
复合动植物蛋白替代鱼粉饲料中添加植酸酶对大口黑鲈生长性能、体组成、营养物质表观消化率及氮磷排放的影响
2022-02-01 10:24:00牛小天THOBELALouisTyasi陈秀梅孔祎迪邹记兴王桂芹
饲料工业 2022年24期
■牛小天 THOBELA Louis Tyasi 陈秀梅 孔祎迪 邹记兴 彭 雷 王桂芹*
(1.吉林农业大学动物科学技术学院,吉林长春 130118;2. 林波波大学,林波波波罗克瓦尼0727;3.华南农业大学海洋学院,广东广州 510642;4.辽宁省克达饲料有限公司,辽宁沈阳 110200)
大口黑鲈(Micropterus salmoides)又名加州鲈,是一种淡水凶猛肉食性鱼类,原产于美国西部的加利福尼亚州[1]。在20 世纪60 年代美国开始集约化养殖[2],80 年代被引进中国大陆开始繁殖与养殖[3],现已成为世界很多国家的饲养品种[4]。其具有肉质肥美、营养价值高、适应能力强、生长周期短、易起捕等优点,深受养殖户和消费者青睐,已成为我国主要养殖的淡水名贵肉食性鱼类之一。在集约化养殖过程中,配合饲料替代冰鲜鱼投喂对大口黑鲈养殖业的可持续发展做出了巨大的贡献,但是随着全球渔业资源逐渐的短缺以及肉食性鱼类饲料主要原料鱼粉的需求量不断增加,导致鱼粉价格居高不下,因此寻找合适的蛋白质源替代蛋白质含量高、氨基酸平衡的鱼粉已成为研究的热点。研究表明,植物蛋白源[5-6]、动物蛋白源[7-8]、昆虫蛋白源[9-10]及新型蛋白源[11]等均可被应用到大口黑鲈配合饲料中,尤其复合动植物蛋白[12]的应用不仅解决了单一蛋白质营养不均衡的缺点,也改善了鱼的生长性能及饲料利用率[13],但忽略了其在鱼类生产中对水环境的影响。众所周知,植酸是植物蛋白源中普遍存在的一种抗营养因子,它不仅能够降低饲料中磷的利用率,还会螯合某些蛋白质和其他矿物元素,这些难以降解的螯合物随着鱼体的代谢过程最终被排泄到水环境中,进而导致饲料中营养物质的流失和水体中氮、磷的负荷加重,使水体容易发生富营养化[13-14]。所以降低饲料成本,寻求适合替代鱼粉的蛋白质源,并开发出一种高效环保的配合饲料就成为了亟待解决的重要问题。
研究证实,植酸酶能够有效水解植物蛋白源中的植酸,降低其抗营养作用,改善鱼的生长和饲料利用,减少氮、磷的排放,从而改善鱼类赖以生存的水环境。饲料中添加1 500 U/kg 植酸酶可改善斑点叉尾鮰(Ictalurus punctatus)的生长性能,降低鱼体的氮、磷排泄[14]。黄颡鱼(Pelteobagrus fulvidraco)稚鱼饲料中添加植酸酶能够提高消化道的消化酶活性和饲料营养物质的表观消化率,并减少粪便中磷的排放[15]。在大豆浓缩蛋白替代鱼粉的真鲷(Pagrus major)饲料中添加植酸酶能够提高鱼体的粗蛋白含量和磷的沉积率[16]。虽然关于复合动植物蛋白源在水产动物上的研究颇多,但目前未见到大口黑鲈饲料中鱼粉被复合动植物蛋白替代的同时补充植酸酶对鱼体影响的报道。故本研究以大口黑鲈为试验对象,分析在复合动植物蛋白替代鱼粉的饲料中添加植酸酶对鱼体的生长性能、形体指标、体组成、营养物质的沉积率、表观消化率及氮磷排放的影响,为开发大口黑鲈高效环保配合饲料提供参考依据。
1 材料与方法
1.1 试验饲料
植酸酶(5 000 U/g)购于天津博菲德科技有限公司,其他原料均由辽宁省克达饲料有限公司提供。本试验采用2×3 双因素试验设计,其中以三氧化二钇(Y2O3)作为内源指示剂。以60%鱼粉组作为对照组(T0 组),在T0 组基础上利用复合动植物蛋白(虾粉、禽副产品粉、发酵豆粕、花生粕和谷朊粉)替代鱼粉,替代水平分别为30%、40%和50%,再设置3个植酸酶添加水平,分别为0.01%、0.02%和0.03%,共配置9组(T0、T1、T2、T3、T4、T5、T6、T7、T8 组和T9 组)等氮(47.50%)等能(19.33 MJ/kg)的试验饲料(见表1),并根据T0组必需氨基酸组成,添加晶体氨基酸(赖氨酸和蛋氨酸)使各组必需氨基酸趋于平衡(见表2)。
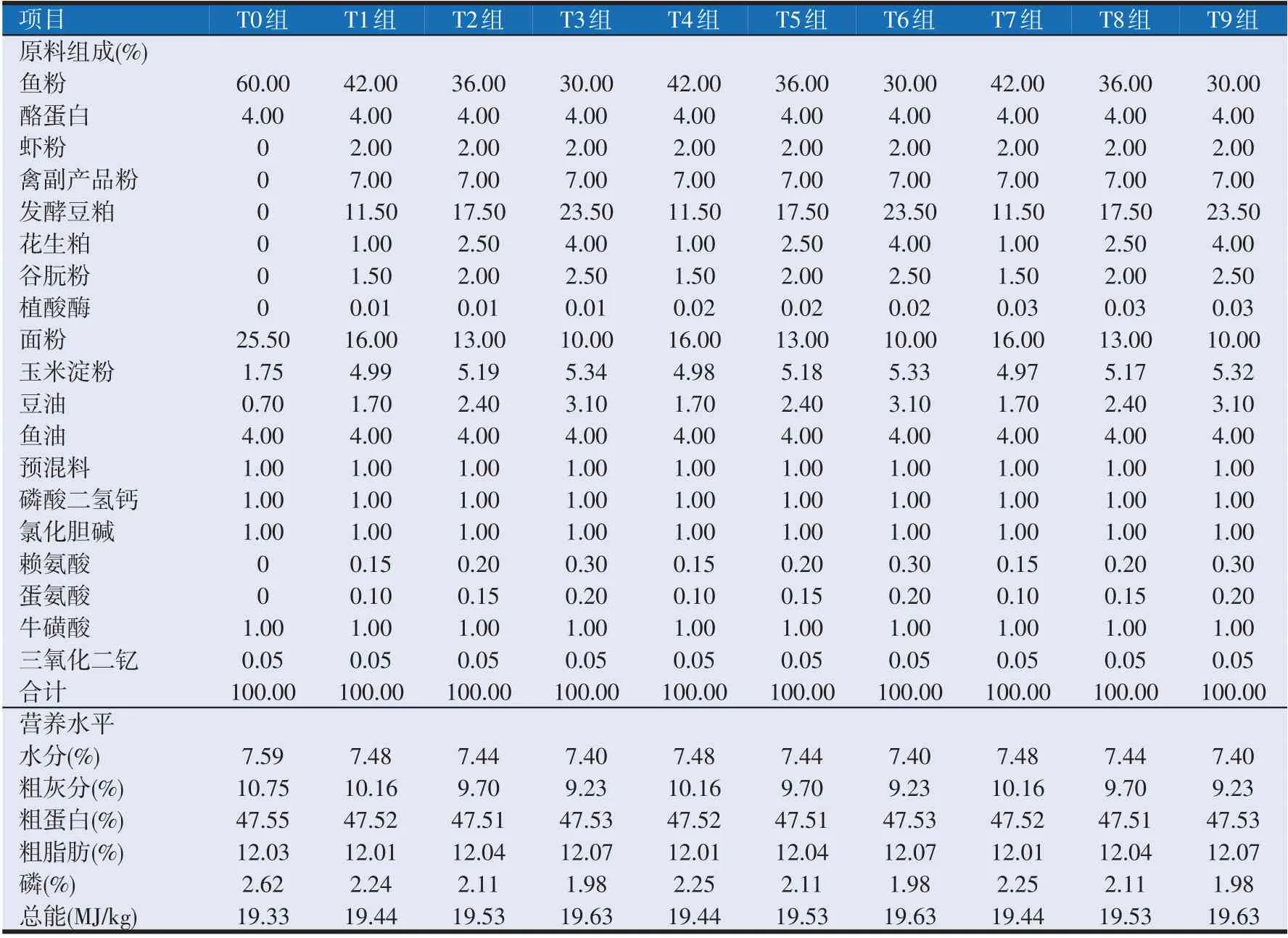
表1 试验饲料组成及营养水平(风干基础)

表2 试验饲料氨基酸组成分析(%)
将各原料粉碎后,过60目筛网,按饲料配方称重混匀后用绞肉机加工成粒径1.2 mm 的颗粒饲料,风干后保存于-20 ℃冰箱。
1.2 试验材料
大口黑鲈购自山东泰丰鸿基农业科技开发有限公司,饲养在吉林农业大学的水产养殖试验室。选取鱼体健壮、规格均一[(14.70±0.06)g]的大口黑鲈900尾,随机分到30个水族箱中,每个水族箱30尾,并将30 个水族箱随机分成10 组(每组3 重复)。暂养2 周后开始试验,暂养期投喂T0组饲料。
1.3 试验方法
养殖试验为期60 d,每天定时饱食投喂2次(8:30和17:00),投喂0.5 h后吸底清除残饵,将残饵置于烘箱中,65 ℃下烘干后称重并记录,待试验结束计算总摄食量时减掉残饵重量。水中氨氮<0.01 mg/L,溶解氧>6.5 mg/kg,pH 6.3~7.2,水温23~25 ℃,增氧泵24 h充气。每天定时排污及换水,换水量为1/3。
1.4 样品采集
养殖试验期间,每天利用虹吸法收集粪便,每次投喂饲料3 h后进行收集,然后置于烘箱中65 ℃下烘干后,-20 ℃保存待测,收集粪便直到养殖试验结束为止。
为期60 d养殖试验结束后,对试验大口黑鲈进行24 h饥饿处理,统计每个试验水族箱中活鱼尾数及饲料总投喂量。每个水族箱中随机选取10 尾鱼,用麻醉剂MS-222 处理后,纱布吸干鱼体表面水分后称重和测体长;其中5尾鱼保存在-20 ℃冰箱,用于全鱼体组成检测;剩余5 尾鱼进行解剖,记录内脏重和肝脏重,计算其形体指标。
1.5 指标计算
1.5.1 生长性能、形体指数、沉积率、表观消化率及总氮和总磷排放量指标
鱼体存活率、末均体重、增重率、特定生长率、饲料系数、蛋白质效率、脏体比、肝体比、肥满度、营养物质沉积率和表观消化率及总氮、总磷排放量等指标按照公式进行计算。
存活率(SR,%)=100×试验末鱼尾数/试验初鱼尾数
末均体重(FBM,g)=100×试验末鱼总体重/试验末鱼尾数
增重率(WGR,%)=100×(末均体重-初均体重)/初均体重
特定生长率(SGR,%/d)=100×(ln末均体重-ln初均体重)/试验天数
饲料系数(FCR)=饲料投喂量/(终末总体重-初始总体重)
蛋白质效率(PER)=(终末总体重-初始总体重)/(饲料投喂量×饲料粗蛋白含量)
脏体比(VSI,%)=100×试验鱼内脏重/试验鱼体重
肝体比(HIS,%)=100×试验鱼肝脏重/试验鱼体重
肥满度(CF,g/cm3)=100×试验鱼体重/试验鱼体长3
营养物质沉积率(NRE,%)=100×[(末均体重×终末鱼体营养物质含量)-(初均体重×初始鱼体营养物质含量)]/(饲料投喂量×饲料中营养物质含量)
表观消化率(ADC,%)=100×[1-(粪便中营养物质含量×饲料中Y2O3含量)/(饲料中营养物质含量×粪便中Y2O3含量)]
总氮排放量(TNW,g/kg)=1 000×(饲料投喂量×饲料粗蛋白含量)×(1-蛋白质沉积率/100)/[(终末总体重-初始总体重)×6.25]
总磷排放量(TPW,g/kg)=1 000×(饲料投喂量×饲料磷含量)×(1-磷沉积率/100)/[(终末总体重-初始总体重)]
1.5.2 营养成分指标
全鱼、饲料和粪便的粗脂肪、粗灰分、水分、粗蛋白和磷的测定分别根据国标法(GB/T 6433—2006、GB/T 6438—2007、GB/T 6435—2006、GB/T 6432—2018 和GB/T 6437—2018)进行检测;饲料中氨基酸含量采用华美辰L-3000 型全自动氨基酸分析仪测定;饲料、粪便中Y2O3含量采用电感耦合等离子体质谱仪(Expec 7000)测定。
1.6 数据分析
数据结果以“平均值±标准差(Mean±SD)”的形式来表示,利用SPSS 22.0 统计软件进行单因素方差和双因素方差分析,若存在显著差异性则进行Duncan’s多重比较,显著性水平为P<0.05。
2 结果与分析
2.1 复合动植物蛋白替代鱼粉饲料中添加植酸酶对大口黑鲈生长性能的影响(见表3)
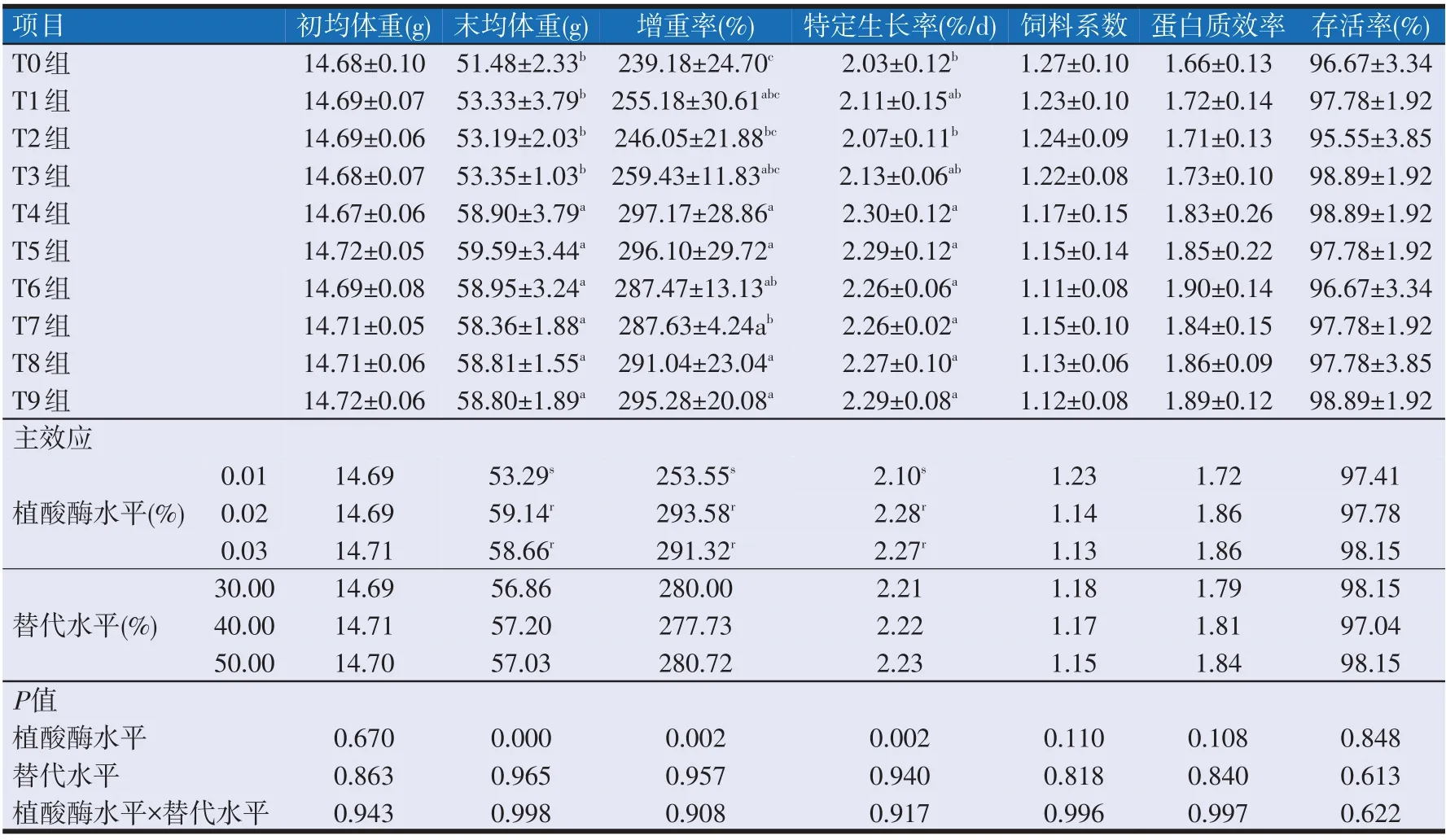
表3 复合动植物蛋白替代鱼粉饲料中添加植酸酶对大口黑鲈生长性能的影响
如表3 所示,大口黑鲈饲料中添加植酸酶水平(植酸酶水平)与复合动植物蛋白替代鱼粉水平(替代水平)交互作用对各组的大口黑鲈初均体重、末均体重、增重率、特定生长率、饲料系数、蛋白质效率和存活率均无显著影响。
植酸酶水平对各组之间的初均体重、饲料系数、蛋白质效率和存活率均无显著性影响(P>0.05);但对末均体重、增重率和特定生长率有显著性影响(P<0.05)。其中T4~T9 组的末均体重显著高于T0~T3 组(P<0.05),T0~T3 之 间 无 显 著 性 差 异(P>0.05)。T0~T3 组之间的增重率无显著性差异(P>0.05),T1、T2、T3、T6组和T7组之间的增重率无显著性差异(P>0.05),T1、T3、T4、T5、T6、T8 组和T9 组之间的增重率无显著性差异(P>0.05),但T4~T9组的增重率均显著高于T0组(P<0.05)。T4~T9组的特定生长率与T1组和T3组之间无显著性差异(P>0.05),T0~T3组之间的特定生长率无显著性差异(P>0.05),但T4~T9组的特定生长率显著高于T0组和T2组(P<0.05)。
替代水平对各组之间的初均体重、末均体重、增重率、特定生长率、饲料系数、蛋白质效率和存活率均无显著性影响。
2.2 复合动植物蛋白替代鱼粉饲料中添加植酸酶对大口黑鲈形体指标的影响(见表4)
如表4所示,植酸酶水平及植酸酶水平与替代水平的交互作用对大口黑鲈的肝体比、肥满度和脏体比均无显著性影响(P>0.05)。
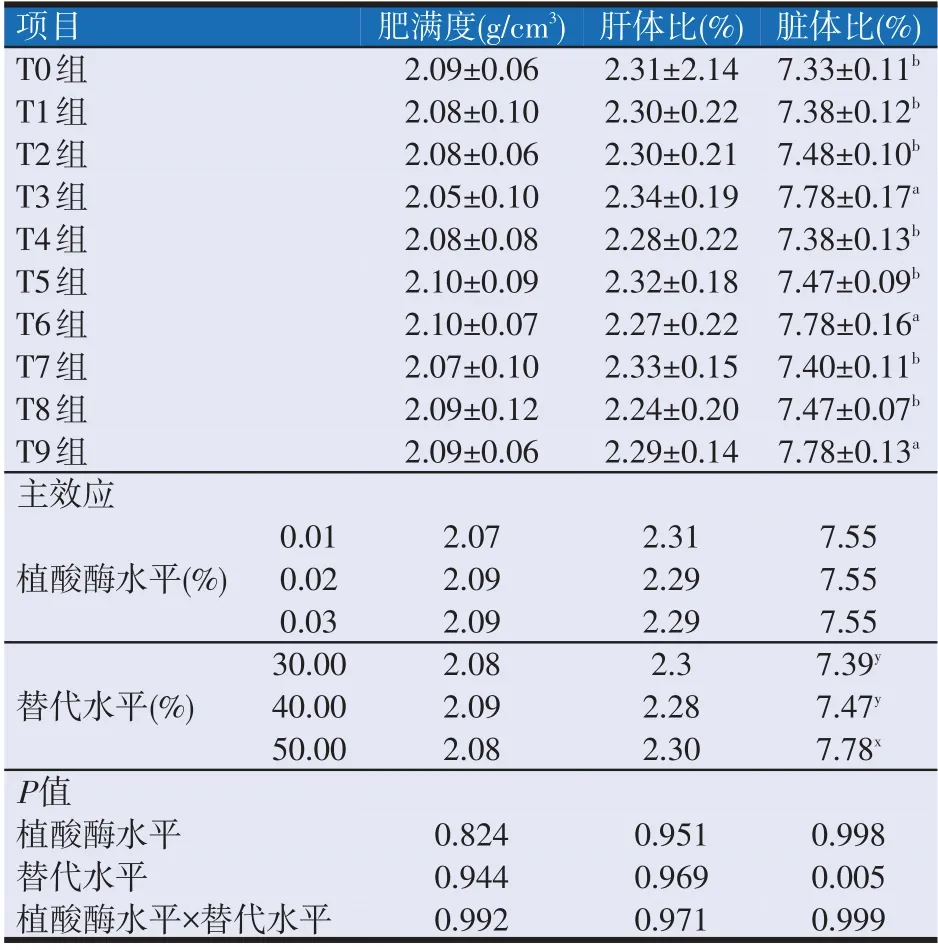
表4 复合动植物蛋白替代鱼粉饲料中添加植酸酶对大口黑鲈形体指标的影响
替代水平对各组的肝体比和肥满度均无显著性影响,但对脏体比有显著性影响。其中T3、T6组和T9组之间的脏体比无显著性差异(P>0.05),T0、T1、T2、T4、T5、T7 组和T8 组间的脏体比无显著性差异(P>0.05),但均显著低于T3、T6组和T9组(P<0.05)。
2.3 复合动植物蛋白替代鱼粉饲料中添加植酸酶对大口黑鲈全鱼体组成的影响(见表5)
如表5所示,植酸酶水平与替代水平的交互作用对各组的水分、粗灰分、粗蛋白、粗脂肪和磷含量均无显著性影响。但植酸酶水平和替代水平对各组的粗蛋白含量分别有显著性影响,其中T6和T9的粗蛋白含量与T5组和T8组无显著性差异(P>0.05),T5组和T8组的粗蛋白含量与T3无显著性差异(P>0.05),T3组与T7组之间无显著性差异(P>0.05),T7组与T2、T4组之间无显著差异(P>0.05),T1组与T0组之间无显著性差异(P>0.05),但是T2~T9组各组间的粗蛋白含量均显著高于T0组和T1组(P<0.05)。
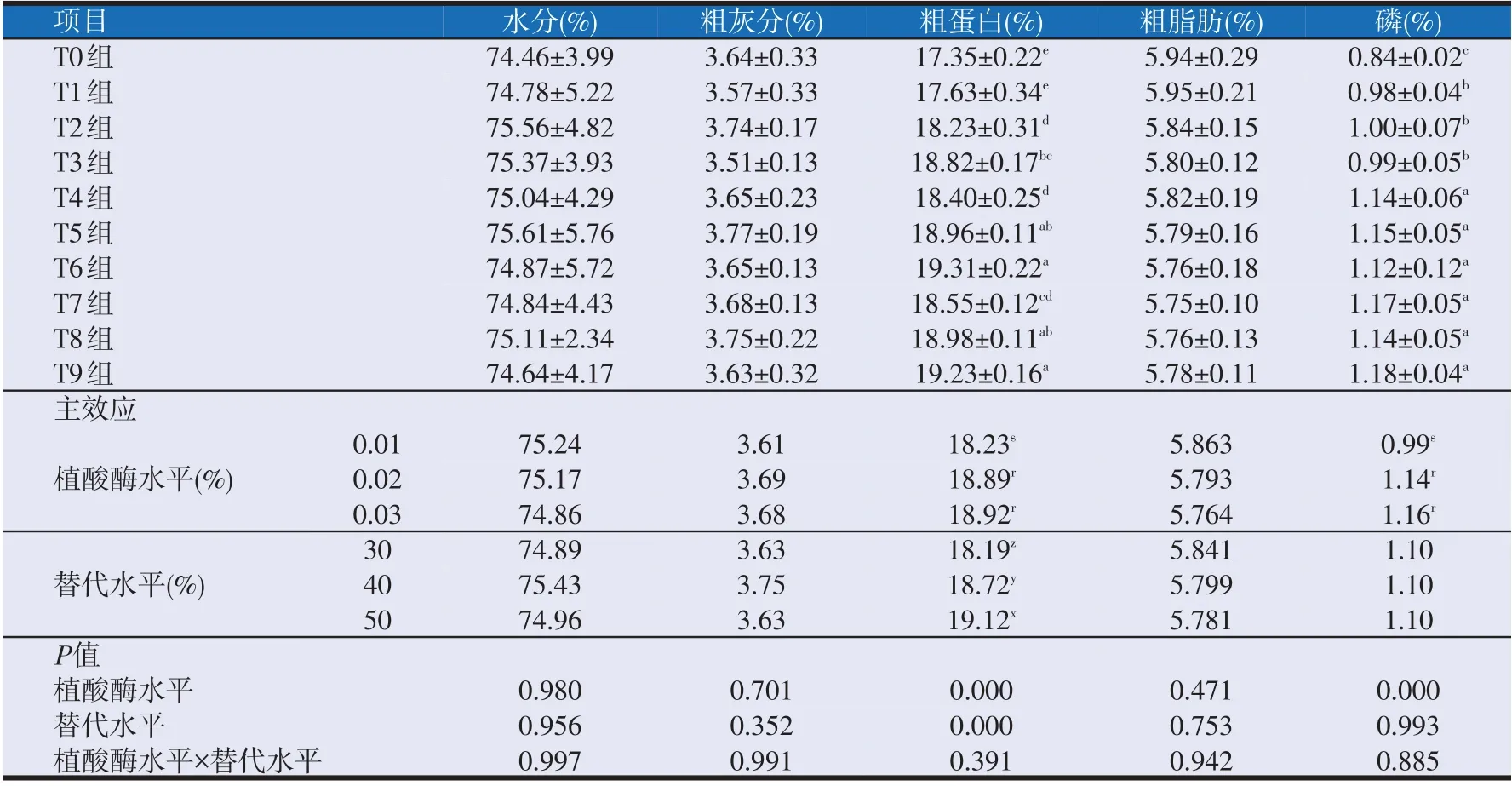
表5 复合动植物蛋白替代鱼粉饲料中添加植酸酶对大口黑鲈全鱼体组成的影响
植酸酶水平对各组的水分、粗灰分和粗脂肪均无显著性影响,但对全鱼的磷含量具有显著性影响。T4~T9 组之间磷含量无显著性差异(P>0.05),但均显著高于T0~T3 组(P<0.05);T1、T2 组和T3 组之间磷含量无显著性差异(P>0.05),但均显著高于T0组(P<0.05)。
替代水平对各组的水分、粗灰分、粗脂肪和磷含量均无显著性影响(P>0.05)。
2.4 复合动植物蛋白替代鱼粉饲料中添加植酸酶对大口黑鲈营养物质沉积率的影响(见表6)
如表6所示,替代水平及植酸酶水平与替代水平的交互作用对各组大口黑鲈粗蛋白沉积率、粗脂肪沉积率和磷沉积率均无显著性影响。

表6 复合动植物蛋白替代鱼粉饲料中添加植酸酶对大口黑鲈营养物质沉积率的影响
植酸酶水平对各组大口黑鲈的粗脂肪沉积率无显著性影响,但对粗蛋白沉积率和磷沉积率均有显著性影响。其中T1~T9组之间的粗蛋白沉积率无显著性差异(P>0.05),T0、T1、T2、T3、T4组和T7组之间的粗蛋白沉积率无显著性差异(P>0.05),而T5、T6、T8组和T9组的粗蛋白沉积率均显著高于T0组(P<0.05)。T4~T9组之间磷沉积率无显著性差异(P>0.05),但均显著高于T0~T3组(P<0.05);T1、T2组和T3组之间磷沉积率无显著性差异(P>0.05),但均显著高于T0组(P<0.05)。
2.5 复合动植物蛋白替代鱼粉饲料中添加植酸酶对大口黑鲈营养物质表观消化率的影响(见表7)
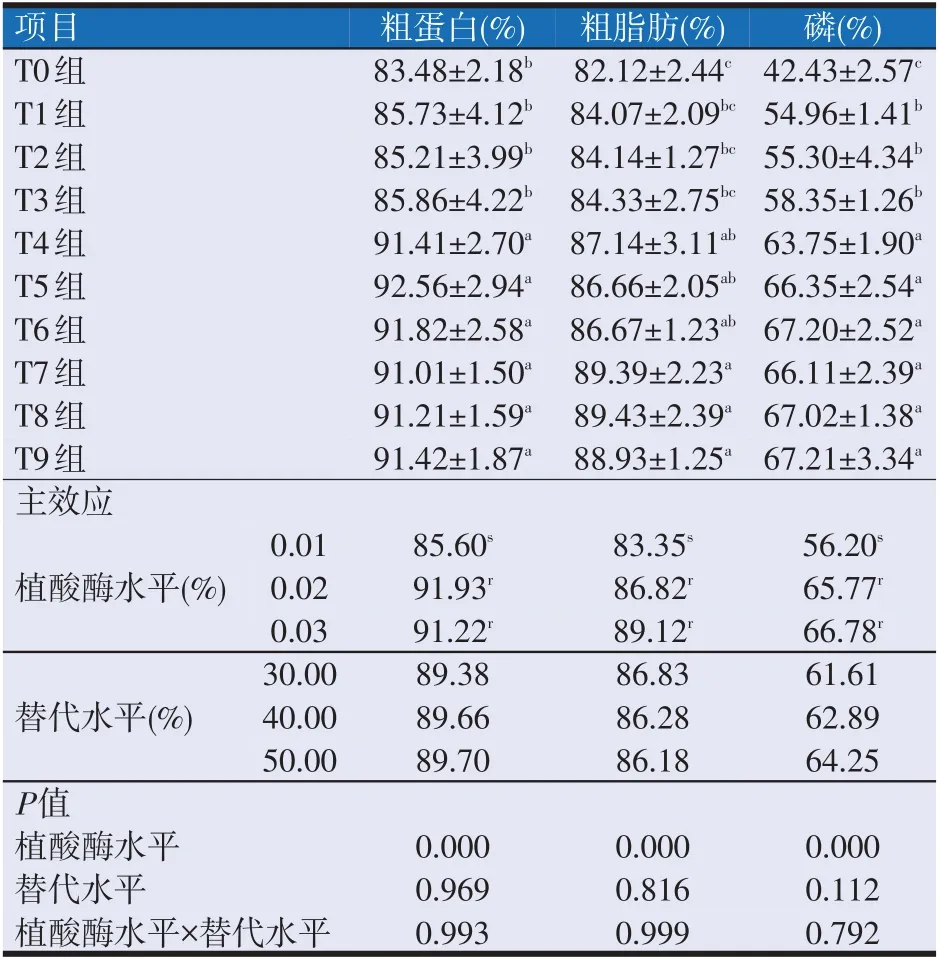
表7 复合动植物蛋白替代鱼粉饲料中添加植酸酶对大口黑鲈营养物质表观消化率的影响
如表7所示,替代水平及植酸酶水平与替代水平的交互作用对各组大口黑鲈的粗蛋白表观消化率、粗脂肪表观消化率和磷表观消化率均无显著性影响(P>0.05)。
植酸酶水平对各组大口黑鲈的粗蛋白表观消化率、粗脂肪表观消化率和磷表观消化率均有显著性影响。其中T4~T9 组之间粗蛋白表观消化率均无显著性差异(P>0.05),但均显著高于T0~T3 组(P<0.05);T0~T3组之间粗蛋白表观消化率均无显著性差异(P>0.05)。T4、T5 组和T6 组与T1、T2、T3、T7、T8 组和T9组之间粗脂肪表观消化率均无显著性差异(P>0.05),T0~T3组之间粗脂肪表观消化率均无显著性差异(P>0.05),T7、T8 组和T9 组之间粗脂肪表观消化率均无显著性差异(P>0.05);但T7、T8组和T9组的粗脂肪表观消化率均显著高于T0~T3 组(P<0.05),T4、T5 组和T6 组的粗脂肪表观消化率均显著高于T0 组(P<0.05)。T4~T9 组之间磷表观消化率均无显著性差异(P>0.05),但均显著高于T0~T3组(P<0.05);T1、T2和T3组之间磷表观消化率均无显著性差异(P>0.05),但均显著高于T0组(P<0.05)。
2.6 复合动植物蛋白替代鱼粉饲料中添加植酸酶对大口黑鲈的总氮、总磷排放量影响(见表8)

表8 复合动植物蛋白替代鱼粉饲料中添加植酸酶对大口黑鲈的总氮、总磷排放量影响
如表8所示,替代水平及植酸酶水平与替代水平的交互作用对各组大口黑鲈的总氮和总磷排放量均无显著性影响(P>0.05)。
植酸酶水平对各组大口黑鲈的总氮排放量和总磷排放量均有显著性影响。其中T1~T9 组之间的总氮排放量无显著性差异(P>0.05),T0、T1、T2、T3、T4组和T7 组之间的总氮排放量无显著性差异(P>0.05),T5、T6、T8 组和T9组之间的总氮排放量无显著性差异(P>0.05),但T5、T6、T8组和T9组的总氮排放量均显著低于T0组(P<0.05)。T4~T9组之间的总磷排放量均无显著性差异(P>0.05),但均显著低于T0~T3组(P<0.05);T1、T2组和T3组之间总磷排放量无显著性差异(P>0.05),但均显著低于T0组(P<0.05)。
3 讨论
3.1 复合动植物蛋白替代鱼粉饲料中添加植酸酶对大口黑鲈生长性能和形体指标的影响
近几年,随着水产配合饲料的开发,蛋白源替代鱼粉的研究越来越热,但研究结果不完全一致,这可能是水产动物的种类、发育阶段、生长环境、投喂方法、蛋白质源及其配比不同等因素造成的。试验在基础饲料中利用复合动植物蛋白替代鱼粉并添加植酸酶投喂大口黑鲈,发现植酸酶水平与复合动植物蛋白替代水平交互作用对大口黑鲈的生长性能和形体指标没有影响。
研究发现,利用复合动植物蛋白替代鱼粉降低了大口黑鲈肝体比,但对其肥满度、脏体比及生长性能没有影响[17]。大口黑鲈配合饲料中利用5%~30%复合蛋白源等量替代鱼粉后导致饲料系数升高,但对其他生长性能指标和形体指标没有影响[18]。分别利用动物蛋白、植物蛋白和复合动植物蛋白替代鱼粉饲喂虹鳟(Oncorhynchus mykiss)发现,复合蛋白源替代鱼粉后导致饲料转化率下降,蛋白质效率降低,而对肝体比没有影响[19]。陆生复合动植物蛋白替代鱼粉后对卵形鲳鲹(Trachinotus ovatus)的生长性能、肥满度、肝体比和脏体比均没影响[20]。本试验复合动植物蛋白替代水平对大口黑鲈的生长性能、肥满度和肝体比均没有影响,但提高了鱼体的脏体比,这与上述报道结果不完全一致,可能是复合动植物蛋白种类、配比或是替代水平不同导致的。
植酸酶能够有效水解植物蛋白源中的抗营养因子植酸,从而促进鱼类的生长和饲料利用。研究发现,鲫鱼(Carassius auratus)饲粮中添加中性植酸酶能够提高其生长性能、粗蛋白利用率和磷沉积率,降低粪磷的排放量,而对脏体比没有影响[13]。饲料中20%的红花粉替代鱼粉并添加2 000 U/kg 植酸酶能够提高虹鳟的生长性能,降低磷的排放量[21]。豆粕替代鱼粉饲料中添加500~2 000 U/kg 的植酸酶能够提高红鳍东方鲀(Takifugu rubripes)幼鱼的生长性能,对形体指标没有影响[22]。本试验结果显示,植酸酶水平改善了大口黑鲈的生长性能,但对形体指标没有影响,这与上述报道相一致,表明在复合动植物蛋白替代鱼粉的饲料中添加植酸酶能够改善大口黑鲈的生长性能。
3.2 复合动植物蛋白替代鱼粉饲料中添加植酸酶对大口黑鲈全鱼体组成的影响
试验对大口黑鲈全鱼的营养成分分析发现,植酸酶水平与复合动植物蛋白替代水平的交互作用对全鱼的水分、粗灰分、粗脂肪、粗蛋白和磷的含量均没有影响,但植酸酶水平和替代水平均对鱼体的粗蛋白质含量有显著性影响。当植酸酶水平和替代水平分别为0.02%~0.03%和40%~50%时能够改善大口黑鲈的鱼体粗蛋白含量;同时饲料中植酸酶水平对全鱼的水分、粗灰分和粗脂肪含量均没有明显的改善,但当植酸酶添加水平在0.02%~0.03%时,全鱼的磷含量显著改善。Ma 等[23]利用家禽副产品粉和豆粕替代鱼粉的研究中发现,复合动植物蛋白替代水平对卵形鲳鲹全鱼的水分、粗灰分、粗蛋白和粗脂肪含量均没有影响,却降低了磷的含量。有研究发现复合动植物蛋白替代鱼粉投喂卵形鲳鲹,能够降低全鱼粗蛋白和磷的含量[20]。谢帝芝等[17]利用复合动植物蛋白替代鱼粉的研究发现大口黑鲈的全鱼粗蛋白含量升高,但对磷、粗脂肪、水分和粗灰分没有影响。南亚野鲮(Labeo rohita)饲料中添加植酸酶和柠檬酸能够提高全鱼粗蛋白的含量[24]。豆浆浓缩蛋白替代鱼粉的饲料中添加不同水平植酸酶研究发现,植酸酶水平对真鲷的鱼体组成没有影响[16]。饲料中添加植酸酶能够提高草鱼(Ctenopharyngodon idella)鱼体的粗蛋白和磷的含量[25]。上述报道与本试验结果不完全一致,可能是鱼的种类、食性及消化系统的差异造成的,但是本试验结果表明复合动植物蛋白饲料中添加植酸酶能够改善大口黑鲈的体组成。
3.3 复合动植物蛋白替代鱼粉饲料中添加植酸酶对大口黑鲈营养物质沉积率的影响
营养物质的沉积率能够反映出鱼体的营养水平及健康状态[26]。本试验结果显示,复合动植物蛋白替代水平及植酸酶水平与替代水平的交互作用对各组大口黑鲈粗蛋白沉积率、粗脂肪沉积率和磷沉积率均没有影响。有研究发现复合动植物蛋白饲料对大口黑鲈的氮和磷沉积率均没有影响[17]。利用家禽副产品粉和豆粕替代鱼粉发现随着替代水平的升高大口黑鲈粗蛋白沉积率降低,磷沉积率升高,脂肪沉积率没有影响[27]。牙鲆(Paralichthys olivaceus)饲料中复合动植物蛋白替代0~32%鱼粉时不影响粗蛋白沉积率和粗脂肪沉积率,当替代水平达到48%时,降低了粗蛋白沉积率[28]。鱼粉分别被两种复合动植物蛋白替代,发现低替代水平对营养物质的沉积率没有影响,而高替代水平能够提高卵形鲳鲹内氮的沉积率,而磷的沉积率没有影响[20]。这与本试验替代水平的研究结果不完全一致,分析可能是复合动植物蛋白替代鱼粉的水平不同及复合蛋白种类不同导致的结果。
本试验植酸酶水平对各组大口黑鲈的粗脂肪沉积率没有影响,但能够提高粗蛋白沉积率和磷沉积率,即当复合动植物蛋白饲料中添加植酸酶0.02%~0.03%时能够提高大口黑鲈的粗蛋白沉积率和磷沉积率。有研究发现豆浆浓缩蛋白替代鱼粉的饲料中添加0.04%植酸酶能够提高真鲷的粗蛋白沉积率、粗脂肪沉积率和磷沉积率[16]。Norag 等[29]对尼罗罗非鱼(Oreochromis niloticus)研究发现,低磷饲料中添加植酸酶能够提高粗蛋白沉积率、粗脂肪沉积率和磷沉积率。这与本试验的结果相类似,表明大口黑鲈的复合动植物蛋白饲料中添加植酸酶能够提高其营养物质的沉积率。
3.4 复合动植物蛋白替代鱼粉饲料中添加植酸酶对大口黑鲈营养物质表观消化率的影响
营养物质表观消化率能够反映出鱼体对营养物质的消化程度,从而间接表明鱼类消化酶活性的强弱[30]。有研究发现复合动植物蛋白替代75%的鱼粉对虹鳟的粗蛋白表观消化率没有影响[19]。刘运正等[31]利用酶解动物软骨蛋白粉和植物蛋白替代鱼粉的研究发现,随着替代水平的升高大菱鲆(Scophthalmus maximus)的粗蛋白表观消化率降低。而本试验结果显示,复合动植物蛋白替代水平对大口黑鲈的营养物质表观消化率没有影响,这与上述报道中不完全一致,可能是鱼的种类不同导致的结果,且本试验的植酸酶水平和蛋白替代水平对大口黑鲈的营养物质表观消化率没有交互作用。
在真鲷的饲料中添加植酸酶发现,随着添加水平的升高粗蛋白表观消化率没有显著变化[16]。在鲫鱼的研究中发现,在含有0.8%磷酸二氢钙的饲料中添加植酸酶能够提高粗蛋白表观消化率、粗脂肪表观消化率和磷表观消化率[13]。南亚野鲮饲料中添加植酸酶能够提高粗蛋白表观消化率、粗脂肪表观消化率和磷表观消化率[24]。这与本试验的结果相一致,这表明在大口黑鲈复合动植物蛋白替代鱼粉饲料中添加植酸酶能够改善营养物质的表观消化率。
3.5 复合动植物蛋白替代鱼粉饲料中添加植酸酶对大口黑鲈鱼体总氮、总磷排放量的影响
近年来,随着大口黑鲈集约化养殖的发展,鱼体健康不仅受到疾病的威胁,还面临着水环境逐渐恶化的影响,其中氮磷是导致水质污染的重要因素之一[32]。本试验结果显示复合动植物蛋白替代水平及植酸酶水平与替代水平的交互作用对大口黑鲈的总氮排放量和总磷排放量均没有影响。大口黑鲈的配合饲料中鱼粉被复合动植物蛋白替代后,对总氮排放量没有影响,但是降低了总磷的排放量[17]。随着卵形鲳鲹饲料中复合动植物蛋白替代鱼粉水平的升高,能够提高其总氮排放量,但是对总磷排放量没有影响[23]。利用复合动植物蛋白替代鱼粉后,能够降低虹鳟的总氮排放量和总磷排放量[20]。上述报道与本试验结果不完全一致,这可能是动植物蛋白源种类不同、比例不同或鱼的种类不同导致的。
研究发现,在鲫鱼的饲料中添加植酸酶能够降低其总磷排放量[13]。斑点叉尾鮰饲料中添加植酸酶能够降低总氮排放量和总磷排放量[14,33]。本试验结果与上述报道一致,表明在复合动植物蛋白替代鱼粉的饲料中添加植酸酶能够降低大口黑鲈的总氮排放量和总磷排放量,减轻水环境的富营养化及污染。
4 结论
本试验条件下,配合饲料中复合动植物蛋白替代40%的鱼粉并添加0.02%~0.03%的植酸酶能够改善大口黑鲈的生长性能、形体指标、体组成、营养物质沉积率、表观消化率和氮磷排放量,对蛋白质的节约与水环境的保护起到重要作用。
猜你喜欢
垂钓(2023年11期)2024-01-21 16:07:04
当代水产(2022年8期)2022-09-20 06:46:06
当代水产(2022年6期)2022-06-29 01:13:12
国外畜牧学(猪与禽)(2021年6期)2022-01-11 05:59:40
基层中医药(2021年6期)2021-11-02 05:46:10
国外畜牧学(猪与禽)(2021年4期)2021-08-31 01:53:40
当代水产(2019年6期)2019-07-25 07:52:08
当代水产(2019年1期)2019-05-16 02:41:50
创新作文(小学版)(2018年10期)2018-08-20 03:11:38
中国畜禽种业(2018年1期)2018-01-18 23:05:48
