
耕层重构对棉田土壤养分、微生物数量与酶活性的影响
2022-01-07 08:41王燕张谦王树林韩硕冯国艺董明钱玉源祁虹
棉花学报 2021年5期
王燕,张谦,王树林,韩硕,冯国艺,董明,钱玉源,祁虹*
(1.河北省农林科学院棉花研究所/农业部黄淮海半干旱区棉花生物学与遗传育种重点实验室/国家棉花改良中心河北分中心,石家庄 050051;2.河北省邢台广播电视大学,河北 邢台 054000)
棉花是河北省主要经济作物之一,主要在河北省中南部种植,耕地方式以传统旋耕为主。连年旋耕导致土壤耕层变薄、土壤紧实化、养分分布不均等,影响棉花的生长发育[1-3]。为解决这一问题,王树林等[4-5]提出耕层重构这一新的土壤耕作方式,是将0~20 cm与20~40 cm土壤互换,同时松动40 cm以下土壤,重新构筑耕层。前期研究发现,土壤耕层重构可以降低土壤容重,平衡土壤养分垂直分布,提高深层土壤蓄水保墒能力[6],改善棉花的生长环境,促进棉花根系的下扎和延伸[4,7],显著缓解常年旋耕导致的土壤结构问题。但是,耕层重构对棉田土壤微生态环境的影响尚不明确。
土壤微生物是土壤环境的重要组成部分,对其生存的土壤环境非常敏感,能对土壤生态功能变化和环境胁迫迅速做出反应,可以作为反映土壤耕作方式和环境变化的重要指标[8]。土壤酶活性可以表征土壤的综合肥力及土壤养分转化过程,反映土壤中各种生化过程的强度和方向,可作为土壤质量的生物学评价指标。土壤结构、土壤微生物群落结构和土壤酶活性等微生态环境受耕作模式影响[9-11]。前人研究发现,与免耕相比,传统旋耕会降低0~20 cm土层真菌数量和土壤微生物生物量碳含量[12],而深松和深翻等土壤耕作方式会显著增加土壤中细菌、真菌和放线菌数量[13]。与旋耕和免耕相比,深耕和深松可打破土壤犁底层,降低土壤容重[14],提高土壤脲酶活性[12]。也有研究表明,相比传统耕作,保护性耕作能显著提高土壤总体酶活性[15]。作为一种新的土壤耕作方式,研究耕层重构对棉田土壤微生态环境的影响对明确其作用机理具有重要意义。
本研究以常年旋耕处理为对照,研究耕层重构对棉田土壤养分、微生物数量和酶活性的影响,并进行相关性分析,明确耕层重构技术对土壤微生态环境的影响,为耕层重构技术的应用提供理论基础。
1 材料与方法
1.1 试验设计
试验于2019年在河北省威县试验站(威县枣元乡东张庄村)进行。试验田为连作棉田,土壤为砂壤土,中等肥力,土壤的基础地力为0~20 cm土层含有机质11.3 g·kg-1、全氮0.9 g·kg-1、速效磷45.6mg·kg-1、速效钾200.8mg·kg-1。供试材料为河北省农林科学院棉花研究所选育的冀棉315。
常规旋耕(Rotary tillage)为对照(CK),旋耕深度15 cm。耕层重构(Restructuring soil layers treatment,T)是将0~20 cm土层土壤与20~40 cm土层土壤互换,同时铲松40~60 cm土层土壤,然后旋耕。具体做法是用铁锹先将0~20 cm土壤移至一处,再将20~40 cm土壤移至另一处,用铁锹铲松40~60 cm土壤,然后先回填0~20 cm土壤,再回填20~40 cm土壤。采用随机区组设计,小区长8m,宽6m,面积48m2,3次重复。4月16日小区撒施复合肥(N、P2O5、K2O质量分数为15%、13%、17%)750 kg·hm-2,并灌底墒水,灌水量为1 200 m3·hm-2;4月24日CK和T处理用微耕机旋耕,所有小区耙耱后播种。4月25日播种,采用地膜覆盖,76 cm等行距种植棉花,留苗密度为7.5 万株·hm-2,10月30日收获。田间管理措施同当地大田。
分别于棉花苗期、蕾期、花铃期和吐絮期对土壤取样,每个小区采用五点法取样,在棉花行间先刮去表层土,然后用土钻分别取0~20 cm、20~40 cm、40~60 cm和60~80 cm土样,随机取5个点混合装袋。混匀过筛后,分为2份:一部分放入冰盒中带回实验室,放入4℃冰箱暂存,用于测定土壤微生物数量;另一部分经常温风干后磨细过筛,用于测定土壤养分含量和土壤酶活性。
1.2 测定项目及方法
1.2.1土壤养分测定。土壤全氮含量测定采用浓硫酸消煮-半微量凯氏法,碱解氮含量测定采用碱解扩散法;全磷含量测定采用氢氧化钠熔融-钼锑抗比色法,速效磷含量测定采用碳酸氢钠浸提-钼锑抗比色法;全钾含量测定采用碳酸钠熔融-火焰光度计法,速效钾含量测定采用乙酸铵浸提-火焰光度计法。有机质含量测定采用重铬酸钾容量法-外加热法[16]。
1.2.2土壤微生物数量测定。微生物数量测定用稀释平板计数法。细菌、真菌和放线菌分别用牛肉膏蛋白胨培养基、马丁氏培养基和高氏Ⅰ号培养基[17]。用涂抹法接种,在28℃条件下分别培养2 d、3 d和7 d,培养结束后取出,选择菌落总数为10~200的培养皿进行统计和计算。
1.2.3耕层重构处理土壤微生物数量。耕层重构处理将0~20 cm土层和20~40 cm土层土壤互换,因此计算0~20 cm和20~40 cm土层微生物数量时引入修正系数r以消除本底差异。T处理土壤微生物数量=土壤微生物的测定数量×修正系数。其中T处理0~20 cm土层的修正系数r=苗期CK处理20~40 cm土层的微生物数量/苗期T处理0~20 cm土层的微生物数量,T处理20~40 cm土层的修正系数r=苗期CK处理0~20 cm土层的微生物数量/苗期T处理20~40 cm土层的微生物数量。
1.2.4土壤酶活性测定。脲酶活性测定采用苯酚钠次氯酸钠显色法,碱性磷酸酶活性测定采用磷酸苯二钠比色法,蔗糖酶活性测定采用3,5-二硝基水杨酸比色法[18]。
1.2.5棉花产量及生物量测定。各小区单独收获测产。在棉花收获期每小区随机取5株棉花地上部,于105℃杀青30m in,然后85℃烘干至恒重测定地上部生物量。
1.3 统计分析方法
试验数据采用M icrosoft Excel2003和SPSS 19.0 软件进行处理和分析,均采用t测验分析差异显著性。
2 结果与分析
2.1 耕层重构对土壤养分含量的影响
耕层重构对吐絮期棉田土壤养分含量的影响如表1所示。与CK相比,吐絮期T处理棉田0~20 cm土层的全氮、全磷、碱解氮、速效磷、速效钾和有机质含量显著降低27.3 8%、25.4 7%、33.6 8%、64.7 8%、34.6 3%和29.6 3%,20~40 cm土层的全氮、全磷、碱解氮、速效磷、速效钾和有机质含量显著增加5.2 6%、16.9 0%、5.7 5%、71.4 1%、29.8 5%和56.7 6%,40~60 cm土层的全氮、全磷、速效磷、速效钾和有机质含量显著增加5.3 6%、6.3 5%、163.3 8%、19.5 6%和23.8 5%,60~80 cm土层的全氮、全钾、碱解氮和速效钾含量显著增加6.2 5%、4.9 4%、7.7 0%和21.1 9%。T处理棉田20~40 cm土层的全磷、碱解氮、速效磷和有机质含量均高于0~20 cm土层,而对照20~40 cm土层的养分含量低于0~20 cm土层。说明耕层重构显著增加了20~40 cm土层除全钾外的养分含量,深层土壤(60~80 cm)的养分含量也有增加趋势,解决了常年旋耕导致的营养物质富集在表层的问题,使土壤养分含量分布更加均匀。
2.2 耕层重构对土壤微生物数量及分布的影响
2.2.1细菌。细菌是土壤微生物的主要组成部分,一般认为土壤细菌数量的增加是土壤肥力提高的重要生物学标志。耕层重构对棉田土壤细菌数量(以菌落形成单位,即colony form ing units,cfu计算)的影响如表2所示。在0~20 cm土层中,棉花蕾期和花铃期T处理细菌数量比CK的显著降低55.7 3%和56.4 4%;在20~40 cm土层,棉花蕾期、花铃期和吐絮期T处理的细菌数量分别是CK的2.30 、5.09 和1.99 倍,差异均达到显著水平;在40~60 cm土层中,棉花花铃期T处理的细菌数量比CK显著增加350.3 9%;在60~80 cm土层中,棉花花铃期T处理的细菌数量比CK的显著增加117.3 8%。CK处理的土壤细菌主要集中在0~20 cm土层,而T处理20~40 cm土层的细菌数量大量增加。说明与传统旋耕相比,耕层重构降低了表层土壤细菌数量而增加了较深土层细菌数量,改变了细菌的垂直分布。这是因为耕层重构处理把细菌含量高的表层土翻到下面,增加了20~40 cm土层的细菌数量;另外,耕层重构改变了土壤的理化性状,提高了20~40 cm土层的养分含量,更利于细菌的生长。
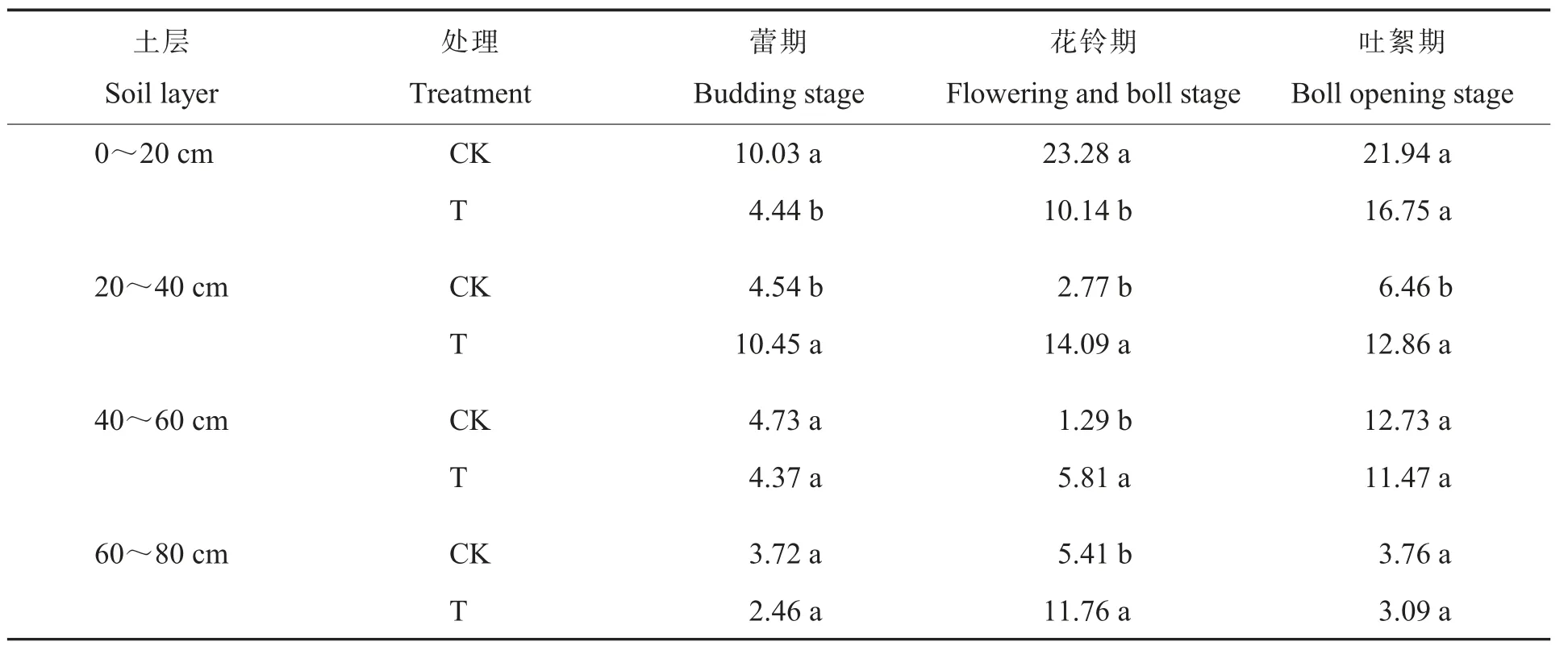
表2 耕层重构对棉田土壤细菌数量的影响Table 2 Effect of restructuring soil layers on bacterial quantity in different soil layers of cotton field(107 cfu·g-1)
2.2.2真菌。真菌在土壤微生物区系中数量最少,却是许多作物病害的病原菌,与作物土传病害的发生直接相关,在土壤中作用不容忽视。耕层重构对棉田土壤真菌数量的影响如表3所示。在0~20 cm土层中,棉花蕾期和吐絮期时,T处理的真菌数量比CK分别显著降低31.9 6%和40.8 4%;在20~40 cm土层中,棉花蕾期、花铃期和吐絮期T处理的真菌数量分别是CK的2.49倍、4.04 倍和2.83 倍,差异均达到显著水平;在40~60 cm土层中,棉花蕾期和花铃期T处理的真菌数量分别是CK的10.93 倍和11.38 倍,差异均达到显著水平。与CK处理真菌主要集中在0~20 cm土层中不同,T处理中20~40 cm土层也含一定数量真菌。说明耕层重构降低了表层土壤真菌数量而增加了较深土层真菌数量,改变了土壤真菌的垂直分布。

表3 耕层重构对棉田土壤真菌数量的影响Table 3 Effect of restructuring soil layers on fungi quantity in different soil layers of cotton field(105 cfu·g-1)
2.2.3放线菌。放线菌能分解土壤中多数细菌和真菌不能分解的化合物。土壤放线菌数量的多少,关系着土壤代谢强度的高低。耕层重构对棉田土壤放线菌数量的影响如表4所示。在0~20 cm土层中,棉花蕾期T处理的放线菌数量比CK显著降低53.7 1%;在20~40 cm土层中,棉花蕾期、花铃期和吐絮期T处理的放线菌数量比CK显著增加72.9 4%、87.1 4%和79.5 3%;在40~60 cm土层中,棉花蕾期和花铃期T处理的放线菌数量比CK显著增加67.2 1%和197.5 0%。CK处理的土壤放线菌主要集中在0~20 cm土层,而T处理20~40 cm土层的放线菌数量大幅增加,与细菌和真菌变化一致。说明耕层重构降低了表层土壤放线菌数量而增加了较深土层放线菌数量,改变了放线菌的垂直分布。
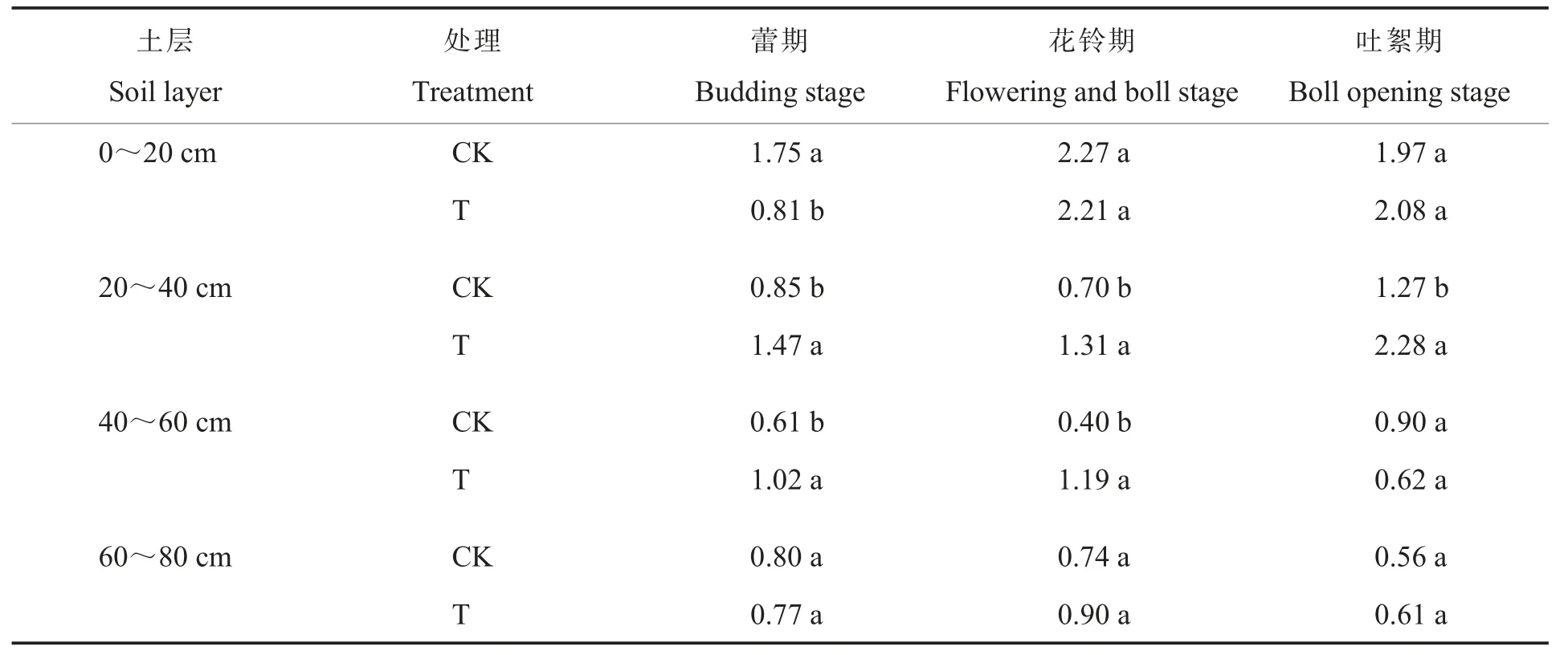
表4 耕层重构对棉田土壤放线菌数量的影响Table 4 Effect of restructuring soil layers on actinomycetes quantity in each layer of cotton field(107 cfu·g-1)
2.3 耕层重构对土壤酶活性的影响
2.3.1脲酶。脲酶能分解有机物生成氨和二氧化碳,其活性反映土壤的氮素供应状况。耕层重构对棉田土壤脲酶活性的影响如表5所示。在0~20 cm土层中,棉花苗期、花铃期和吐絮期T处理的脲酶活性分别比CK显著降低44.5 6%、37.5 5%和32.7 8%;在20~40 cm土层中,棉花苗期、花铃期和吐絮期T处理的脲酶活性分别比CK显著增加26.4 3%、58.4 5%和11.4 6%;在40~60 cm土层中,棉花蕾期和花铃期T处理的脲酶活性分别比CK显著增加12.3 4%和42.6 2%;在60~80 cm土层中,棉花蕾期和吐絮期T处理的脲酶活性分别比CK显著增加22.6 5%和16.4 0%。说明耕层重构处理降低了表层土壤脲酶活性,而提高了深层土壤脲酶活性。CK处理的脲酶活性随着土壤深度的增加而呈降低趋势,随着棉花生育时期的推进呈先降低后升高趋势(20~40 cm土层除外)。除蕾期外,T处理脲酶活性随着土壤深度的增加呈先上升后下降趋势,其中20~40 cm土层脲酶活性最高,可能与20~40 cm土层微生物数量和养分含量的升高有关。
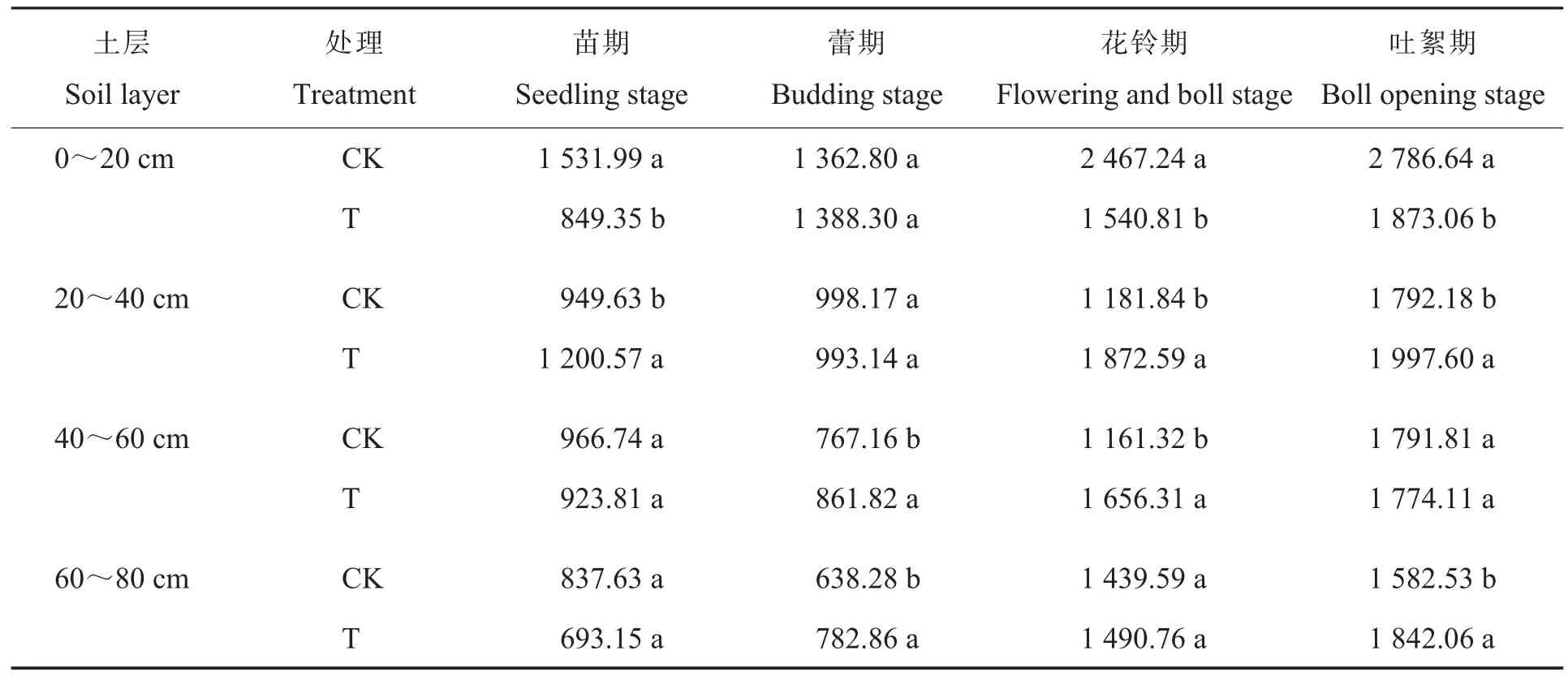
表5 耕层重构对棉田土壤脲酶活性的影响Table 5 Effect of restructuring soil layers on the urease activity in each layer of cotton field (U·g-1)
2.3.2碱性磷酸酶。碱性磷酸酶是能催化土壤有机磷化合物矿化的酶,其活性是评价土壤磷素生物转化方向与强度的指标。耕层重构对棉田土壤碱性磷酸酶活性的影响如表6所示。与CK相比,在0~20 cm土层中,棉花苗期、蕾期、花铃期和吐絮期T处理的碱性磷酸酶活性分别显著降低52.2 8%、38.8 2%、52.2 3%和45.1 3%;在20~40 cm土层中,棉花苗期、蕾期、花铃期和吐絮期T处理的碱性磷酸酶活性分别比CK显著增加90.0 8%、19.0 5%、23.4 6%和30.8 4%;在40~60 cm土层中,棉花苗期和吐絮期T处理的碱性磷酸酶活性分别比CK显著增加289.9 4%和14.0 0%;在60~80cm土层中,棉花苗期T处理的碱性磷酸酶活性较CK显著增加29.7 9%。说明耕层重构处理降低了表层土壤碱性磷酸酶活性而提高了深层土壤碱性磷酸酶活性。CK处理的碱性磷酸酶活性随着土壤深度的增加而呈降低趋势。T处理的碱性磷酸酶活性随着土壤深度的增加而呈先增加后下降趋势,可能与土壤养分和微生物数量的垂直分布改变有关。
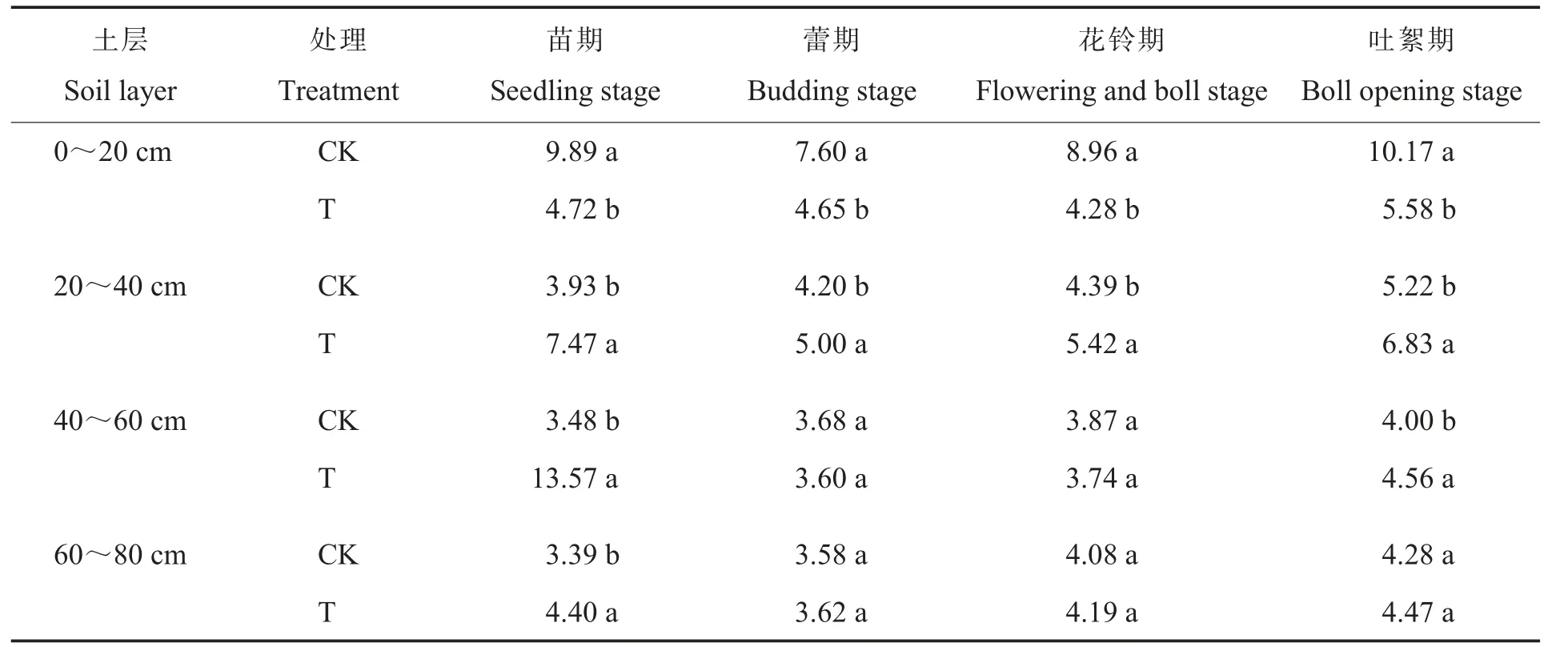
表6 耕层重构对棉田土壤碱性磷酸酶活性的影响Table 6 Effect of restructuring soil layers on the alkaline phosphatase activity in each layer of cotton field(U·g-1)
2.3.3蔗糖酶。蔗糖酶能够把蔗糖水解为单糖从而被机体吸收,蔗糖酶是评价土壤肥力的重要指标。耕层重构对棉田土壤蔗糖酶活性的影响如表7所示。在0~20 cm土层中,棉花苗期、蕾期、花铃期和吐絮期T处理的蔗糖酶活性分别比CK显著降低73.1 8%、25.6 3%、43.4 9%和53.5 3%;在20~40 cm土层中,棉花蕾期、花铃期和吐絮期T处理的蔗糖酶活性分别比CK显著增加134.7 2%、116.5 4%和72.4 2%;在40~60 cm土层中,棉花吐絮期T处理的蔗糖酶活性比CK显著增加45.0 3%。与土壤脲酶和碱性磷酸酶变化趋势相似,耕层重构处理降低了表层土壤蔗糖酶活性而提高了较深土层蔗糖酶活性。
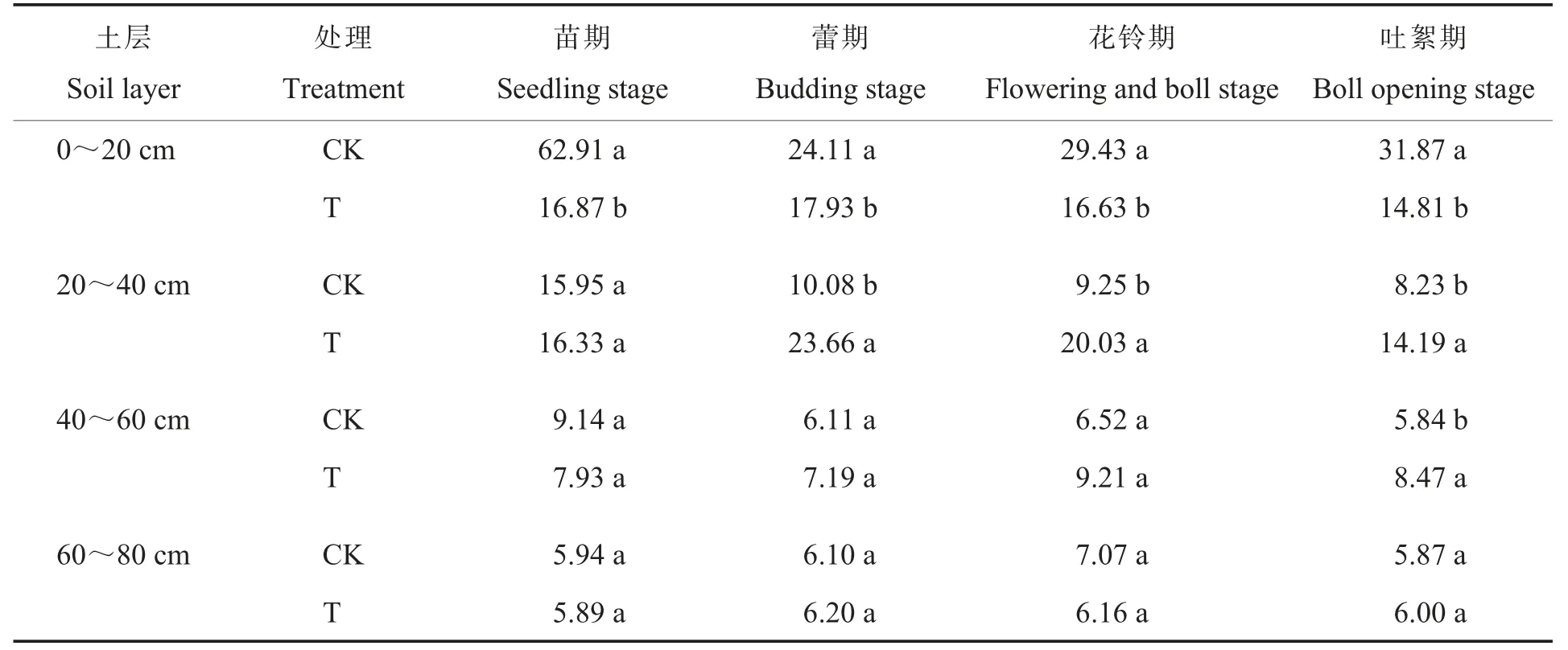
表7 耕层重构对棉田土壤蔗糖酶活性的影响Table 7 Effect of restructuring soil layers on the sucrase activity in each layer of cotton field(U·g-1)
2.4 相关性分析和回归分析
采用SPSS 19.0 软件对土壤微生物数量、土壤酶活性和土壤养分进行简单相关性分析和回归分析。设X1为细菌数量(107cfu·g-1),X2为真菌数量(105cfu·g-1),X3为放线菌数量(107cfu·g-1),X4、X5和X6分别为脲酶活性、碱性磷酸酶活性和蔗糖酶活性(U·g-1),Y1为全氮含量(g·kg-1),Y2为全磷含量(g·kg-1),Y3为全钾含量(g·kg-1),Y4为有机质含量(g·kg-1),Y5为碱解氮含量(mg·kg-1),Y6为速效磷含量(mg·kg-1),Y7为速效钾含量(mg·kg-1),进行相关性分析,结果见表8。土壤中的全氮含量与脲酶活性、碱性磷酸酶活性、蔗糖酶活性显著正相关,全磷含量与细菌数量、真菌数量、放线菌数量、脲酶活性、碱性磷酸酶活性、蔗糖酶活性显著正相关,有机质含量、碱解氮含量和速效钾含量与真菌数量、脲酶活性、碱性磷酸酶活性、蔗糖酶活性均显著正相关,速效磷含量与真菌数量、放线菌数量、脲酶活性、碱性磷酸酶活性和蔗糖酶活性显著正相关。土壤脲酶活性、碱性磷酸酶活性和蔗糖酶活性三者之间呈极显著正相关。这说明本研究中的3种酶在促进土壤物质转化和能量交换时,不仅显示其专有特性,还存在共性关系,也说明土壤中含氮有机物、有机碳和磷素的转化之间是相互影响的。此外,细菌数量与蔗糖酶活性显著正相关,真菌和放线菌数量均与脲酶活性、碱性磷酸酶活性、蔗糖酶活性显著正相关。
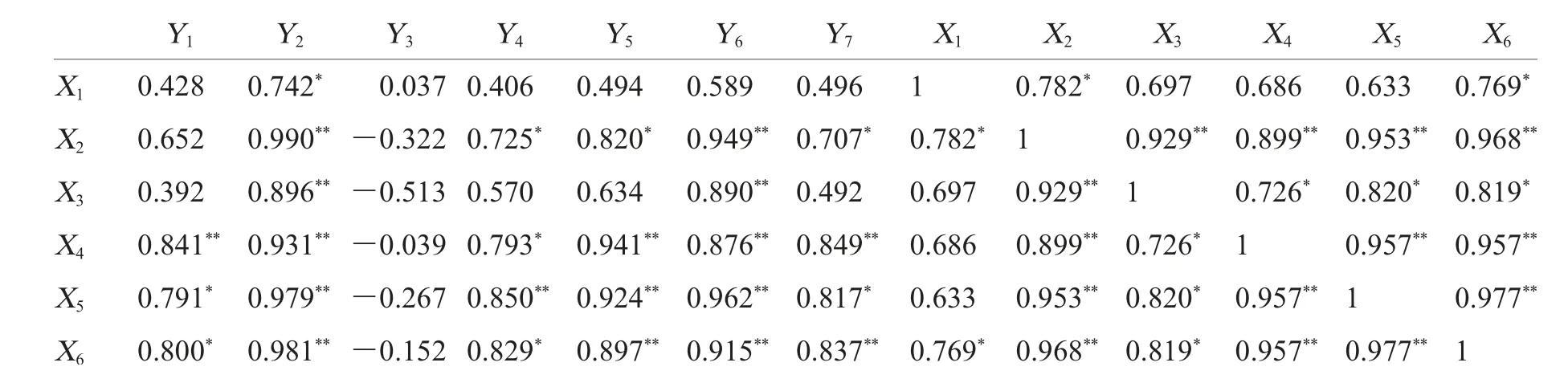
表8 土壤微生物数量、土壤酶活性和土壤养分之间的相关性Table 8 Correlative coefficient between m icrobial quantities,soil enzyme activities and soil nutrients in restructuring soil layers
进一步进行回归分析,得出以下较理想的回归 模 型:Y1=0.232 +0.000209 X4,R2=0.708**,F=14.541 ;Y2=0.476 +0.06 X2+0.03X5,R2=0.994 *,F=427.048 ;Y4=3.52 +0.709X5,R2=0.722 **,F=15.595 ;Y5=5.375 +0.021X4,R2=0.885 **,F=46.349 ;Y6=-26.32 +7.536X5,R2=0.925 **,F=74.117 ;Y7=-28.663 +0.08X4,R2=0.721 **,F=15.521 。
上述模型R2数值后的*和**分别表示在0.05 和0.01 水平上显著相关。由此可知,土壤全氮、碱解氮和速效钾含量与土壤脲酶活性显著正相关,有机质和速效磷含量与碱性磷酸酶活性显著正相关,全磷含量与真菌数量和碱性磷酸酶活性显著正相关,土壤全钾含量与土壤微生物数量和土壤酶活性没有显著相关关系。土壤脲酶活性、碱性磷酸酶活性和真菌数量是反映土壤肥力的重要指标。
2.5 耕层重构对棉花产量和生物量的影响
T处 理 的 籽 棉 产 量(4 500.64 kg·hm-2)和CK(4 605.05 kg·hm-2)没有显著差异,但是T处理的棉花地上部生物量(11 733.05 kg·hm-2)比CK(10 773.05 kg·hm-2)增加8.9 1%,差异达到显著水平。说明耕层重构可以促进棉花生长,提高棉花地上部生物量。
3 讨论
3.1 耕层重构对棉田土壤养分含量的影响
耕作方式会对土壤结构和养分等产生深刻的影响,还会改变土壤微生物群落特征,决定了土壤质量变化的方向和程度[19]。目前中国农业生产中多种耕作方式并存,比较常见的有免耕、少免耕、旋耕、翻耕、深松、垄作、秸秆还田覆盖,且不同耕作方式、耕翻深度及年际可以变化组合,而不同耕作方式及组合对土壤的影响是不同的[1]。研究发现,保护性耕作下土壤耕作层的氮、磷、钾等养分含量显著提高,且在0~10 cm土层富集,但在10~30 cm土层没有显著变化[1,20]。也有研究发现,翻耕处理下土壤养分含量最高,其次是旋耕处理,而免耕处理下土壤养分含量最低,且土壤容重越高则土壤养分含量越低[21]。王树林等认为耕层重构处理降低了20~40 cm土层的容重,增加了该土层的养分含量[7]。在本研究中,耕层重构处理将0~20 cm土层和20~40 cm土层互换,同时松动40~60 cm土层,降低了20~40 cm土层容重,同时把土壤养分富集的表层土转置到下层,把养分含量不高的下层土转置到上层,从而使0~20 cm土壤养分含量低于对照,而20~80 cm土壤养分含量高于对照,与前人研究结果[7]一致。说明耕层重构后增加较深土层的养分含量,打破了常年旋耕导致的营养物质在表层的富集,使土壤养分垂直分布更加均匀。
3.2 耕层重构对棉田土壤微生物数量的影响
土壤微生物在土壤中参与有机质和各种养分的分解转化,是评价土壤肥力的重要指标之一,其数量受土壤环境如土壤耕作措施的影响[22-24]。研究发现,适当的耕作措施如深松和翻耕能够降低土壤容重,增加土壤孔隙度和含水量,促进植株根系生长,有利于增加土壤气体交换,从而增加土壤微生物总体含量,提高深层土壤微生物数量,改善土壤微生态环境[25-27]。本研究中,在引入修正系数消除土层互换引起的本底差异时,由于没有测定土壤重构处理后到棉花播种前土壤的微生物的数量,故用棉花苗期的微生物数量计算修正系数。结果表明,与旋耕相比,耕层重构处理后0~20 cm土层微生物数量降低,而20 cm以下土层微生物数量增加,原因可能是土壤的微生物主要集中在0~20 cm土层,耕层重构将0~20 cm土层和20~40 cm土层互换,减少了表层土微生物的集聚,并将微生物数量较多的0~20 cm土层翻转下去,增加了深层土的微生物数量;另外耕层重构改善了耕层下部土壤物理结构,降低下部土壤容重[4],增加透气性和储水量[28],增加耕层下部土壤有机质等养分的积累,促进棉花根系下扎[7],深层根系分泌物和脱落物的增加为微生物繁殖提供了有机质等养分,利于微生物繁殖,从而影响20 cm以下土层的土壤微生物数量。土壤中真菌数量与植物病害发生密切相关,前期研究表明耕层重构降低了棉花的黄萎病病情指数[7],本研究中耕层重构显著降低了0~20 cm土层的真菌数量,可能会减轻棉花病害的发生。
3.3 耕层重构对棉田土壤酶活性的影响
酶是土壤新陈代谢的重要因素,参与土壤的许多重要的生物化学和物质循环过程,与土壤理化特性、肥力状况和农业措施有着显著的相关性,也是评价土壤肥力的重要指标之一。土壤酶的活性易受耕作措施影响,原因可能是耕作措施改变了土壤的理化性质和微生物的群落结构等。胡心意等认为,与浅耕相比,深耕可提高耕层土壤磷酸酶和β-葡萄糖苷酶的活性[29]。前人研究表明,不同耕作方式下相关土壤酶活性表现为翻耕>旋耕>免耕[30]。虽然不同耕作方式对土壤酶活性的影响不同,但是土壤酶活性都表现为从上层土壤到下层土壤逐渐降低[31]。本研究表明,与旋耕对照相比,耕层重构后棉田土壤0~20 cm土层的脲酶活性、蔗糖酶活性和碱性磷酸酶活性降低,而20 cm以下土层的土壤酶活性升高。其原因可能是耕层重构后转深土层的养分含量提高,为土壤酶提供丰富的底物;另外耕层重构能增加深层土壤的微生物数量,促进植株根系下扎[7],增加土壤酶的来源。耕层重构处理后20~40 cm土层与0~20 cm土层的土壤微生物数量和土壤酶活性差异不显著或显著升高,说明耕层重构技术打破了土壤养分、微生物和酶在土表层的富集,可改善土壤的生态环境。而耕层重构改善土壤生态环境的持续时间,需要进一步研究。
3.4 土壤微生物数量、土壤酶活性和土壤养分的相关性分析
前人研究表明,土壤微生物、酶与相关土壤肥力指标有着密切的关系,土壤微生物和土壤酶对外界环境的变化更为敏感,可以将其与土壤养分相结合对土壤肥力进行评估。王灿等认为,土壤中脲酶活性、转化酶活性和碱性磷酸酶活性之间呈极显著正相关,与土壤全磷含量、硝态氮含量和速效磷含量也存在相关性[32]。也有相关研究表明,土壤微生物数量与土壤有机质、有效氮、有效磷、有效钾含量呈正相关,与土壤pH呈负相关[33-34]。刘艳等认为,土壤脲酶、过氧化氢酶活性和土壤有机质及多种养分含量显著正相关,过氧化物酶活性和土壤有机质及多种养分含量显著负相关[35]。本研究中相关性分析结果表明,土壤微生物、土壤酶与土壤肥力相关指标关系密切;进一步回归分析结果表明,真菌数量、土壤脲酶活性和碱性磷酸酶活性与土壤肥力相关指标呈显著线性关系,是反映土壤肥力的重要指标。土壤微生物数量增多,土壤酶活性增强,有利于全氮、全磷、有机质、碱解氮、速效磷和速效钾等含量的提高,对养分的分解和转化具有积极作用;另外,养分也可通过改变微生物的生存环境调节微生物的数量和酶活性。
3.5 耕层重构对棉花产量和地上部生物量的影响
前期研究发现,耕层重构处理打破犁底层,使0~60 cm土层土壤容重显著降低,便于棉花根系下扎与土壤水分下渗,在灌水与大雨后使水分更多地蓄积在下层土壤中,达到蓄水保墒的目的,可促进棉花生长;耕层重构处理在干旱年份可以提高棉花产量,而在多雨年份增产效果不明显[7]。在本研究中,耕层重构处理对籽棉产量影响不显著,但可以显著提高棉花地上部生物量。这可能是因为耕层重构处理后棉田深层土壤养分含量增加,微生物数量和土壤酶活性提高,养分代谢增强,更利于棉花对营养的吸收和利用,促进棉花生长发育。
4 结论
与旋耕相比,耕层重构处理增加了棉田20~80 cm土层的养分含量,打破了由于常年旋耕导致的营养物质在表层的富集,使土壤养分垂直分布更加均匀。耕层重构处理导致表层土壤微生物数量和土壤酶活性降低,而使20~40 cm土层微生物数量和土壤酶活性增加,可提高土壤养分代谢强度;耕层重构处理可以提高棉花地上部生物量。相关分析和回归分析表明,土壤脲酶活性、碱性磷酸酶活性和真菌数量是反映土壤肥力的重要指标。因此,耕层重构可以提高20~40 cm土层的养分含量、微生物数量和土壤酶活性,提高较深土层的养分代谢强度,是改良土壤微生态环境的有效措施。
猜你喜欢
东南大学学报(自然科学版)(2022年4期)2022-08-12
建材发展导向(2022年10期)2022-07-28
土壤学报(2022年1期)2022-03-08
吉林蔬菜(2021年2期)2021-07-19
安徽农业科学(2020年23期)2020-12-28
农民致富之友(2020年10期)2020-05-11
农民致富之友(2020年4期)2020-04-03
当代陕西(2020年24期)2020-02-01
当代陕西(2020年24期)2020-02-01
当代陕西(2020年24期)2020-02-01
