
毛竹不同扩张阶段林分的结构特征1)
2022-01-06 11:42王秀云宋绪忠杨华吴柏林林晓越
东北林业大学学报 2021年11期
王秀云 宋绪忠 杨华 吴柏林 林晓越
(浙江省林业科学研究院,杭州,310023) (龙游县林业水利局)
毛竹(Phyllostachysedulis)属禾本科竹亚科刚竹属,是一种高大散生状乔木型植物,主要分布在我国亚热带地区[1],其株高可达20 m,胸径可达20 cm,是我国南方重要的笋材两用竹种,对增加林农收入、促进地方经济发展起到了积极作用。毛竹依靠竹鞭快速的繁殖、强大的适应改造和空间拓展能力,不断向邻近森林群落扩张、发笋成竹,形成竹阔混交林并不断纯林化[2]。毛竹林不断无序扩张,改变群落结构和组成、降低动植物生物多样性、抑制群落自然更新、减弱生态系统功能、破坏森林土壤理化性质与微生物组成等[3-4],给邻近的森林群落构成了巨大威胁,引起政府、生态学与林学界学者的关注。
现有研究认为,毛竹能向邻近森林群落扩张,既受自身生物属性影响,也与邻近森林生物与非生物因素(可侵入性)有关[5]。对毛竹扩张的研究多集中在对邻近森林群落生物多样性、土壤理化性质产生的影响研究[6-7],对毛竹不同扩张阶段林分结构变化研究鲜有报道。由物种组成、径阶分布、生物多样性、混交、竞争、种群分布格局构成的林分结构是研究森林经营学的重要基础[8-10],也是评价森林健康状况、森林群落功能复杂性与稳定性的重要指标,通常用于衡量森林抗干扰和自我修复能力[11-14]。
本文采用时空替代法,在浙江龙游县溪口国有林场选择典型的毛竹林向常绿阔叶林扩张的样地,通过群落特征调查,比较分析毛竹扩张前的常绿阔叶林、扩张中形成的竹阔混交林和扩张后形成的毛竹纯林三者的乔木层物种组成、径阶结构和物种多样性等非空间结构和空间结构特征,以期为客观评价毛竹扩张对阔叶林的生态后效提供参考,对提前防控毛竹扩张对邻近阔叶林产生的影响,及恢复与管理受侵森林群落或生态系统均具有重要意义。
1 研究区概况
研究区设在浙江省龙游县溪口国有林场,位于119°3′22″~119°8′30″E,28°44′12″~28°54′3″N。年平均气温为17.3 ℃,最冷月平均气温为5.1 ℃,最热月平均气温29.2 ℃,≥10 ℃积温2 500~5 100 ℃,无霜期209~310 d;年降水量1 542 mm,年相对湿度79%。属中亚热带季风气候,四季分明,气候温暖,雨量充沛,光照适宜。土壤主要有黄壤和红壤2个土类:黄壤一般在海拔600 m以上,土层厚度一般50~60 cm,肥力一般;红壤主要分布在海拔600 m以下,土层厚度一般在40~80 cm。其森林植被属于中亚热带常绿阔叶林,垂直带为浙闽山丘甜槠(Castanopsiseyrei)、木荷(Schimasuperba)植被区,主要森林类型有毛竹林、针阔混交林、常绿阔叶林和杉木(Cunninghamialaceolata)林等,优势树种主要有青冈(Cyclobalanopsisglauca)、苦槠(Castanopsissclerophylla)等壳斗科(Fagaceae)树种、木荷、马尾松(Pinusmassoniana)、杉木、毛竹等;人工林主要有杉木、马尾松。林地的立地条件较好,土层深厚,土壤肥沃,植被生长期长,适宜毛竹生长。
龙游县的毛竹林分布广泛,其面积达2.75万hm2,占森林面积的43.1%,而且竹产业经济效益高,带动了龙游县经济发展和居民生活水平提高,但近年来,由于竹产业效益下滑,林农放弃对毛竹林的经营,毛竹林自然扩张明显,邻近常绿阔叶林受到入侵威胁致群落树种组成、生物多样性、土壤菌根系统受扰等,并且形成了大量竹阔混交林和毛竹纯林,因此选择该区开展毛竹林自然扩张干扰对其邻近常绿阔叶林林分结构的影响研究非常具有代表性。
2 研究方法
2.1 样地设置与调查
2018年4月,在溪口国有林场典型选取毛竹林向常绿阔叶林扩张的3个界面,每个界面沿毛竹林扩张方向,设置扩张前常绿阔叶林、扩张中形成的竹阔混交林和扩张后形成的毛竹林样地各3块(表1),样地规格为30 m×30 m,记录每个样地的坐标、海拔、坡度、坡向、坡位,测量郁闭度。将每个样地划分为5 m×5 m的大样方,将样方内胸径大于5 cm的所有树木(包括毛竹)编号、挂牌,记录树种名称,并进行每木检尺,测量胸径、树高、冠幅、枝下高、坐标等指标。
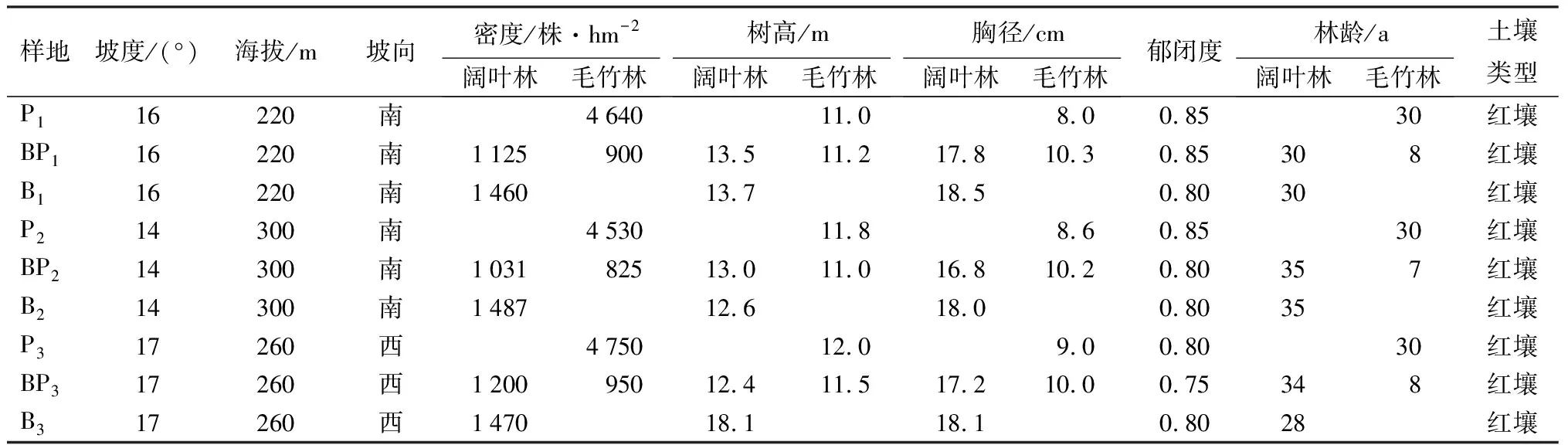
表1 研究样地基本概况
样地海拔高度为100~300 m,阔叶林主要为次生常绿阔叶林,林龄30~35 a,乔木层优势种主要为甜槠,伴生树种主要包括锥栗、青冈、木荷和冬青等。乔木层高度12.9 m,平均胸径18.0 cm,郁闭度0.80,株密度1 472株·hm-2。竹阔混交林为毛竹向阔叶林扩张6~8 a后形成,竹阔数量比接近4∶5,其中毛竹高度11.0 m,平均胸径10.0 cm,密度为890株·hm-2。毛竹林主要是30 a前自然扩张形成,留有少量阔叶树种,其中毛竹密度为4 650株·hm-2,平均胸径8.0 cm,平均高度11.0 m。
2.2 林分结构特征指标测定
林分结构特征包括林分非空间结构特征与空间结构特征。林分非空间结构特征包括种类组成、径阶分布、树高分布和物种多样性等。
采用上限排外法与林分实际情况相结合,以4 cm为一个径阶,分别统计毛竹3个扩张阶段林分林木在不同径阶的数量占比;以2 m为一个高度级,分别统计毛竹3个扩张阶段林分林木在不同高度级的数量占比。
多样性指数以相对重要值为测算依据,采用物种丰富度指数、物种多样性指数和均匀度指数来评价物种多样性。物种丰富度(R)等于物种总数;物种丰富度Margalef指数(Dm),即Dm=(S-1)/lnN;物种多样性采用Shannon-wiener指数(H′),即H′=-∑PilnPi;均匀度采用Pielou指数(Jsw),即Jsw=H′/lnS。式中:S为每一块样地中的物种总数,N为所有物种的个体数之和;Pi为物种i的相对重要值。乔木层的重要值=(相对多度+相对显著度+相对频度)/3。
林分空间结构主要采用描述树种空间隔离程度的树种混交度(Mi)、体现林木个体大小分化差异的大小比数(Ui)和反映林木个体水平分布格局的角尺度(Wi)3个参数对毛竹不同扩张阶段林分的空间结构分析[15-17]。本研究选定相邻木4株,标准角α0为72°。计算公式如下:

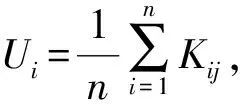
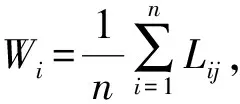
2.3 数据处理
采用单因素方差分析和最小显著差异法比较各多样性指数在毛竹林、竹阔混交林和常绿阔叶林间各结构指标的差异显著性。通过SPSS21.0进行数据分析和绘图。使用Winkelmass软件,对9块30 m×30 m的样地林分林分空间结构进行计算分析。
3 结果与分析
3.1 树种组成
毛竹不同扩张阶段,林分内毛竹立竹度、阔叶树种组成差别较大,扩张前期、中期和后期林分内毛竹立竹度依次为0、890和4650株·hm-2,扩张中期较前期、后期较中期其毛竹立竹度分别增加了890、3 760株·hm-2;相反,阔叶树种密度分别是1 472、1 082、120株·hm-2。说明毛竹扩张过程中,毛竹立竹度显著增加,毛竹扩张改变了群落优势树种组成。
由表2可知,扩张前期、中期和后期物种组成和数量特征均差异明显。对于乔木层来说,扩张前期的常绿阔叶林中甜槠占优势,重要值为62.59,伴生树种主要包括锥栗、青冈、木荷和冬青等;扩张中期形成的竹阔混交林,优势种甜槠的重要值降为46.21,伴生种毛竹、锥栗的重要值分别是26.17和6.68;扩张后期林分的优势种是毛竹,其重要值高达92.04,而甜槠和锥栗的重要值分别降到了3.17和1.58。说明毛竹不同扩张阶段,林分物种组成及物种在群落中的地位存在明显不同。
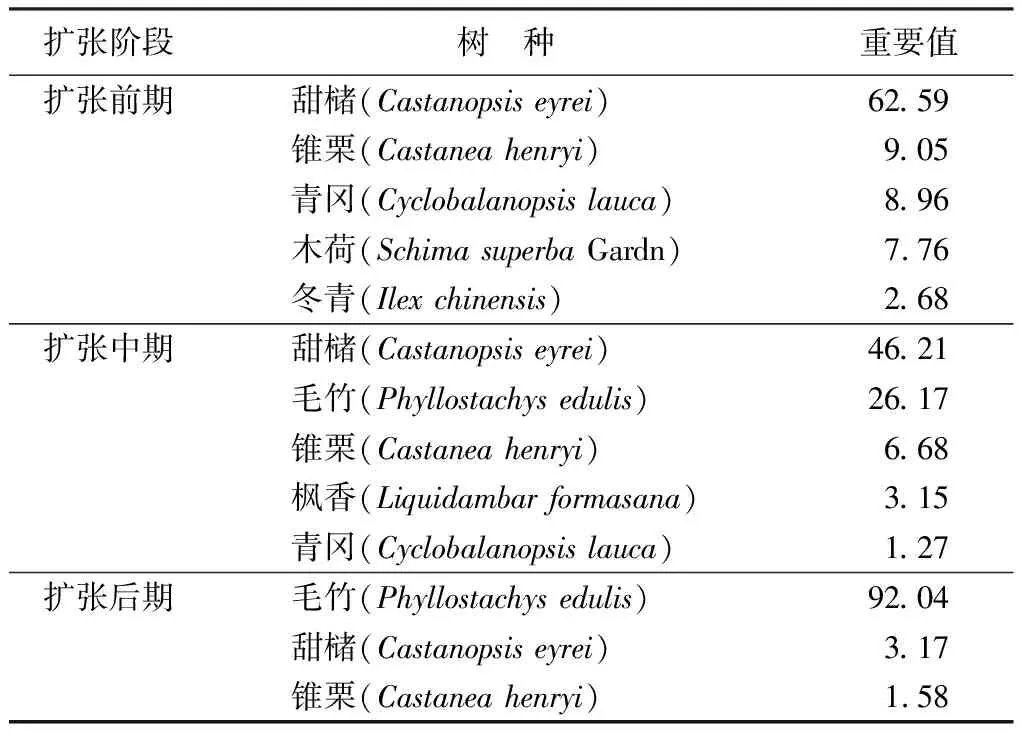
表2 不同扩张阶段林分的树种组成
3.2 乔木层径阶和垂直分布的变化特征
3.2.1 径阶分布的变化特征
表3可知,毛竹林不同扩张阶段,林分乔木层径阶结构变化明显,乔木在各径阶的数量分布不同。扩张前期阔叶林径阶分布范围最广,从4径阶一直到48径阶均有分布,而且呈倒“J”型曲线,即林分直径分布随着径级的增大,数量随之减少,其中8、12、16、20和24径阶的数量占比较高,依次为21.54%、18.46%、16.92%、13.85%和7.69%,总和为78.46%。到扩张中期,阔叶林演变为竹阔混交林,径阶分布范围没有变化,但分布由倒“J”型变为单峰山状曲线,且主要集中在8、12、16和20径阶,分别是28.32%、34.51%、10.62%和9.73%,总和为83.18%,其中较大径阶(28~48 cm)数量占比比阔叶林减少了42.5%;扩张后期,毛竹林的径阶结构明显变窄,大径阶个体数量明显减少,主要集中于8和12径阶,多度百分比分别是37.5%和62.5%。

表3 不同扩张阶段林分径阶结构
3.2.2 垂直分布的变化特征
从表4可发现,毛竹林扩张前期、扩张中期到扩张后期,林分乔木层垂直结构分层现象减弱。扩张前期阔叶林分垂直分布范围最广,从4~18 m高度级均有分布,其中4、6、8、10和14 m高度级的数量占比较高,依次为27.69%、15.38%、13.85%、15.38%和12.31%,总和为84.62%;扩张中期林分垂直分布比较分散,呈多峰山状曲线,4、6、8、10、12和14 m高度级的数量占比较高,分别是17.70%、11.50%、21.24%、16.81%、12.39%和15.93%,平均占比15.93%,林分垂直分布结构不稳定;扩张后期毛竹林的垂直分布结构明显变窄,且缺失16和18 m的较大高度级个体分布,主要集中于8、10、12和14 m高度级,多度百分比分别是31.25%、18.75%、18.75%和20.83%,总和为89.58%。
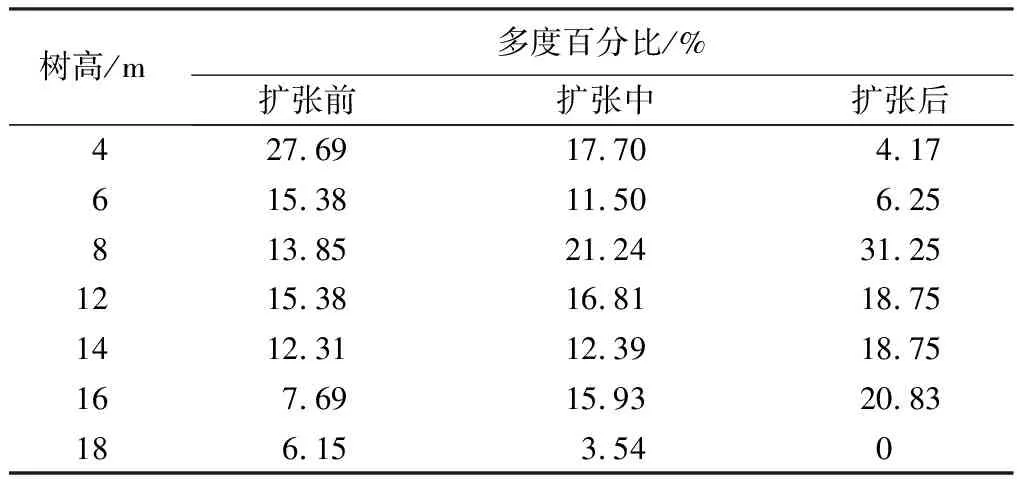
表4 不同扩张阶段林分垂直结构
3.3 物种多样性的变化特征
从表5可知,毛竹扩张前期、中期和后期林分乔木层的物种多样性指数差异显著(P<0.05),4个多样性指数均表现为扩张前期最高,扩张中期居中,扩张后期最低。其中Margalef指数、Shannon-Wiener指数和Pielou均匀度3个多样性指数分别从扩张前期的3.92、3.09和0.96降至扩张后期的0.14、0.08和0.03,反映出毛竹林扩张后,受干扰的群落物种种类明显减少,而且物种分布均匀度下降,其生物多样性变化明显。

表5 不同扩张阶段林分乔木层物种多样性
3.4 空间结构
3.4.1 树种混交情况
毛竹不同扩张阶段样地混交度频率分布见表6,平均值见表7(不同小写字母表示不同林分混交度差异显著,P<0.0.5)。扩张前期,阔叶林以强度混交和极强度混交为主,其二者林木占比为59.3%。扩张中期形成的竹阔混交林混交度分布比较均匀,混交度平均值由扩张前期的0.663降到0.540,说明大多数林木周围只有1~2株不同种的林木,树种混交程度不高。扩张后期形成毛竹林,样地内毛竹占主导地位,仅存在少量阔叶树,零度混交的林木占比为92.3%。而且经单因素方差分析和多重比较(LSD)检验,毛竹扩张3个阶段的林分平均混交度存在显著性差异(P<0.05),说明毛竹扩张不仅改变了阔叶林的树种多样性,而且也改变了林分组成。
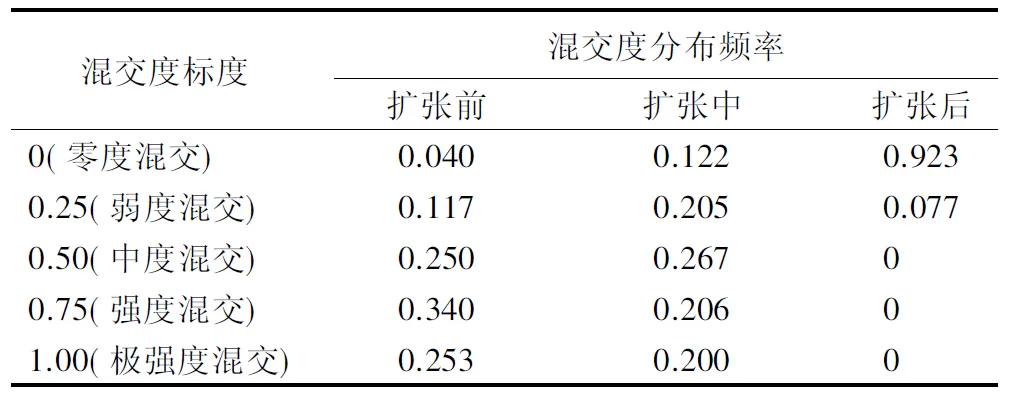
表6 不同扩张阶段林分混交度分布频率
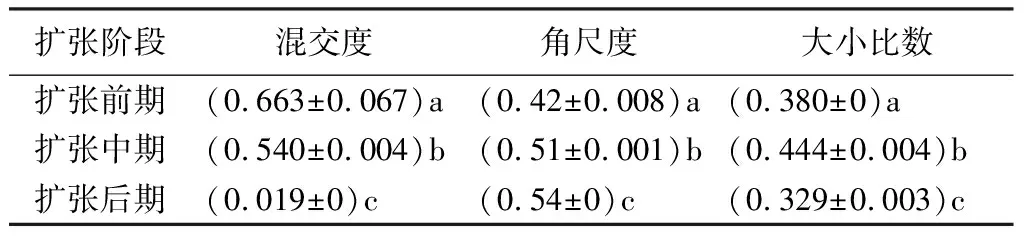
表7 毛竹不同扩张阶段林分空间结构特征
3.4.2 林木空间分布格局
不同扩张阶段林分的角尺度参数分布见表8。由表8可知,毛竹扩张前期的阔叶林中,角尺度Wi=0.25和Wi=0.50的频率较高,分别为0.33和0.47,说明存在较多的均匀分布和随机分布,且不存在聚集分布。扩张中期,Wi=0.50的频率最高,为0.53,而Wi=0.25和Wi=0.75的频率较小,分别是0.19和0.27,说明存在较多的随机分布和少量的均匀分布,且不存在绝对均匀分布的林木。扩张后期的毛竹林,Wi=0.50和Wi=0.75的频率较高,分别是0.57和0.26,说明存在较多的随机分布和聚集分布;Wi=0.25的频率较低,为0.12,且不存在绝对均匀分布的林木。不同扩张阶段林分平均角尺度(W)见表7。扩张前期林分的平均角尺度W=0.42,林木空间分布格局为均匀分布;扩张中期的W=0.51,为随机分布;而扩张后期的W=0.54,为聚集分布,但聚集程度较低。

表8 不同扩张阶段林分角尺度分布频率
3.4.3 林木大小分化情况
大小比数是用以描述林木个体大小的分化程度的结构指标。扩张前期、中期和后期的林分大小比数频率分布如表9所示。扩张前期,林分内整体处于优势和亚优势生长状态的林木占比较大为0.611,处于劣势和极劣势生长状态的受压木较少为0.204,中庸木占比0.186,整体处于亚优势的生长状态。扩张中期形成的竹阔混交林大小比数频率分布比较均匀,胸径大小差异明显,但处于中庸生长状态的林木占比最高为0.333,同时存在较大比例(0.281)的劣势和极劣势生长状态的林木,整体亚优势接近中庸的生长状态,林分没有形成绝对优势的种群,林分结构不稳定。扩张后期,林分处于优势和亚优势的林木达0.582,而且不存在处于极劣势生态状态的受压木,林分平均大小比数为0.329,处于亚优势生长状态。而且通过单因素方差分析三者林分林木大小比数的频率分布差异显著(P>0.05),说明毛竹扩张明显改变了林木生长竞争态势。
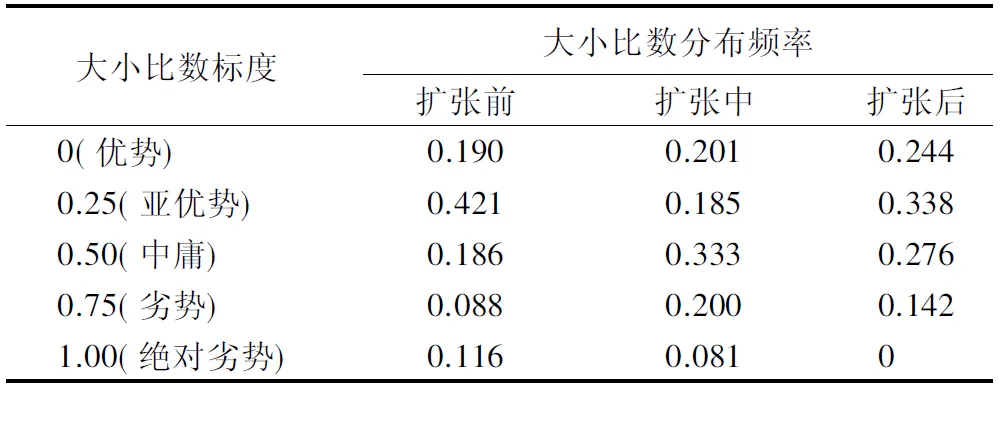
表9 不同扩张阶段林分大小比数分布频率
4 结论与讨论
研究发现,毛竹扩张干扰不仅改变了阔叶林的树种组成,而且影响了树种在群落中的地位,毛竹扩张前期、中期和后期毛竹立竹度从0,依次增加到890和4 650株·hm-2,阔叶林的优势树种甜槠的重要值从扩张前期的62.59,扩张中期的46.21,直至降到扩张后期的7.94。这与许多研究结果相符。如欧阳明等[18]研究发现受毛竹扩张影响,阔叶林中建群种红楠的重要值由扩张前的30.22最终减少为0;白尚斌等[3]通过对毛竹扩张的动态定位监测研究得出,毛竹扩张过程中毛竹数量逐渐增加,而其它树种种类与数量明显减少。物种组成是群落结构的基础,物种数量减少会导致群落结构不稳定,毛竹向阔叶林扩张,最终入侵占领成为优势种,不仅改变了其群落结构,而且会对阔叶林的生态功能产生严重影响。
竹子扩张会影响邻近群落结构[5]。林分径阶和垂直结构是影响群落种间竞争、森林生态系统物质循环、能量流动和信息传递的关键。本研究发现,毛竹扩张干扰对阔叶林的分层有明显影响,导致其径阶分布范围收窄,从干扰前的4~48径阶均有分布,到扩张后期的毛竹林径阶分布主要集中在4和8径阶,而且减少了大径阶林木个体数量;同时毛竹林自然扩张导致群落垂直分布范围变窄,从扩张前期的4~18 m高度级均有分布到扩张后期林木高度主要集中在8~14 m,而16和18 m高度级的个体数量为零。
生物多样性物种丰富度和分布均匀度的综合体现,对群落能否保持动态稳定与抵抗外界干扰有着至关重要的影响作用。本研究结果表明物种丰富度指数、Margalef指数、shannon-Wiener指数和Pielou均匀度4个多样性指数均表现为扩张前期阔叶林最高,扩张中期居中,扩张后期最低,且差异显著,反映出毛竹林扩张后,受干扰的群落物种种类明显减少,而且物种分布均匀度下降,其生物多样性下降明显。这一结果与杨淑贞等[19]、林倩倩等[6]、欧阳明等[18]的研究相符。
由分别反映不同树种配置、竞争和分布格局的混交度、大小比数和角尺度组成的林分空间结构决定了树木之间的竞争优势和空间生态位,在很大程度上影响着林分的稳定性、发展的可能性和经营空间的大小和生态功能的发挥[20]。本研究发现毛竹林扩张前期,阔叶林平均混交度是0.663,到扩张中期降为0.54,直至扩张后期的0.019。毛竹扩张前期阔叶林的大小比数为0.380,林木整体呈亚优势生长态势;扩张中期竹阔混交林大小比数0.444,整体亚优势接近中庸的生长状态,林分结构不稳定;扩张后期,林分平均大小比数为0.329,处于亚优势生长状态,说明毛竹扩张明显改变了林木生长竞争态势。从林分空间分布看,扩张前期阔叶林的平均角尺度W=0.42,林木空间分布格局为均匀分布;扩张中期竹阔混交林的W=0.51,为随机分布;而扩张后期毛竹林的W=0.54,为聚集分布,但聚集程度较低。表明毛竹扩张干扰改变了林木个体的空间分布,进而改变了林分内光照、水分、温度、营养物质等微环境的空间分布,并对群落的动态稳定和抗干扰能力产生影响。
毛竹扩张符合“内禀优势、资源机遇与干扰促进”的生物入侵机制[21-22],即毛竹依靠生长迅速、繁殖力强、形态多样、集团协同等内禀优势,通过遮阴、机械损伤、凋落物抑制、养分竞争与化感作用等竞争方式,排挤其他植物并不断向邻近森林群落入侵扩张,直至毛竹纯林化。因此,如何有效防控毛竹无序扩张是一个非常有挑战性的问题,蔡亮等[1]提出了挖沟、灌水的设想;Suzuki et al.[23]对地上成竹或竞争排斥阶段的扩张,主要通过加大采笋量与伐竹量加以抑制。但在劳动力成本日益高涨的情形下,这些方法难以推行。因此,为避免毛竹扩张对邻近阔叶林的影响,如何采取科学有效的经营措施目前仍然需要深入研究。
猜你喜欢
安徽农学通报(2020年17期)2020-10-09
绿色中国(2019年18期)2020-01-04
农家科技下旬刊(2017年8期)2017-11-13
中国林副特产(2017年5期)2017-10-16
科技创新导报(2016年21期)2016-12-17
贵州科学(2016年2期)2016-06-13
华南农业大学学报(2016年3期)2016-06-01
