
矮化砧及对应中间砧苹果叶片光合对光照和CO2响应的模型模拟与评价
2022-01-20 02:15唐玉薇冯建荣杨伟伟
西北农业学报 2021年12期
唐玉薇,吴 彤,路 翔,王 凯,童 路,吴 佩,张 东,冯建荣,杨伟伟
(1.石河子大学 农学院,特色果蔬栽培生理与种质资源利用兵团重点实验室,新疆石河子 832000;2.西北农林科技大学 园艺学院,国家苹果改良中心杨凌分中心,陕西杨凌 712100)
光合作用是苹果碳的主要来源,直接影响果实的产量和品质[1-2],受环境因子、品种特性及栽培措施等因素的影响。其中,光和CO2是光合作用的主要能量来源和底物,是直接影响光合作用的环境因子[3-4]。栽培措施也会影响光合作用,如负载量调控、整形修剪和土肥水管理技术等。另外,嫁接砧木的类型会通过影响激素平衡[5]、源-库平衡[6-7]、氮吸收及分配[8-9],从而影响叶片光合能力。
目前,砧木类型对苹果叶片光合作用影响的研究非常多,但结果差异很大。有报道称,矮化砧木苹果叶片光合速率显著高于乔化砧木[10-11],而另外的一些研究结果则与之相反[12-13]。陈汝等[14]发现嫁接在M9上的苹果叶片净光合速率(Pn)高于其他M系列砧木。而Hayat[15]和张建光等[16]发现嫁接在M26上的苹果叶片光合效率高于M9。然而,果树的生产效率取决于冠层尺度光合生产力。由于树体结构差异,叶片尺度的光合能力并不能完全反映其冠层光合生产力。冠层光合生产力的准确计算及模拟主要基于叶片尺度的光合模型及冠层尺度的光截获模型[17-19]。应用叶片尺度光合模型对光合-光合有效辐射(Pn-PAR)和光合-二氧化碳(Pn-CO2)响应过程的模拟[20-22]可以获得许多重要的光合生理参数(如初始量子效率AQY、初始羧化效率η、最大净光合速率P nmax、光补偿点LCP和CO2饱和点CSP等)[23-24],可用于光合生理过程的评价及冠层光合生产力的模拟和评价[25-26]。基于叶片尺度的Pn-PAR和Pn-CO2过程的模拟大量应用于作物,可实现作物群体冠层光合生产力的精准模拟[27-28]。作为果树栽培的重要环节,砧木类型及砧木作为自根砧或中间砧是否会影响光合模型精度及适宜性,还未有报道。
当前中国苹果矮砧密植发展迅速,不同矮化砧叶片及冠层尺度的光合评价可为矮化砧木科学评价提供理论依据。亟需筛选适宜苹果不同砧木叶片尺度的光合模型。本试验以‘长富2号’苹果为试材,以M9和M26分别作为基砧和中间砧,基于现有光合模型,评价并筛选适宜于不同矮化砧木类型苹果叶片的光合模型,为砧穗组合的科学评价提供方法参考。
1 材料与方法
1.1 试验场地与材料
试验在陕西千阳县南寨镇西北农林科技大学千阳苹果试验示范站(北纬34.6°,东经106.9°,海拔883 m)进行。该地年平均降水量677.1 mm,无霜期197 d,年均日照时数约2 122.2 h,平均气温11.8℃,全年≥10℃活动积温3 462℃。2012年建园,品种为‘长富2号’,按照高纺锤形整形,2019年为树体生长第8年(包含苗龄),采用现代化矮砧集约种植模式,标准化管理。
根据矮化砧木类型,设置4个处理:M9、M26、M9/八棱海棠和M26/八棱海棠,株行距为1.3 m×4 m,采用水肥一体化滴灌方式灌溉。每处理设置3个生物学重复。土壤为砂壤土(有机质含量11.13 g·kg-1,碱解氮含量51.37 mg·kg-1,速效磷含量6.55 mg·kg-1,速效钾含量134.37 mg·kg-1)。
1.2 试验方法
1.2.1Pn-PAR和Pn-CO2响应曲线测定 试验于2019年8月进行,所有测量选择天气晴朗日照充足的9:00-11:00进行。每个处理随机选取树冠外围同一高度和方向、发育良好的成熟叶片,测定3个数值并记录,然后取3个数值的平均值作为基础数据,利用Li×6800(Li-Cor,Lincoln,Nebraska,USA)测 定Pn-PAR和Pn-CO2响 应曲线。Pn-PAR响应曲线的光合有效辐射(PAR)梯度:0、50、100、200、400、600、1 000、1 500和1 800μmol·m-2·s-1,叶室其他条件为标准条件(CO2=400μmol·mol-1,T≈30°C,VPD≤1.5 k Pa)。Pn-CO2响应曲线CO2的梯度:400、300、200、100、50、400、600、800、1 000、1 200、1 500和1 800μmol·mol-1,叶室其他条件为标准条件(PAR=1 500μmol·m-2·s-1,T≈30℃,VPD≤1.5 k Pa)。每处理测定3~5个叶片,每次测定读取3个稳定数值,取平均值。测定指标包括叶片的Pn、蒸腾速率(Tr)、胞间CO2浓度(Ci)和气孔导度(Gs)。
1.2.2Pn-PAR和Pn-CO2响应曲线模型Pn-PAR和Pn-CO2响应曲线的模拟基于现有成熟模型,模型类型、表达式及输入和输出值见表1。1.2.3 数据处理 运用R软件的‘nls’[34]对各砧木处理的Pn-PAR和Pn-CO2响应曲线进行模拟,估算模型参数。基于观测值和模拟值计算模型决定系数(R2)、均方根误差(RMSE)、平均绝对误差(MAE)和相对误差(RE),评估模型精度,各计算公式如下:

表1 叶片光合模型信息Table1 Leaf photosynthetic models information
式中:Y o为净光合速率实测值,¯Y o为净光合速率实测平均值,Y m为净光合速率模型模拟值,Y o,m为光合参数实测值,Y m,m光合参数模型模拟值,n为样本数。
使用Excel 2010进行数据整理和分析,使用Origin 8.5绘图。
2 结果与分析
2.1 不同矮化砧及对应中间砧苹果叶片P n-PAR响应曲线气体交换参数的比较
如图1所示,不同砧木处理苹果叶片Pn-PAR曲线气体交换参数总体变化趋势基本一致,随着PAR的增加,各处理苹果叶片的Pn、Gs和Tr均逐渐增大,Ci逐渐下降,但不同处理苹果叶片气体交换参数变化幅度存在一定的差异,中间砧组合的Pn、Gs和Tr整体显著高于自根砧组合,且M26/八棱海棠、M26的Pn、Gs和Tr分别显著大于M9/八棱海棠、M9。当PAR为400~1 500μmol·m-2·s-1时,各处理Pn差异显著,相同光强下,M26/八棱海棠和M26的Pn整体较高。当PAR超过1 500μmol·m-2·s-1时,M9/八棱海棠和M26的Pn随光强的增加而减少,出现光抑制现象,此时,M26/八棱海棠的Pn显著高于其他处理,M9的Pn最低。PAR为500~1 500μmol·m-2·s-1时,M26/八棱海棠Ci、Gs和Tr变化波动较大,在1 000μmol·m-2·s-1达到峰值,显著高于其他处理。
2.2 不同矮化砧及对应中间砧苹果叶片P n-PAR响应模型模拟精度比较
Pn-PAR模型拟合结果表明,非直角双曲线模型(NRH)和直角双曲线修正模型(MRH)模型对各处理的模拟精度高于直角双曲线模型(RH)和指数方程模型(EE)(表2)。NRH模型对M9/八棱海棠和M26的Pn-PAR响应曲线模拟得到的R2值 最 大,RMSE和MAE最 小;MRH模 型对M26/八棱海棠和M9的Pn-PAR响应曲线模拟得到的R2最大,RMSE和MAE最小。

表2 不同矮化砧木处理下苹果叶片各P n-PAR响应模型的模拟精度Table2 Accuracy of models in simulating P n-PAR response of apple leaves under different dwarfing rootstocks
基于Pn-PAR光响应曲线实测光合参数可知(表3),M26/八棱海棠和M26光合特性优于M9/八棱海棠和M9。M26/八棱海棠的P nmax最大,M26次之,分别比M9/八棱海棠和M9高6.8%、17.36%,M9的P nmax显著低于其他3个处理。各处理LCP表现为M26/八棱海棠和M9高于M9/八棱海棠和M26。LCP的大小顺序为M9>M9/八棱海棠>M26/八棱海棠>M26。M9/八棱海棠的R d最高,M26的R d最低。
通过比较基于光合模型估计的光合参数与实际测定的光合参数(表3),RH模型得到的各处理AQY偏差最大,比实测值高73.81%~92.11%,MRH模型得到的各处理AQY偏差次之,MRH模型拟合的P nmax最接近实测值。
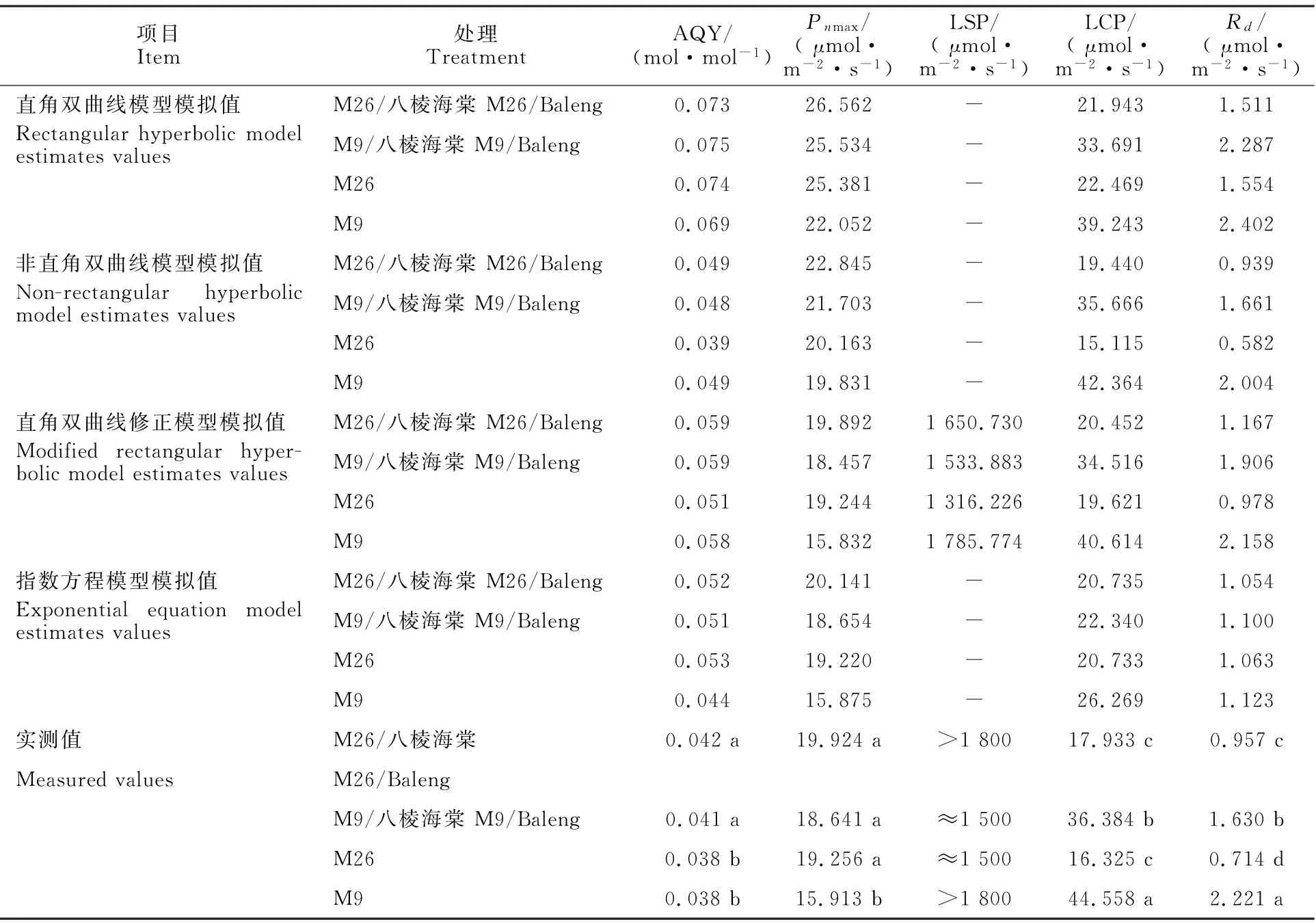
表3 不同矮化砧木处理下苹果叶片各P n-PAR响应模型主要光合参数的模拟值与实测值Table3 Model prediction and actually measured values of photosynthetic parameters related to P n-PAR response of apple leaves under different dwarfing rootstocks
与实测值相比,NRH模型LSP模拟值偏差2.194~0.718μmol·m-2·s-1,R d模拟值偏差0.01~0.217μmol·m-2·s-1,最接近实测值。
由表4可知,NRH模型对各处理AQY、LCP、R d模拟的RE最小。MRH模型对M26/八棱海棠、M9/八棱海棠、M26的P nmax模拟的RE最小,分别为0.002、0.023,而EE模型对M9的P nmax模拟的RE最小,为0.002。

表4 不同矮化砧木处理下苹果叶片各P n-PAR响应模型光合参数模拟值与实测值的相对误差比Table4 Relative errors of model predicted and actually measured values of photosynthetic parameters related to P n-PAR response of apple leaves under different dwarfing rootstocks
2.3 不同矮化砧及对应中间砧苹果叶片P n-CO2响应曲线气体交换参数比较
各砧木处理苹果叶片Pn-CO2曲线气体交换参数总体变化趋势基本一致(图2),但不同处理气体交换参数变幅存在一定差异。在低CO2浓度阶段(<600μmol·mol-1),各处理的Pn快速上升,上升趋势近似直线,其中,M26/八棱海棠和M26的Pn增长速度明显高于另外两个处理,之后随CO2浓度升高,在CO2浓度高于600 μmol·mol-1时M26/八棱海棠和M26的Pn上升速度均逐渐减缓,并在CO2浓度高于800 μmol·mol-1时,二者Pn值趋于稳定,而M9/八棱海棠和M9的Pn在此CO2浓度后增幅逐渐减小,并在CO2浓度高于1 000μmol·mol-1时二者Pn趋于稳定。
在CO2浓度低于300μmol·mol-1时,各处理间Ci差异不明显,随着CO2浓度的增加,各处理苹果叶片呈线性上升趋势,变幅为M26>M26/八棱海棠>M9>M9/八棱海棠,相同CO2浓度下Ci差异显著,M26/八棱海棠和M26分别高于M9/八棱海棠和M9。各处理的Gs和Tr整体均呈现缓慢下降的趋势,相同CO2浓度下,各处理Gs和Tr差异显著,M26的Gs和Tr最大,而M9/八棱海棠的Gs和Tr均显著低于其他处理。
2.4 不同矮化砧及对应中间砧苹果叶片P n-CO2响应模型模拟精度比较
从表5可以看出,各模型Pn-CO2响应曲线模拟精度不同。RH和M-M模型对各处理苹果叶片Pn-CO2响应曲线模拟精度相同。MRH模型对各处理Pn-CO2响应曲线光合参数模拟值与实测值间的R2最大,RMSE、MAE最小,精度最高。

表5 不同矮化砧木处理下苹果叶片各P n-CO2响应模型的模拟精度Table5 Accuracy of models in simulating P n-CO2 response of apple leaves under different dwarfing rootstocks
比较各砧木处理的苹果叶片光合参数实测值可知(表6),M26/八棱海棠和M26的η分别显著高于M9/八棱海棠和M9,而P nmax和CSP相反。
M9的CCP最大,M9/八棱海棠次之,M26/八棱海棠最小。各处理的R p大小顺序为M9/八棱海棠<M26/八棱海棠<M26<M9。
从表6可知,RH和M-M模型中各处理的P nmax、CCP和R p模拟值完全一致,且这2种模型均无法估计CSP。RH模型可估计η,而M-M模型则无法估计。MRH模型可估计各处理苹果叶片Pn-CO2主要光合参数,其P nmax和CSP估计值与实际值接近,但η和R p模拟值均大于实测值,且偏差较大。

表6 不同矮化砧木处理下的苹果叶片各P n-CO2响应模型主要光合参数模拟值与实测值比较Table6 Model of predicted and actually measured values of major photosynthetic parameters related to P n-CO2 response of apple leaves under different dwarfing rootstocks
为精确判断上述3种模型对各处理苹果叶片Pn-CO2曲线主要光合参数拟合的模型拟合精度,本研究同样根据各处理苹果叶片的Pn-CO2响应曲线主要光合参数模拟值与实测值的RE进行定量评价(表7),可得MRH模型对各处理的P nmax、CCP、R p的模拟RE值最小。
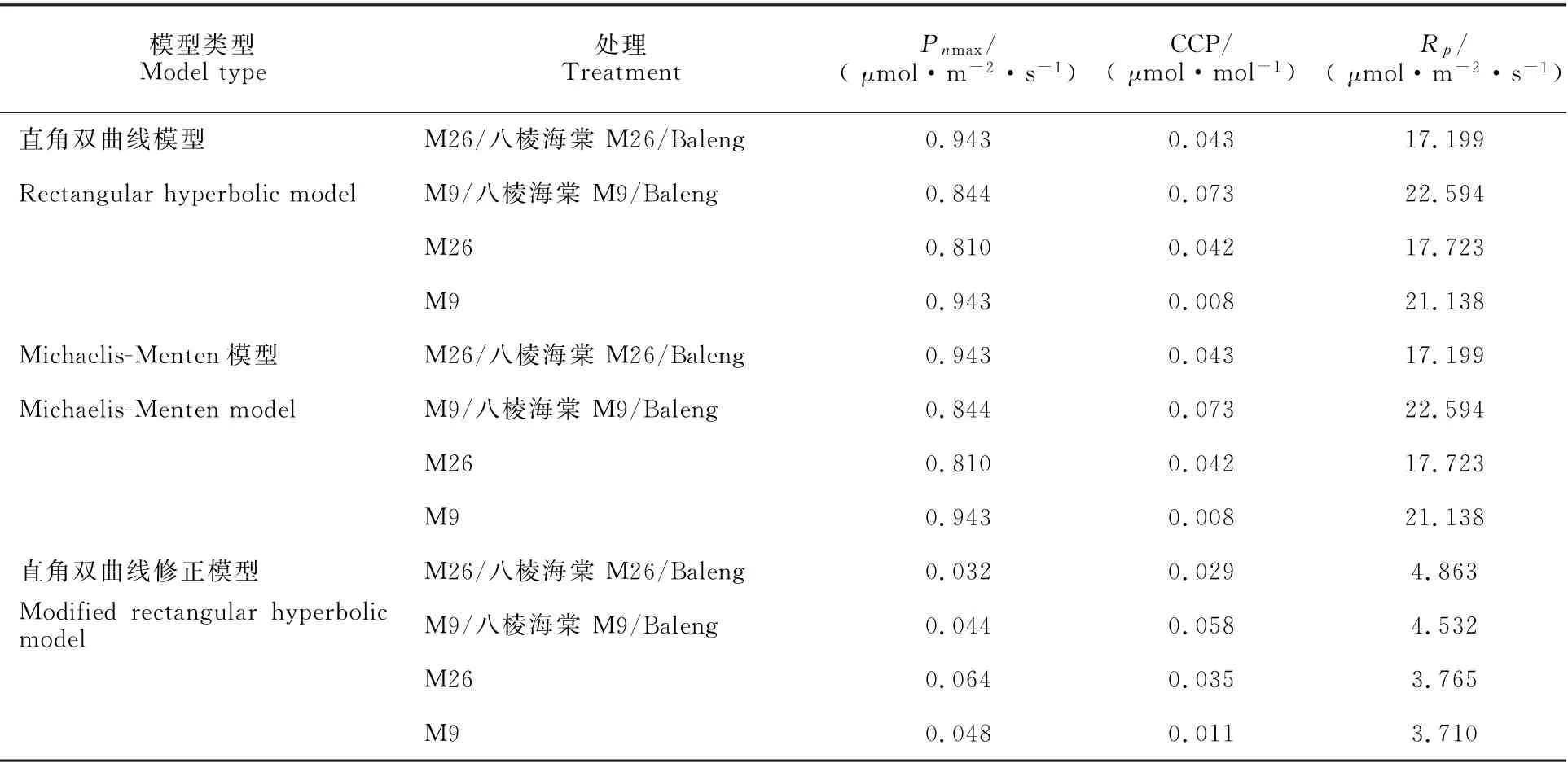
表7 不同矮化砧木处理下苹果叶片各P n-CO2响应模型主要光合参数模拟值与实测值的相对误差Table7 Relative errors of model predicted and actually measured values of major photosynthetic parameters related to P n-CO2 response of apple leaves under different dwarfing rootstocks
3 讨论
3.1 不同矮化砧及对应中间砧苹果叶片P n-PAR和P n-CO2适宜 模型筛 选
本试验中,各模型的R2相对较高(R2>0.99),说明用于模拟不同砧木处理苹果叶片的光合模型具有一定合理性,但R2只表示观测值与模拟值间相关性高,可能存在模拟值统一偏高或偏低的情况[35]。因此,通过引入RMSE和MAE可进一步对模型精度定量评估,RMSE、MAE越小、R2越接近于l,表明模拟精度越高[36-37]。
本文4种模型中(表3和表4),NRH模型与MRH模型对各砧木处理Pn-PAR曲线模拟精度较高。前人利用3个经验模型(RH、NRH和MRH模型)对山杏、油松、沙棘的Pn-PAR响应曲线进行模拟,结果表明,NRH和MRH拟合的总体效果要好于RH[38-40]。但适宜模拟苹果自根砧和中间砧的Pn-PAR响应模型不同,其中NRH模型对M26/八棱海棠和M9/八棱海棠的Pn-PAR响应曲线模拟精度高,效果最好,MRH对M26和M9的Pn-PAR响应曲线模拟精度高,效果最好(表4)。也有研究表明,NRH模型对梭梭的效果最好而RH模型对柽柳拟合效果最好[41]。产生上述现象的原因可能是由于物种差异,生理特征不同,导致适宜的Pn-PAR模型不同。由表4和表5可知,MRH模型对各类型砧木处理Pn-CO2曲线模拟精度高于RH和M-M模型。
各类光合模型各有优缺点,应对比选择最佳模型估算光合参数。对于夏玉米,研究人员发现基于其Pn-PAR曲线,RH与NRH模型会高估叶片AQY和P nmax[18]。本试验中,RH模型估计得到的叶片AQY的RE大于0.73,P nmax的RE大于0.31。由表4可知,RH、EE、NRH模型无法直接估算LSP,主要由于RH、EE、NRH模型均是没有极值的渐近线,Pn-PAR响应曲线主要光合参数必须借助其他方法进行估算,但是这样得到的LSP远低于实际测量值[42]。MRH模型为RH模型的修正模型,可估计Pn-PAR响应曲线各主要光合参数,且最接近实测值[43]。本试验中,适宜估计不同砧木处理光合参数的模型不同。M26/八棱海棠和M9/八棱海棠、M26的AQY、LCP、R d估计优先选择NRH模型,P nmax求解优先选择MRH。M9的AQY、P nmax求解优先选择EE模型,LCP、R d求解优先选择MRH模型(表3)。前人研究发现RH模型和M-M模型本质上是相同的,因而二者Pn模拟值完全一致[37],本试验中通过对比2个模型的模拟精度发现,二者模拟精度完全一致(表5)。M-M模型本身无η这个参数,所以无法估计η,MRH模型、RH模型估计的η和R p的RE较大。但MRH模型可克服RH模型未考虑曲线凸度缺陷及无法拟合CO2饱和现象和Pn下降的缺点,能够反映出光合作用的CO2饱和现象,并得出CSP,且与实测值较接近。从这点看,MRH模拟对各砧木Pn-CO2曲线主要光合参数的模拟优于其他2种模型。M26/八棱海棠和M9/八棱海棠、M26的P nmax、CCP、CSP求 解 优 先 选 择NRH模 型。M9的P nmax、CCP求解优先选择NRH模型,而CSP求解优先选择M-M模型(表6)。
3.2 基于P n-PAR和P n-CO2响应曲线的不同矮化砧及对应中间砧苹果叶片光合特性比较
在低光强范围内(<600μmol·m-2·s-1),M26/八棱海棠和M26的Pn高于M9/八棱海棠和M9(图1),表明弱光条件下,前者的光合效率要强于后者,适应弱光的能力强。当PAR增强到一定程度后,由于光抑制现象,M9/八棱海棠和M26的Pn停止增长,出现明显的下降,而M26/八棱海棠和M9未出现光饱合或者光抑制现象。
Pn-PAR响应曲线主要光合参数能较好地反映逆境条件下植物的光合潜能、光能利用率及光抑制水平高低等特性[44]。由表4知,M26/八棱海棠和M26具有较低的AQY、LCP和LSP,对强光和弱光的适应性均比M9/八棱海棠和M9强,能利用较弱的太阳辐射进行光合作用且又能适应阳光较充足的环境,同时P nmax较高而R d较低,可减少碳损耗,这是其对不同光环境做出的适应性反应和采取的生理适应机制[45]。
各处理苹果叶片Pn-CO2曲线气体交换参数总体变化趋势基本一致(图2),但不同处理气体交换参数变幅存在一定的差异。其中,M26为0~600μmol·mol-1时Gs和Ci,Tr呈现相反的变化趋势,说明M26在此阶段Ci和Tr受到Gs的影响,但并未对整体Ci、Tr变化趋势造成决定性影响。
Pn-CO2响应曲线主要光合参数是反映叶中Rubisco含量及活性和CO2利用效率的重要指标[46]。本试验中,M26/八棱海棠和M26的Rp、P nmax、CCP和CSP均低于M9/八棱海棠和M9,而η较高(表8),表明M26/八棱海棠和M26叶片对低CO2浓度环境的适应性更好,增大CCP和CSP范围,提高CO2利用率[47]。
4 结论
综上所述,基于叶片尺度光合模型对不同矮化砧及对应中间砧苹果叶片光合能力进行评价时,可优先考虑使用非直角双曲线模型和直角双曲线修正模型。使用M26作为基砧和中间砧时,有效提高‘长富2号’的耐弱光能力和低CO2浓度利用效率,光合生产潜力大。但不同砧木亦会影响冠层结构,笔者将进一步耦合叶片尺度光合模型和冠层结构及光截获模型,在冠层尺度深入研究矮化砧木对苹果冠层结构和功能的影响。
猜你喜欢
中国果树(2022年5期)2022-06-09
节水灌溉(2021年9期)2021-10-13
快乐作文(1.2年级)(2021年2期)2021-09-10
中国科技纵横(2021年13期)2021-09-06
落叶果树(2021年6期)2021-02-12
中国房地产业·下旬(2020年12期)2020-01-11
快乐作文(1.2年级)(2019年12期)2019-09-10
快乐作文(1.2年级)(2019年8期)2019-09-10
新农业(2017年3期)2017-05-17
大陆桥视野·下(2016年11期)2017-02-28
