湿地植物香蒲根系抗氧化酶活性和根系分泌物对阿特拉津胁迫的响应
2022-01-24 07:53:38武淑文侯磊刘云根范黎明叶敏
农业环境科学学报 2021年12期
武淑文,侯磊,刘云根,范黎明,叶敏*
(1.云南农业大学植物保护学院,昆明 650201;2.西南林业大学生态与环境学院,昆明 650224)
阿特拉津是世界各地农业生产中使用最广的除草剂,其在土壤上的吸附亲和力较低,易随农田灌溉和降雨迁移进入水体环境,而适中的水溶性、较长的残留期使其在地表水和地下水中检出频率高[1]。阿特拉津通过在水生植物体内累积而干扰光合作用,阻碍碳水化合物合成等代谢过程,造成水生植物种群的退化和消亡[2],严重威胁水生态系统的生物多样性[3]。因此,阿特拉津的生态毒理学效应和污染植物修复受到广泛关注。目前,对阿特拉津植物毒性效应及作用机理的研究发现,阿特拉津诱导植物体内活性氧自由基(ROS)的大量产生,引起水葱[4]、千屈菜[5]、菖蒲[6]、美人蕉[7]、芦苇[8]等湿地挺水植物叶片抗氧化防御系统[包括超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、还原型谷胱甘肽(GSH)等]的应激响应,且随着胁迫强度的不断提高,ROS产生量超过抗氧化系统防御能力,植物细胞膜脂过氧化反应加剧,细胞膜结构和功能受损,最终抑制植物的生长。与地上茎叶相比,植物根系因直接与除草剂接触[9],而最先受到胁迫,产生的生理生化应激反应更剧烈。马兵兵等[10]关于阿特拉津长期胁迫对狼尾草氧化应激指标影响的研究发现,狼尾草根系氧化应激系统对阿特拉津胁迫的响应较叶片更为敏感。
根系分泌物是植物根系释放到周围环境的有机物质的总称,在植物面临胁迫时,根系通过生物反馈调节机制改变分泌物的组成和含量,改善根际区域微生态环境,缓解养分有效性低、重金属或有机污染物胁迫等逆境,保证植物的存活[11]。LI 等[12]的研究发现,在杀虫剂呋虫胺胁迫下,大白菜根系呋虫胺含量升高并产生氧化应激响应,引起体内丙二醛(MDA)含量的增加,根系分泌的低分子量有机代谢物发生明显改变,植物对胁迫环境的耐受性增强。潘声旺等[13]的研究发现,在芘胁迫下羊茅属植物根系分泌物中低分子量有机物释放量增多,成分变复杂,从而有利于促进根际多环芳烃的降解,提高植物的适应性。此外,面对同样的胁迫环境,不同植物根系分泌物的响应差异较大,如FU 等[14]发现两种基因型的水稻在镉胁迫条件下,植物根系分泌的低分子量有机物差异明显,有机酸数量增加,水稻中镉含量与有机酸含量呈显著正相关,氨基酸的组成和数量均发生了明显变化。目前,关于植物对农药胁迫生理响应的研究主要集中于陆生植物根系分泌物对根际生态环境的影响,及分泌物中化感物质的分离鉴定和产生机理,而以水生植物为对象,探讨水生植物在阿特拉津胁迫下根系抗氧化系统及根系分泌物的研究鲜见报道。此外,植物在修复除草剂污染时均需与污染物持续接触,因此开展阿特拉津长期胁迫下水生植物根系的生理响应研究有助于更好地理解植物对污染的修复机理。
香蒲(Typha angustifoliaL.)是一种常见的多年生草本植物,具有易繁殖、生长旺盛、根系结构发达、根际生物群落丰富、可以在不同环境条件下生长等优势,被广泛应用于污水及农业面源污染处理的人工湿地[15]。张翠萍等[16]利用生长香蒲和芦苇的人工湿地处理六氯苯(HCB)发现,由于香蒲根系比芦苇根系更发达密集,根系环境稳定[17],而更能适应外界环境变化,其根系分泌物含量和HCB 降解率均高于芦苇组和对照组。CEJUDO 等[18]研究了香蒲、泽泻茨菇、稗等植物根系对阿特拉津的积累能力,结果表明由于香蒲根系发达且根皮层孔隙多、易吸收污染物而在3 种植物中积累的阿特拉津量最多,有效降低了水体阿特拉津浓度。MOORE 等[19]利用香蒲处理农业暴雨径流中阿特拉津,结果表明阿特拉津浓度降低约54%,而未种香蒲的对照仅降低17%~28%。本研究以香蒲为受试对象,通过水培实验,探究在阿特拉津胁迫条件下根系对阿特拉津积累、抗氧化酶活性以及分泌物组成与含量的变化特征,以期为深入研究阿特拉津胁迫条件下植物修复的根际调控过程与作用机制提供理论依据。
1 材料与方法
1.1 实验材料
阿特拉津(纯度≥97%)购于上海源叶生物科技有限公司。实验所用香蒲购于昆明泛亚花卉公司,为一年生幼苗。清洗去除植物根系淤泥后,自来水中缓苗24 h,再移入5% Hoagland 营养液中预培养驯化适应14 d。待长出新根后,挑选生长正常、长势一致的香蒲幼苗[平均鲜质量(21.02±4.7)g]移入塑料圆桶(直径18 cm、高19 cm)上的定植篮内,植物根部穿过篮底浸入营养液中,篮内用砾石固定植物,定植篮底部高于桶内液面。用黑色塑料袋包裹做避光处理(防止阿特拉津光解),每桶放置3株幼苗。
1.2 实验设计
配制1 g·L−1阿特拉津母液,再用5%Hoagland 营养液稀释至实验浓度。阿特拉津浓度设置参考文献[5−6]和预实验结果,依次为0(对照)、0.2、0.4、2 mg·L−1,每组3 个重复。胁迫实验在温室进行,光照/黑暗时间为14 h/10 h,对应温度为30 ℃/20 ℃,白天光强约为180µmol·m−2·s−1。每日补充蒸馏水保持溶液体积不变,每周更换含有相应阿特拉津浓度的营养液,保证实验周期内阿特拉津胁迫稳定。胁迫45 d 后收获植物测定生物量,将采集的植物根系分成两部分,一部分测定植物根系分泌物,另一部分测定植物根系抗氧化酶指标和阿特拉津含量。
1.3 测定指标和方法
香蒲生物量的测定:植物样品采集后放入电热恒温鼓风干燥箱内,105 ℃杀青15 min 后,70~80 ℃烘至恒质量,记录植物干质量。
根系分泌物的收集与分离:参考韦雪晶等[20]报道的方法并加以改进。将香蒲根系用去离子水冲洗干净,浸入500 mL 0.05 mmol·L−1氯化钙溶液12 h,期间用黑色塑料袋盖住整个根系避光处理。含有根系分泌物的水溶液经0.45 µm 微孔滤膜过滤后加入二氯甲烷连续萃取3 次,合并萃取液有机相,真空旋转蒸发仪浓缩近干,加入2 mL 色谱纯二氯甲烷润洗有机相并经0.45µm微孔滤膜过滤后待测。
根系分泌物的GC−MS 分析:采用安捷伦7890A−5975C 型气相色谱/质谱联用法(GC−MS)测定根系分泌物。色谱条件:毛细管柱为HP−5ms 柱(30 m×250µm×0.25µm);载气为He,流量1 mL·min−1,进样量1µL;不分流进样;柱程序升温,起始温度50 ℃,保持2 min,以每分钟3 ℃升温至180 ℃,以每分钟9 ℃升温至250 ℃,保持20 min。质谱条件:采用电子轰击源(EI),轰击电压70 eV,离子源温度230 ℃;扫描方式为全扫描模式(SCAN),扫描速度0.2 s,扫描范围M/Z30−600AMU。
抗氧化系统指标测定:过氧化氢酶(CAT)、超氧化物歧化酶(SOD)、谷胱甘肽(GSH)、丙二醛(MDA)等均采用南京建成生物工程研究所开发的试剂盒进行测定。
根系阿特拉津提取与测定:提取方法参照BEN⁃NETT 等[21]报道的方法并加以改进。准确称取香蒲根1 g,用研钵研磨成糊状,加入1 mL蒸馏水润湿样品后加入10 mL 乙酸乙酯并转移至50 mL 离心管于摇床振荡20 min,收集有机相提取液。样品重复上述提取过程2 次,合并的提取液经装有5 g 无水硫酸钠的漏斗脱水干燥后转移至旋转蒸发仪,在小于40 ℃条件下减压浓缩至近干,用甲醇溶液(V甲醇∶V水=1∶1)定容至1 mL 后于4 ℃保存备用。阿特拉津的测定根据国家环境保护标准《水质 阿特拉津的测定高效液相色谱法》(HJ 587—2010)并加以改进。采用安捷伦(Ag⁃ilent 1260)高效液相色谱仪,色谱条件:C18 反相柱(100 mm×4.6 mm),填空颗粒粒径2.6 µm;V甲醇∶V水=50∶50 作为流动相,流速为0.5 mL·min−1,检测波长223 nm;柱温40 ℃,进样量为5µL;采用梯度洗脱,流动相0~10 min,55%~65%,10~11 min,60%~95%;洗柱6 min(甲醇95%,水5%)。
1.4 数据分析与处理
香蒲根系分泌物鉴定采用总离子流图谱与NIST08 质谱数据库的标准图谱核对分析,通过计算机检索进行未知物测定,确定各组分物质名称,选取匹配度在90%以上的检出物质进行分析,根据色谱图中各检出成分出峰面积占总峰面积的比例计算相对含量。
采用SPSS 20.0 软件对阿特拉津胁迫下香蒲生物量、根系阿特拉津含量和抗氧化系统指标数据进行单因素方差分析(One−way ANOVA),采用Duncan 比较不同处理间的差异性(P<0.05)。利用Canoco 5 进行冗余分析(Redundancy analysis,RDA),评价不同因子间的相互关系。采用Origin 2018绘制扫描图谱。
2 结果与讨论
2.1 阿特拉津胁迫对香蒲生物量、根系积累和抗氧化系统的影响
阿特拉津胁迫对香蒲生物量和根系阿特拉津含量的影响如图1 所示。与对照相比,阿特拉津浓度≥0.2 mg·L−1显著降低了香蒲生物量(P<0.05)。对照处理香蒲根系未检测到阿特拉津累积,阿特拉津在植物根系中的积累量随其处理浓度的增加而显著增加(P<0.05),在高浓度(2.0 mg·L−1)处理下积累量达最大值0.365 mg·kg−1(以鲜质量计,下同),是最低处理浓度(0.2 mg·L−1)时的2.52 倍。阿特拉津胁迫可抑制香蒲生长,香蒲根系对阿特拉津的积累作用随胁迫浓度的升高而加强。
阿特拉津胁迫对香蒲根系抗氧化酶指标的影响如图2 所示。随着阿特拉津浓度升高,SOD 活性受到显著抑制(P<0.05),2.0 mg·L−1处理时抑制作用最强,SOD 活性较对照下降89.56%。CAT 活性随阿特拉津浓度的升高呈现先升后降的趋势,在0.4 mg·L−1处理时活性最高,较对照提高123.7%(P<0.05)。GSH 活性与对照相比差异不显著(P>0.05)。低浓度阿特拉津处理(0.2 mg·L−1)对根系MDA 含量影响不显著(P>0.05),而在高浓度(2.0 mg·L−1)时含量达到最大值且显著高于其他处理和对照(P<0.05)。
研究发现,阿特拉津胁迫导致植物生物量下降和生长发育迟缓,如水葱[4]、千屈菜[5]和芦苇[8]等水生植物在阿特拉津胁迫下生物量显著降低。本研究发现,随着阿特拉津浓度增加,根系阿特拉津累积量明显升高,而香蒲生物量显著降低。
阿特拉津胁迫诱发植物体内生成大量的活性氧自由基(ROS),使植物产生氧化应激响应从而激活内源酶促防御系统进行自我保护[22]。抗氧化内源保护系统由抗氧化酶类和非酶抗氧化剂组成,其中SOD、CAT 是酶类抗氧化保护系统的两类主要活性酶,SOD催化歧化超氧阴离子自由基()生成H2O2和O2[23],而CAT 能够清除高含量的H2O2分子[24]。GSH 为典型的非酶抗氧化剂,其可结合并清除膜脂过氧化的中间产物(过氧化物),避免其对细胞的破坏[25]。一般认为,低浓度阿特拉津能够提高植物抗氧化酶活性以清除胁迫产生的ROS,而高浓度则导致其活性下降,可能引起植物的损伤[22]。也有研究发现,植物面对逆境胁迫时,抗氧化酶会相互调控作出不同的响应,其活性呈现不同的变化趋势[8]。本研究中,随着阿特拉津浓度的增加,根系SOD 活性较对照显著下降,这是由于根系与阿特拉津溶液直接接触并吸收积累污染物,SOD活性对阿特拉津处理尤为敏感,低浓度的阿特拉津积累量可能对其已造成了氧化胁迫,导致SOD 活性降低,削弱其保护作用,继而影响其对植物体内活性氧的正常清除[26]。阿特拉津胁迫下根系CAT活性表现为先增强后减弱的趋势,在0.4 mg·L−1处理下CAT活性最大,且显著高于对照,即在轻度胁迫下(≤0.4 mg·L−1),植物通过激活CAT 酶活性来清除有害物质,但随着胁迫强度的增大,活性氧产生量超过CAT 清除能力,植物细胞发生膜脂过氧化反应而受损,造成CAT活性下降。研究发现,阿特拉津胁迫下皇竹草叶片SOD 与CAT 活性呈现出与本研究类似的变化规律[26]。根系GSH 活性随着阿特拉津胁迫程度增加呈先升高再下降的趋势,但各处理间无显著差异,分析认为香蒲根系GSH 活性的升高是植物主动应对一定水平(≤0.2 mg·L−1)阿特拉津胁迫的反应。对比发现,香蒲根系SOD、CAT和GSH活性在阿特拉津胁迫下表现出的变化趋势各不相同,反映了植物根系是通过抗氧化活性酶间的相互协调来防御阿特拉津胁迫的[26]。在浓度高于0.4 mg·L−1的阿特拉津长期胁迫下,香蒲根系抗氧化系统防御能力被明显削弱,植物生长受到显著抑制。
MDA 是细胞膜脂过氧化的重要产物,其含量的高低反映了植物细胞膜受伤害的程度[6]。研究发现,黑麦草体内高阿特拉津含量能够引起细胞膜质过氧化程度的加重,使MDA 含量升高[27]。本研究中,香蒲根系阿特拉津和MDA 含量在浓度≥0.4 mg·L−1的阿特拉津胁迫下显著升高,表明该胁迫水平诱发的活性氧积累超过了抗氧化酶的防御能力,造成SOD、CAT 和GSH活性均降低,细胞膜脂过氧化反应加剧而使根系受损,引起MDA 含量的提高;而阿特拉津浓度为0.2 mg·L−1时,根系阿特拉津积累量低,未引起MDA 含量的显著增加,这与阿特拉津胁迫下狼尾草根系MDA含量的变化趋势相一致[10],说明该水平胁迫并未对香蒲根系造成明显的氧化损伤,这是SOD、CAT 和GSH等抗氧化防御系统在应对阿特拉津胁迫过程中综合调控的结果。
2.2 阿特拉津胁迫对香蒲根系分泌物的影响
香蒲在0、0.2、0.4 mg·L−1和2.0 mg·L−1阿特拉津胁迫下根系分泌物全扫描GC−MS 图谱如图3 所示。由图可知,香蒲根系分泌物的特征峰明显且密集分布,基线漂移较小,所测结果可信。不同浓度阿特拉津胁迫下的扫描图谱特征峰数量及分布面积差异较明显。
阿特拉津胁迫对根系分泌物类别数和化合物种类数的影响如表1 所示。在阿特拉津胁迫下香蒲根系分泌物主要包括烷烃、烯烃、胺、酚、醇、酯、酮和有机酸类化合物,其中阿特拉津处理对分泌物类别总数无明显影响,而化合物种类数均高于对照,2.0 mg·L−1处理较对照增加32.14%,即阿特拉津浓度的增大促进了根系化合物的分泌。此外,不同浓度阿特拉津胁迫下根系分泌的化合物均以结构较简单的烷烃类居多,结构相对复杂的胺、酚、醇、酯和酮类等化合物较少。
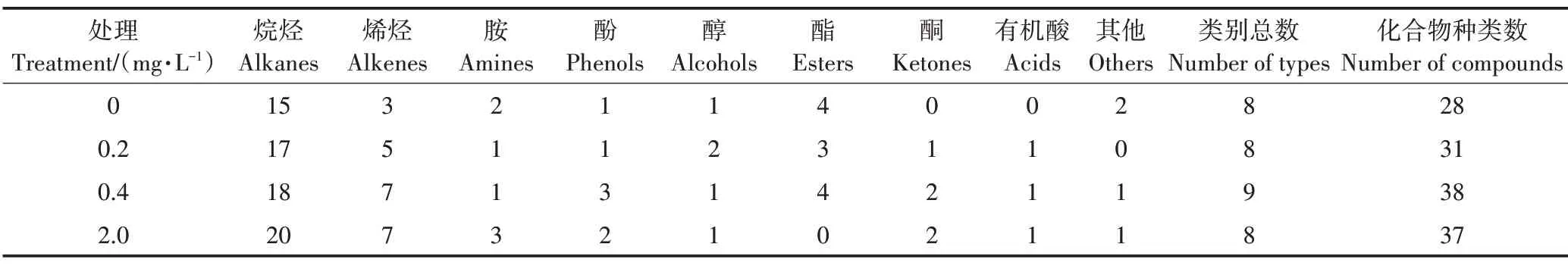
表1 阿特拉津胁迫下香蒲根系分泌物化合物种类数的变化Table 1 Number of compounds in root exudates of cattail under atrazine stress
阿特拉津胁迫对根系分泌物相对含量的影响如表2 所示。由表可知,根系分泌物中烷烃类相对含量最大,胺类次之。各阿特拉津处理对烷烃类化合物相对含量无明显影响,而烯烃类、胺类化合物相对含量均高于对照,说明阿特拉津促进了烯烃类、胺类的分泌。低浓度(0.2 mg·L−1和0.4 mg·L−1)阿特拉津抑制了酚类化合物分泌,而高浓度(2.0 mg·L−1)时相对含量大幅度增加(16.22%),是0.2 mg·L−1时相对含量(1.5%)的10.81 倍,说明高浓度阿特拉津提高了酚类的分泌量。
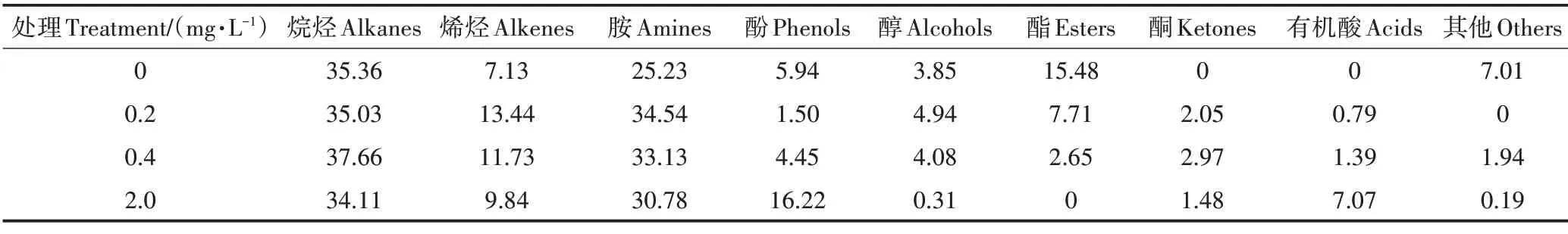
表2 阿特拉津胁迫下香蒲主要根系分泌物相对含量(%)Table 2 Relative content of main root exudates of cattail under atrazine stress(%)
醇类相对含量在低浓度(0.2 mg·L−1)处理下达最大值,较对照增加1.09 个百分点,最高浓度(2.0 mg·L−1)时达最小值,与对照相比降低3.54 个百分点,即低浓度阿特拉津促进醇类的分泌,而高浓度抑制醇类的分泌。阿特拉津胁迫下酯类化合物相对含量均低于对照,高浓度处理降低了酯类分泌。对照组未检测到酮类,各处理组对酮类相对含量的影响无明显规律。对照组未检测到有机酸类,而在0.2、0.4 mg·L−1和2.0 mg·L−1阿特拉津胁迫下,有机酸类的相对含量随着胁迫浓度的升高而增加。综上,高浓度阿特拉津抑制醇类及酯类的分泌,却刺激了有机酸类的分泌,且相对分泌量随胁迫浓度升高而增加。
根系分泌物是植物与环境进行物质交换和信息传递的重要媒介[28],任何影响植物生长和生理的过程均会引起根系分泌物数量和种类的改变[29]。本研究中,在阿特拉津胁迫下香蒲根系分泌的烷烃、烯烃、胺、酚、醇、酯、酮和有机酸类化合物中,烷烃类种类数最多,相对含量最大,这与不同磷浓度下香蒲检测到的主要根系分泌物类别相类似[30],但具体的化合物种类特征有所不同。本实验阿特拉津处理会刺激香蒲根系分泌化合物种类的增多。研究表明,植物在受到胁迫时会通过增加根系分泌物组分的释放来缓解毒害作用[31]。芦苇在盐胁迫下根系分泌物中检测出的化合物种类随着盐浓度的增加呈先升高后降低的趋势[20],这与本研究中香蒲根系的表现略有不同,可能与植物种类、污染物类型等因素有关[32]。值得一提的是,当阿特拉津浓度≥0.4 mg·L−1时,根系分泌的化合物种类数变化较小,说明阿特拉津胁迫虽可刺激分泌物的产生,但当胁迫超过一定程度后,会引起植物生长调节的代谢途径的紊乱,使生长受到抑制,根系分泌物组分数量的增加受到限制。
面对环境刺激,植物除了调整根系分泌物种类以外,还能通过改变含量来响应逆境胁迫[33]。本研究中根系分泌的各类化合物相对含量对阿特拉津胁迫的响应不同,其中,对醇类分泌量表现为低浓度促进高浓度抑制,对酯类显现出抑制效应,这两类物质均属于根系初生或次生代谢产物。王亚男等[34]对修复石油烃污染土壤的鸢尾进行根系代谢组学分析发现,石油烃改变了植物根系代谢物的含量,植物可通过调节自身代谢过程适应环境污染物的胁迫。研究表明,植物在有机污染物胁迫下会增加低分子量有机物的分泌以提高环境适应性[35]。王娇龙等[36]研究了紫玉兰在芘胁迫下根系分泌物的释放特征,发现芘胁迫提高了根系分泌物含氮化合物含量。本研究中阿特拉津胁迫刺激根系烯烃类、胺类化合物分泌,说明香蒲在受到阿特拉津胁迫时,通过根系自我调控分泌物的分泌量来适应环境,其中胺类属于植物次生代谢产物中的含氮有机物[37],植物为提高应对胁迫环境的适应性,分泌胺类次生代谢物作为适应的物质基础[38]。
本研究中高浓度阿特拉津刺激酚类的分泌,主要是因为植物产生的酚类化合物有利于阿特拉津的生物降解,为适应环境,植物根系提高酚类分泌进而增强阿特拉津的生物降解活性[39]。一些研究表明,植物处于有毒有害物质胁迫时也可通过增加根系有机酸的分泌来适应环境胁迫[40]。如除草剂异丙甲草胺浓度的增大导致了小麦根系分泌物中有机酸分泌量的升高[41]。本研究也发现了类似现象,可能是香蒲根系通过增加根系有机酸的分泌调节根际微域环境以缓解阿特拉津的毒性。
2.3 香蒲根系阿特拉津含量、根系抗氧化系统与根系主要分泌物的关系
以香蒲根系阿特拉津含量和抗氧化系统活性作为环境因子,根系主要分泌物相对含量作为物种变量进行RDA 分析,结果如图4 所示。由图可以发现2 类因子与阿特拉津胁迫下香蒲根系阿特拉津含量、抗氧化系统和分泌物响应有相关关系:香蒲根系阿特拉津含量与MDA含量呈正相关,与SOD活性呈负相关;根系阿特拉津含量、MDA 含量均与酚类和有机酸类的相对含量呈正相关,与酯类、醇类表现为负相关,而与烯烃类、胺类、酮类和烷烃类相关性不显著。
阿特拉津胁迫植物产生毒害作用的机理主要是抑制植物的光合作用[35]。碳水化合物是植物光合作用的主要产物,阿特拉津对光合作用的抑制会降低根系碳水化合物的代谢。碳水化合物是能量的主要来源,是植物生长发育的物质基础,与蛋白质代谢、脂质代谢、核酸代谢和生物量积累密切相关,同时关系到根系分泌物的产生[42]。当植物根系的碳水化合物代谢受到抑制时,植物生物量积累的过程受到影响,进而干扰植物根系分泌物的产生。WANG 等[42]利用非靶向代谢组学分析邻苯二甲酸二(2−乙基己基)酯(DEHP)胁迫对紫花苜蓿根系分泌物的影响,发现由于DEHP 对光合作用的抑制,使苜蓿根系碳水化合物代谢明显减弱,植株生物量下降,根系分泌物的组成和含量发生显著改变。
RDA 分析结果表明根系阿特拉津含量的增加会提高MDA 含量,促进有机酸类和酚类的分泌,从而抑制醇类和酯类的释放,该规律与其他研究结果一致。如ZHANG 等[33]对阿特拉津胁迫下狼尾草氧化应激及根系分泌物变化的研究发现,阿特拉津胁迫使植物体内活性氧大量积累,MDA 含量升高,且随着阿特拉津胁迫时间延长,根系碳水化合物代谢受到明显抑制,醇类含量降低,而脂肪酸代谢被激活,脂肪酸含量随之升高;DEBIANE 等[43]提出阿特拉津通过调控根系脂质组成和膜脂不饱和脂肪酸水平促进脂肪酸代谢以保护植株免受活性氧侵害的机理;WANG 等[42]的研究发现,苜蓿根系分泌物中脂肪酸含量在高浓度DEHP 胁迫下呈现升高的趋势。这表明污染物浓度和胁迫持续时间能够影响植物根系脂肪酸代谢,植物通过分泌更多有机酸来抵抗逆境胁迫。酚类物质是重要的植物次生代谢产物之一,众多研究发现,酚类化合物具有抗氧化性,可以参与活性氧的清除,预防氧化损伤,其含量的增加可以提高植物逆境防御能力和环境的适应性[44]。汪文云等[45]研究镉、芘单一及交互胁迫下红树植物秋茄根系酚类化合物的响应,结果表明根系酚类含量随污染物处理浓度增加而增大,从而增强了秋茄对污染胁迫的耐性。但也有研究表明,根系阿特拉津含量的增加诱发活性氧的过量积累和细胞膜脂过氧化,引起膜脂过氧化产物MDA 的增加,细胞膜结构被破坏,透性发生改变,相关代谢过程受阻,根溢泌的低分子量有机化合物数量和分泌量明显增加[46]。张福锁[47]的研究发现,缺锌导致小麦体内SOD 活性降低,根系细胞膜透性增加,低分子量有机化合物分泌量明显提高。目前,关于阿特拉津胁迫下根系分泌物酯类的变化研究较少。在逆境条件下酯类含量会发生改变,可作为胁迫响应信号。韦雪晶等[20]研究盐胁迫下芦苇根系分泌物的变化,发现分泌物中酯类含量随着盐浓度增加而降低。综上所述,阿特拉津胁迫诱导的氧化压力激活了植物体内抗氧化防御体系,引起植物生理代谢活动的变化,进而导致根系分泌物组成和含量的改变。这些变化可能是植物为减轻阿特拉津的毒害而进行的自我生理调节,也可能是植物体内某些代谢过程受阻,破坏了根系正常的生理活动,从而造成植物细胞膜结构受损,引起内部有机物质的渗漏,还可能是两者综合协调的结果,其具体机理还有待于进一步的研究。
3 结论
(1)香蒲可以在浓度≤2.0 mg·L−1阿特拉津长期胁迫下存活和生长,其根系对阿特拉津具有明显的吸收积累能力。
(2)在阿特拉津胁迫下香蒲根系SOD、CAT 和GSH活性表现出不同的变化趋势,反映了植物根系通过抗氧化活性酶间的相互协调来应对胁迫。在2.0 mg·L−1阿特拉津的长期胁迫下,香蒲根系抗氧化系统防御能力明显削弱,GSH 活性下降,SOD 活性显著低于对照,根系MDA含量明显升高。
(3)在阿特拉津胁迫下,香蒲根系的主要分泌物包括烷烃、烯烃、酯、胺、醇、酚、酮和有机酸类化合物,其中烷烃类化合物种类最多且相对含量最大。随着阿特拉津胁迫浓度升高根系分泌的化合物种类增多,酚类和有机酸类分泌量增大,而酯类和醇类的分泌量受到抑制。冗余分析结果显示,香蒲根系阿特拉津含量与MDA 含量呈正相关,两者均与酚类和有机酸类的相对含量呈正相关。
猜你喜欢
疯狂英语·新读写(2023年10期)2023-11-22 03:31:32
现代农业科技(2022年4期)2022-03-08 11:04:36
环境保护与循环经济(2021年7期)2021-11-02 08:10:52
家教世界·创新阅读(2021年8期)2021-08-23 14:50:30
甘肃农业大学学报(2019年4期)2019-09-18 09:14:08
现代园艺(2017年13期)2018-01-19 02:28:05
宁夏画报(2016年8期)2016-11-05 11:11:34
贵州农业科学(2016年10期)2016-04-11 09:13:19
华南农业大学学报(2015年5期)2015-12-04 03:04:38
花卉(2015年12期)2015-11-24 05:39:52