
黄金枸骨叶片复绿期叶绿素代谢的调控特征1)
2022-01-06 11:42黄婧苏金周鹏张强张敏
东北林业大学学报 2021年11期
黄婧 苏金 周鹏 张强 张敏
(江苏省林业科学研究院,南京,211153) (南京林业大学) (江苏省林业科学研究院) (南京林业大学) (江苏省林业科学研究院)
近年来,彩叶树种因其独特的叶色在园林绿化中备受重视,彩叶树种成为绿化发展的新趋势,目前许多研究人员争相引进和培育各种彩叶植物。植物的色彩是由叶片内色素的种类、含量及其分布决定的[1-2],光照作为重要的环境因子影响植物色素合成及相关酶的活性,从而影响叶片呈色。遮荫可明显改变植株的光照强度,植物在遮荫环境下,形态学和生理学上都会表现出一定的适应性。
高等植物叶绿素合成是由许多酶参与的一个复杂过程[3],其中任何一个环节发生突变都可能影响叶绿素的合成,从而引起叶色的变化。首先是通过谷氨酸合成δ-氨基乙酰丙酸,再由δ-氨基乙酰丙酸转变为胆色素原;然后4分子的胆色素原聚合成为1分子线性的四吡咯分子羟甲基胆色烷,最后异构化形成环状的尿卟啉原Ⅲ,这是叶绿素各前体物质中最先具有卟啉环结构的分子。形成卟啉环骨架后,粪卟啉原Ⅲ会氧化成原卟啉Ⅸ,接着再形成镁-原卟啉Ⅸ和原叶绿素酸酯(最后一个前体物质)。在此过程中,δ-氨基乙酰丙酸(ALA)、胆色素原(PBG)、尿卟啉原III(Urogen III)、粪卟啉原III(Coprogen II)、原卟啉IX(ProtoIX)、镁-原卟啉(Mg-Proto)和原叶绿素酸酯(Pchlide)是其重要合成前体物[4]。谷氨酰-tRNA还原酶(GluTR)是δ-氨基乙酰丙酸合成的关键酶,催化δ-氨基乙酰丙酸向尿卟啉原Ⅲ的转化;而催化δ-氨基乙酰丙酸生成胆色素原的δ-氨基乙酰丙酸脱水酶(ALAD);以及催化羟甲基胆素重排环、合成天然四吡咯化合物的尿卟啉原III合酶(UmgenIIIs),是植物叶绿素合成途径的关键酶[5]。同时,高等植物体内合成的叶绿素可通过脱植基、脱镁等一系列反应而降解。在叶绿素降解过程中,催化叶绿素脱去植醇的叶绿素酶(Chlorophyllase)、脱镁螯合酶(MDCase)也是高等植物叶绿素降解过程的关键酶[6-7]。
彩叶冬青黄金枸骨(Ilexxattenuata‘Sunny Foster’)属于光照敏感型叶色突变体,是冬青品种选育中重要的种质资源。冬青叶色受光照强度影响,在全光或强光条件下,叶片呈现黄色,在弱光环境下叶片的色彩淡化,如建筑物的遮挡,园林植物相互间的遮挡,出现色彩变浅、叶片复绿的现象,降低观赏价值,这已成为园林施工中的难题,但目前关于彩叶树种叶片复绿研究较少。为了研究黄金构骨叶片受光照强度调控机制,通过人工遮荫的方法,使黄金枸骨叶片从黄色变为绿色,对此过程中叶绿素合成主要中间产物的含量和关键酶活性进行动态追踪,描绘叶绿素生物合成代谢特征,寻找叶片变色过程中叶绿素合成的关键节点,为黄金枸骨在园林绿化中的应用提供技术支持,也为冬青叶色改良提供新的思路。
1 材料与方法
1.1 实验材料
试验地点在江苏省南京市江宁区林业科学研究院温室大棚。试验时间为2018年9—10月份。选择生长良好且长势一致的1年生黄金枸骨苗木,移栽至塑料容器中,每盆1株,共30盆,进行常规管理。利用黑色遮荫网进行四面遮荫处理,经测定,遮荫前后光照强度分别为1 500、150 μmol·m-2·s-1。6个采样时间分别为:遮阴前(全黄)、遮阴6、15、22、29、36 d。每个时期进行3次重复试验,每个重复随机选取3株黄金枸骨进行各项指标测定。
1.2 各生理指标测定方法
叶绿素的质量摩尔浓度测定:叶绿素的质量摩尔浓度测定参照Aron[8]的方法测定。取0.1 g叶片,加入体积分数为80%丙酮2~3 mL,磨成匀浆,转入10 mL棕色容量瓶中,再加80%的丙酮水溶液定容至10 mL,避光浸提。将色素提取液倒入比色杯内。以80%丙酮为对照,在波长645、663和440 nm处测定吸光值,并计算叶片的叶绿素a、叶绿素b、总叶绿素的质量分数。
叶绿素前体物质的质量摩尔浓度测定:δ-氨基乙酰丙酸(ALA)的质量摩尔浓度的测定参照Dei[9]的方法。ALA的质量摩尔浓度以535 nm的摩尔消光系数7.2×104mol-1·cm-1计算。
胆色素原(PBG)的质量摩尔浓度的测定参照徐培洲等[10]的方法。胆色素原的质量摩尔浓度以535 nm的摩尔消光系数6.1×104mol-1·cm-1计算。
尿卟啉原Ⅲ(Urogen Ⅲ)和粪卟啉原Ⅲ(Coprogen Ⅲ)的质量摩尔浓度的测定参照Bogorad[10]的方法,Urogen Ⅲ的质量摩尔浓度以405.5 nm的摩尔消光系数5.48×105mol-1·cm-1计算,Coprogen Ⅲ的质量摩尔浓度以399.5 nm的摩尔消光系数4.89×105mol-1·cm-1计算。
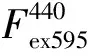
叶绿素代谢关键酶活性测定:谷氨酰-tRNA还原酶活性、δ-氨基乙酰丙酸脱水酶(ALAD)活性、尿卟啉原Ⅲ合酶(Uroporphyrinogen III synthase)活性、叶绿素酶和脱镁螯合酶活性检测,采用对应的ELISA试剂盒测定。
1.3 数据处理
对测定或统计的各指标采用SPSS15.0软件进行方差分析和Duncan’s多重比较,利用Excel进行数据运算。
2 结果与分析
2.1 叶绿素合成前体物质的质量摩尔浓度变化
由表1、表2可知,黄金构骨叶片变绿过程中,叶绿素a、叶绿素b和总叶绿素含量显著增加。叶绿素合成初期的前体物质δ-氨基乙酰丙酸和胆色素原的质量摩尔浓度显著减少(P<0.05)。δ-氨基乙酰丙酸的质量摩尔浓度从0.11 μmol·g-1减少到0.89 μmol·g-1,降幅为46.24%,其中0~15 d下降最显著。胆色素原的质量摩尔浓度从0.35 μmol·g-1减少到3.55 μmol·g-1,降幅为38.77%。
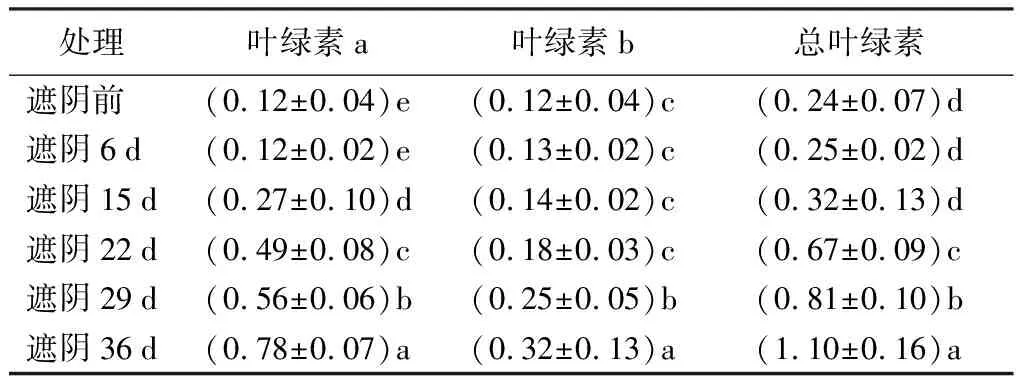
表1 叶绿素质量分数的变化 mg·g-1

表2 叶绿素合成前体物质的质量摩尔浓度 μmol·g-1
叶绿素合成过程中,位于胆色素原下游的前体物质尿卟啉原Ⅲ的质量摩尔浓度显著升高(P<0.05),从0.36 μmol·g-1升高到0.72 μmol·g-1,增幅为100.46%,遮阴第6 d后增幅显著。粪卟啉原Ⅲ的质量摩尔浓度也显著升高(P<0.05),从0.62 μmol·g-1升高到1.25 μmol·g-1,增幅为100.27%,遮阴第6、22、36 d增幅最显著。
尿卟啉原Ⅲ下游的原卟啉IX、镁-原卟啉和原叶绿素酸酯这3个前体物质的质量摩尔浓度都显著升高(P<0.05)。原卟啉IX的质量摩尔浓度从0.63 μmol·g-1升高到1.11 μmol·g-1,增幅为76.85%,镁-原卟啉的质量摩尔浓度从0.77 μmol·g-1升高到1.29 μmol·g-1,增幅为67.63%,原叶绿素酸酯的质量摩尔浓度从0.23 μmol·g-1升高到0.65 μmol·g-1,增幅达185.57%。
2.2 叶绿素代谢关键酶活性的变化
由表3可知,叶绿素合成关键酶谷氨酰-tRNA还原酶和δ-氨基乙酰丙酸脱水酶的活性随着叶片变绿显著下降(P<0.05),谷氨酰-tRNA还原酶活性从3 712.302 U/g下降到3 033.036 U/g,降幅为18.3%;δ-氨基乙酰丙酸脱水酶的活性从3 988.89 U/g下降到2 599.89 U/g,降幅为34.82%。而叶绿素合成关键酶尿卟啉原Ⅲ合酶的活性随着叶片变绿显著上升(P<0.05),从3 441.48 U/g上升到4 192.55 U/g,增幅为21.82%,从遮阴第6 d开始显著上升。
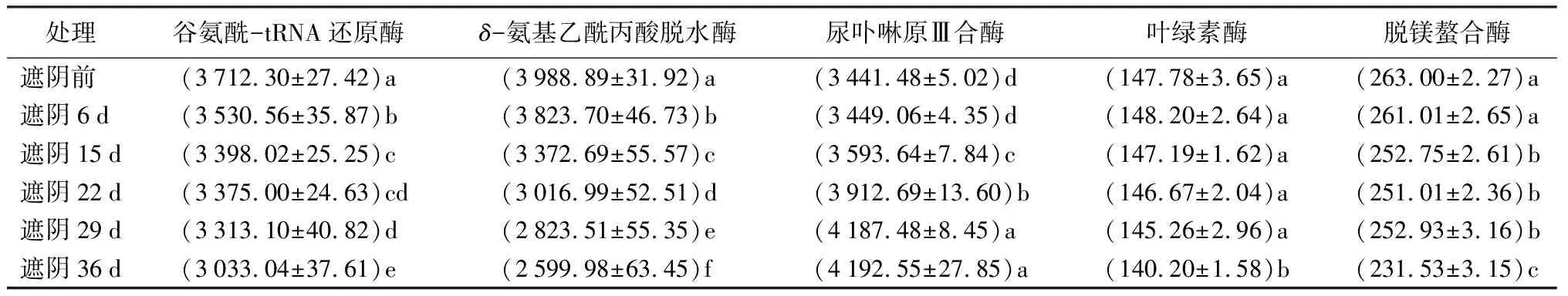
表3 叶绿素代谢关键酶活性 U·g-1
叶绿素降解关键酶叶绿素酶和脱镁螯合酶的活性都随着叶片变色而下降。叶绿素酶的活性从147.781 U/g下降到140.200 U/g,主要在29~36 d时显著下降(P<0.05),降幅为5.13%,其他时期无显著差异。脱镁螯合酶的活性随着叶片变绿显著下降(P<0.05),从263.003 U/g下降到231.531 U/g,降幅为11.97%,遮阴第6 d开始显著降低。
3 讨论
本文研究了黄金枸骨叶片复绿期的叶绿素代谢调控特征,发现随着叶色复绿,叶绿素合成关键酶活性升高,叶绿素合成前体物质含量显著增加,叶绿素的积累起始于尿卟啉原Ⅲ的增加,起始时间为遮荫后6~15 d;同时叶绿素降解酶活性降低,促进叶绿素的积累,最终实现了叶片在遮荫条件下叶绿素含量升高,叶片由黄转绿。
本研究对叶绿素合成通路中重要的前体物质的质量摩尔浓度分析发现,叶绿素合成通路上游的物质δ-氨基乙酰丙酸和胆色素原的质量摩尔浓度显著减少,而其后生成的尿卟啉原Ⅲ的质量摩尔浓度显著升高,并且尿卟啉原Ⅲ之后,前体物质的质量摩尔浓度都显著升高,说明遮荫引起的叶绿素积累可能起始于尿卟啉原Ⅲ的增加。
对叶绿素合成关键酶活性变化分析发现,谷氨酰-tRNA还原酶和δ-氨基乙酰丙酸脱水酶的活性随着叶片变绿显著下降,尿卟啉原Ⅲ合酶活性显著上升,导致尿卟啉原Ⅲ的质量摩尔浓度显著升高,是引起叶绿素积累的关键。δ-氨基乙酰丙酸合成的关键酶谷氨酰-tRNA还原酶(GluTR)活性显著降低,是由于GluTR编码基因的转录受光的调控,遮荫后导致GIuTR酶活性的降低,这与Ilag et al.[13]的研究结果一致。Ilag et al.[13]研究发现GluTR编码基因的转录受光的调控,遮荫后光照强度的降低导致GIuTR酶活性的降低,最终使δ-氨基乙酰丙酸的质量摩尔浓度降低,在其他植物遮荫处理时也发现类似现象[14]。催化δ-氨基乙酰丙酸转化为胆色素原的δ-氨基乙酰丙酸脱水酶活性显著降低,是由于其底物δ-氨基乙酰丙酸的质量摩尔浓度减少,从而引起胆色素原的质量摩尔浓度呈下降趋势。而尿卟啉原Ⅲ合酶活性显著上升,是引起叶绿素积累的关键步骤,也说明尿卟啉原Ⅲ合酶受底物的质量摩尔浓度的影响较小,与俞金健等[15]的研究结果一致。Wang et al.[16]研究发现紫薇叶绿素缺失突变体在低光照条件下,粪卟啉原Ⅲ的质量摩尔浓度显著增加,尿卟啉原Ⅲ合酶活性在6~15 d显著上升,与本研究结果一致。
对叶绿素降解相关酶活性变化研究发现,叶片变色过程中脱镁螯合酶活性降低导致合成的叶绿素停留在植物体内,促进叶绿素的积累。脱镁螯合酶在遮荫前期(6~15 d)活性就显著降低,叶绿素酶到遮荫末期(29~36 d)活性下降,说明这两个降解通路对光照的敏感度不一样。同样说明叶绿素的积累一是由于叶绿素合成中间途径酶活性增强,导致前体物质的积累,二是叶绿素降解酶活性的下降,降低叶绿素的降解。
通过动态追踪叶色复绿期的叶绿素合成前体物质的质量摩尔浓度和代谢酶活性发现,叶绿素积累的关键物质——尿卟啉原Ⅲ的质量摩尔浓度显著增加,叶绿素积累的关键酶——尿卟啉原Ⅲ合酶活性的显著升高,叶绿素合成和积累的关键时期——遮荫后6~15 d,此时叶绿素合成相关酶活性开始显著升高,叶绿素降解酶活性显著降低,叶绿素的质量分数也显著增加。本研究结果初步分析了黄金枸骨叶绿素合成代谢受光照调控的特征,为冬青叶色改良提供新的思路,也为其栽培生产和园林绿化应用提供技术支持。
猜你喜欢
分子催化(2022年1期)2022-11-02
河北画报(2020年10期)2020-11-26
英美文学研究论丛(2018年2期)2018-08-27
现代食品(2016年24期)2016-04-28
云南中医学院学报(2015年2期)2015-07-31
医学研究杂志(2015年5期)2015-06-10
化学反应工程与工艺(2015年1期)2015-04-16
中国塑料(2014年4期)2014-10-17
应用化工(2014年3期)2014-08-16
雕塑(2000年4期)2000-06-24
