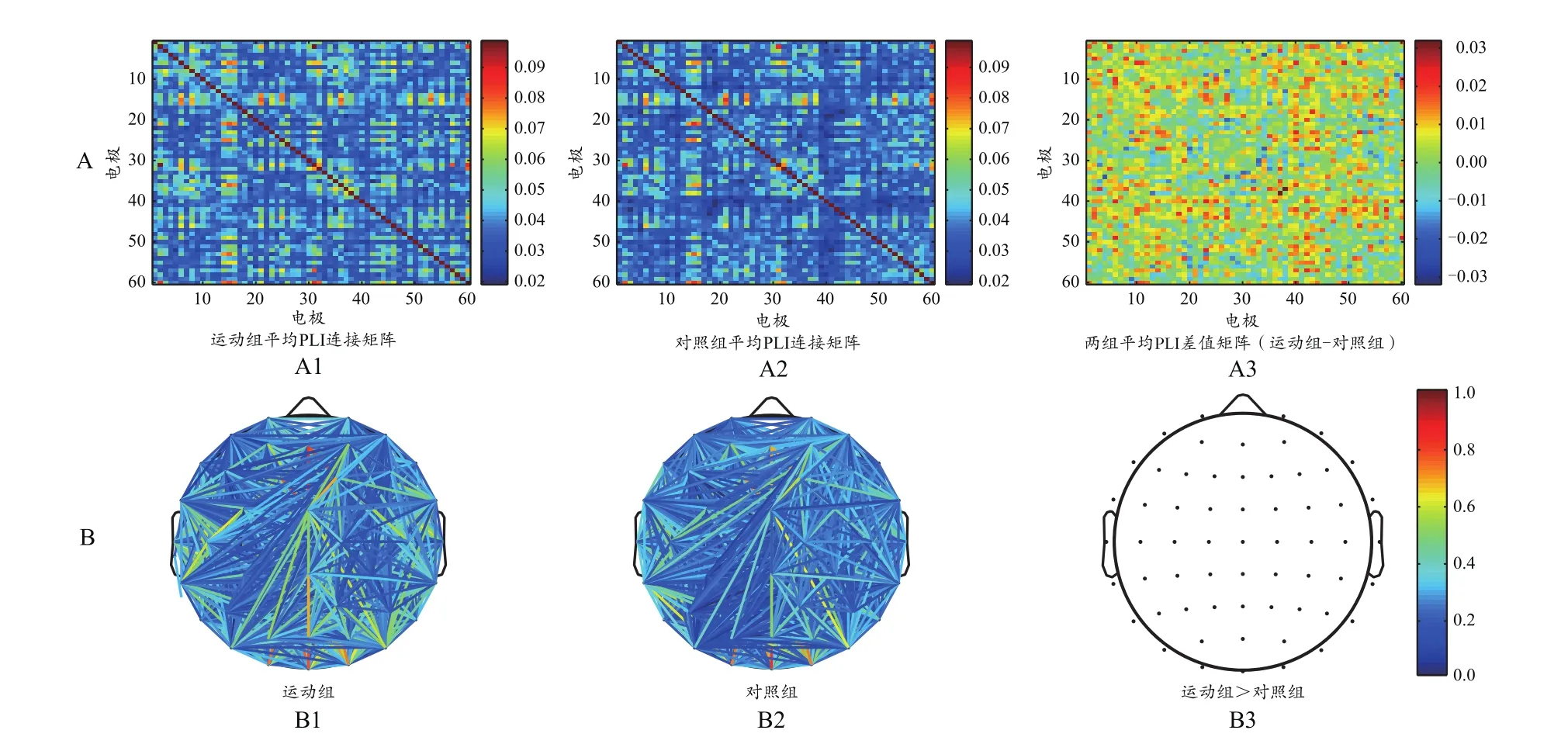
图10 θ频段范围内睁眼静息态的总平均PLI连接矩阵(A)与脑功能网络分布(B)Figure 10. Total Mean PLI Connectivity Matrix (A) and Brain Functional Network Distribution (B) of Open Eyes Resting States in the θ Range
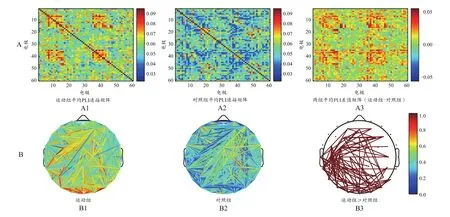
图11 α频段范围内睁眼静息态的总平均PLI连接矩阵(A)与脑功能网络分布(B)Figure 11. Total Mean PLI Connectivity Matrix (A) and Brain Functional Network Distribution (B) of Open Eyes Resting States in the α Range

图12 β频段范围内睁眼静息态的总平均PLI连接矩阵(A)与脑功能网络分布(B)Figure 12. Total Mean PLI Connectivity Matrix (A) and Brain Functional Network Distribution (B)of Open Eyes Resting States in the β Range

图13 γ频段范围内睁眼静息态的总平均PLI连接矩阵(A)与脑功能网络分布(B)Figure 13. Total Mean PLI Connectivity Matrix (A) and Brain Functional Network Distribution (B)of Open Eyes Resting States in the γ Range
3 讨论与分析
3.1 静息态功率谱结果分析
本研究表明,长期足球运动训练后,静息态下大脑α、β、γ 3个频段功率谱发生显著性增加,进而发现足球运动能提高大脑的感知行为。在闭眼静息状态下,专业运动员顶叶和枕叶区的α波更高(Babiloni et al., 2010)。α频段活动反映警觉性、记忆表现和注意力需求的皮质和丘脑-皮质网络有关(Hsieh et al., 2011;Klimesch, 2012),枕叶、顶叶、中央区以及额叶等区域表现相对较高。有研究表明,运动可能会增加α频段活性(Crabbe et al., 2004),随着任务难度的增加α功率降低(即事件相关去同步化),反映了分配给任务执行的皮层资源数量成反比关系(Bouchard, 2012)。在安静休息期间,睁眼或闭眼时θ频段(4~8 Hz)的脑电图活动减弱(Devilbiss et al., 2019)。在即时记忆任务中,δ频段功率谱的升高,以及β和γ频段功率谱的降低表明皮质活动减慢,表现出类似运动的效果(张立伟 等, 2018)。本研究发现,运动组α、β和γ频段功率谱均增加,表明高频段(α和β)更多地参与了抑制过程(Klimesch, 2012)。其中,α频段功率谱的增加,可能与下丘脑功能增强有关,人体内在注意集中时会使大脑α频段的同步化增强(吕墨竹 等,2019;孙红敏 等,2012)。β频段的出现与警觉状态相关,对周围事物很敏感,注意集中于环境,呈分散状(甘霖, 2017)。β振荡与运动和躯体感觉皮层的需求有关(Baker, 2007),也与自上而下的控制有关(Engel et al., 2010)。高β波与压力、高能耗和高唤醒有关(Wang et al., 2012),也有研究认为其与警觉或焦虑相关(Dzedzickis et al., 2020)。与乒乓球、网球等对注意要求高的运动项目相比,足球运动具有一定的身体对抗性和更多的团队配合。足球运动员在比赛时会受到外界的干扰,机体对外界刺激的处理和对多信号、多目标的处理能力效率能够有效帮助其在关键时刻做出最优选择。因此,足球运动训练可能提高了运动员的注意,而这种注意集中属于内在注意,可能是长期足球运动训练提高了运动员的内在注意集中的能力。
3.2 静息态溯源结果分析
运动技能的学习和训练与大脑结构和功能的变化有关(Dayan et al., 2011)。长期运动技能训练可导致与运动功能相关的大脑区域的结构变化,以及涉及感觉、空间和注意过程的皮质区域的结构和功能的变化(李懿婷等, 2020; Wang et al., 2016)。有研究表明,优秀耐力运动员的脑白质束增强,内侧颞叶灰质体积增大(Chang et al.,2015; Schlaffke et al., 2014)。有氧运动(如中等强度步行)可能会改变并改善大脑的结构和功能(Erickson et al.,2011)。足球运动属于长时间、高技能的运动项目,对运动员的耐力和运动技能的协调能力均有一定要求,必须同时处理许多线索作出适当反应,因此改善运动员的视觉注意处理和视觉运动协调尤其重要(Wolf et al., 2014)。
本研究溯源结果显示,运动员的激活区域分布在颞中回、额中回、额内侧回、顶下小叶。其中,额内侧回与执行机制和决策能力有关;额中回参与注意控制;颞中回分为前端和后端,前端参与视觉网络、默认网络和语言网络,后端参与语言网络和运动网络;顶叶是感觉和监控身体各部分对外界刺激反应的皮质构成,主要响应疼痛、触摸、温度、压力等感觉,该区域还与数学和逻辑相关(Hampshire et al., 2010; Kaufman et al., 2016)。本研究表明,足球运动训练促进了这些区域的激活,可能产生了结构的可塑性改变,改善了感觉、运动和认知调节。有研究发现,职业高尔夫球手运动前区和顶叶区皮质的灰质体积增加(Jäncke et al., 2009)。运动训练干预或专家的大脑皮质容量增加(Thomas et al., 2013)。内侧颞叶,特别是海马体的体积增加与更好的情景记忆表现和学习功能正相关(Ezzati et al., 2016; Pohlack et al., 2014)。增加训练量可以改善男性青少年精英足球运动员大脑皮层区域(Varley et al., 2017)。Zhu等(2021)对青年人进行为期9周的适度有氧运动干预后,发现左侧额内侧回灰质体积、左侧枕外侧皮质厚度、左侧楔前叶厚度显著增加。即使在青年人中,耐力锻炼能触发神经生物学过程,导致大脑结构变化,如前额叶区域体积的增加,进而推动认知能力的提高,有氧运动与这些区域的更大激活有关。综上所述,本研究表明,足球训练可能导致与感觉-运动相关脑区的激活,提高足球运动员的认知水平。
3.3 静息态脑功能连接网络分析
静息态功能连接指在休息期间,由于共同的激活历史,脑区之间自发的功能变化(Fox et al., 2007; Snyder et al., 2012)。在智力测试中得分较高的个人倾向于在静息状态与任务执行状态之间表现出较小的功能连接变化(Schultz et al., 2016),表明其更有效地适应任务需求。大脑网络的结构引导执行各种任务时出现的连接模式,相关研究通过静息态fMRI分析了运动训练对大脑功能结构的影响(Guerra-Carrillo et al., 2014; Kelly et al., 2014)。静息状态中测量的模式可以预测工作记忆性能(Stevens et al.,2012)和知觉任务中的刺激检测(Sadaghiani et al., 2015)。
本研究显示,脑功能连接结果与脑源定位结果一致,足球运动训练放大了特定脑区之间的功能连接,增加了感觉-运动区之间的连通性(Gross et al., 2005),特别是在β频段(Van Wijk et al., 2012)。与相关研究结果一致,运动组的α和β频段脑功能连接均有所增加(Dal Maso et al.,2018)。fMRI影像显示,在一次20 min中等强度运动后,感觉-运动网络之间的静息态连接增加(Rajab et al.,2014)。舞蹈者在控制中央前回和中央后回以及双侧壳核的运动控制和整合的皮质-基底神经节中表现出更高的连接密度(Burzynska et al., 2017; Li et al., 2015)。但也有研究得到不同结果,与干预前相比,训练后的内在功能连通性降低(Ma et al., 2011; Vahdat et al., 2011)。静息状态下体操运动员脑功能连接在小脑、额-顶区和扣带回-盖区等区域内的连通性降低,是因为这些区域的神经效率得到了提高(Wang et al., 2016)。本研究表明,足球专业运动员和非运动员之间的差异包括增强的正向连接,以及区域间的反相关性。加强的连接可能与连接区域共同激活的历史有关(Guerra-Carrillo et al., 2014),表明高水平足球运动员可能会激活这些区域的连接。有研究假设,静息状态下区域之间的反相关性反映了冲突认知需求的解决,与认知表现的改善有关(Keller et al.,2015)。一项以检测与注意策略和耐力自行车任务阶段有关的大脑皮层连接模式连通性分析结果显示,无论采用何种注意策略,所有电极对和频段(α和β)在静息状态下EEG相干值都高于循环期间EEG相干值(Di Fronso et al.,2018)。静息状态下大脑网络存在运动强度的依赖性调节,力竭运动会暂时降低大脑网络效率(Büchel et al., 2021)。因此,静息状态网络评估可以显示中枢神经系统在不同训练情况下的准备状态和效率,以及在训练监测中发挥前瞻性作用。
综上所述,本研究显示,静息状态下,各频段在颞叶、额叶和顶叶等脑区的激活较明显,同时基于PLI特征脑功能连接结果显示,在额叶、颞叶、顶叶和枕叶等脑区连接有显著性差异。这些大脑区域大多属于感觉-运动系统,说明足球运动员唤醒水平更高,更有利于视觉注意的激活。这与运动干预引起的认知增益有关,改善了与感觉-运动相关脑区之间的连接性(Baniqued et al., 2018; Gal‐len et al., 2016)。有氧运动会引起人体新陈代谢、呼吸系统和心血管系统的变化,并促进大脑结构和功能的暂时性和永久性改变(Colcombe et al., 2003)。足球运动训练诱导大脑改变的可能机制:脑部毛细血管的生长(Vivar et al., 2013);神经元之间突触连接的长度和数量的增加(Tan et al., 2016);神经营养因子(如脑源性神经营养因子和胰岛素样生长因子)水平增加引起神经元的生成,刺激神经发生和血管生成(Voss et al., 2013)。这些由于运动训练引起大脑结构的可塑性和适应性改变(Demirakca et al., 2016; Gao et al., 2019),改善了感觉、运动和认知功能的调节(梁祎明 等, 2018; Cerasa et al., 2017)。随着时间的推移,导致静息状态网络中的功能连接加强,足球运动员在视觉注意和视觉运动协调加工中整合了相关脑区,在结构和功能连接架构上均表现出了一定的可塑性变化。这些脑连接整合的可塑性改变可能影响了多模态的注意表现。网络结构和功能变化的共同作用有助于提高运动员对高注意要求的反应和执行力。
4 结论
1)足球运动员睁眼静息态大脑的α、β、γ 3个频段的功率谱显著增加。研究表明,足球运动可以提高内在注意,促进神经元之间的密切联系,提高认知行为的处理速度。
2)足球运动员的激活区域在颞中回、额中回、额内侧回、顶下小叶,与改善感知觉、情绪和精神活动,以及学习记忆有关。研究表明,足球运动促进这些区域的激活,可能产生了结构可塑性变化,改善感觉、运动和认知调节。
3)足球运动员全频段在额叶、颞叶以及枕叶等脑区存在显著性差异。其中,α频段在额叶、颞叶以及枕叶等脑区存在显著性差异,左半球的连接强度略高于右半球;β频段在额叶和顶叶等脑区存在显著性差异。研究表明,足球运动可激活与运动控制和执行功能相关的特定脑区之间的功能连接。
综上所述,足球运动员静息态额叶、颞叶和枕叶等脑区的激活较明显,脑功能连接增强。足球运动训练改善感觉、运动和认知的调节,促进这些脑区的功能连接性,激活与运动控制和执行功能相关的特定脑区之间的功能连接。提示,视觉注意、知觉预期、复杂运动执行能力和认知功能得到了提高,优化了静息态相关脑区的激活与连通性。
猜你喜欢
昆明医科大学学报(2021年2期)2021-03-29
浙江大学学报(理学版)(2021年1期)2021-01-26
中老年保健(2020年6期)2020-12-04
中国生物医学工程学报(2019年4期)2019-07-16
中学科技(2018年9期)2018-12-19
饮食科学(2017年4期)2017-05-03
健康管理(2017年3期)2017-04-20
读者(2016年18期)2016-08-23
中国医学科学院学报(2013年3期)2013-03-11
中国神经精神疾病杂志(2013年1期)2013-03-11
