
超级增强子调控基因表达的生物学作用及其在哺乳动物中的应用前景
2021-12-31 01:38:10康晓龙
畜牧兽医学报 2021年12期
赵 磊,康晓龙
(宁夏大学农学院,银川 750021)
增强子(enhancer)最早发现于家兔β-珠蛋白(β-globin)的转录研究中[1],之后在小鼠[2]等物种的基因组中得以证实。随着研究的进一步深入,人们发现增强子作为一种顺式作用的DNA调控元件,能远距离、无方向性地增加靶基因的转录输出,从而影响细胞的发育分化[3-7]。随着后基因组时代的到来,人类对于基因组和增强子的认识日益加深,在此基础上Whyte等[8]于2013年首先提出了超级增强子的概念。此后,关于超级增强子的多项研究均表明了超级增强子在基因表达调控中的重要性[9],因此,本文通过归纳超级增强子调控基因表达的生物学作用与主要应用,以期为后续开展超级增强子在动物重要表型调控中的分子作用研究提供参考与思路。
1 超级增强子及其特性
超级增强子(super enhancer,SE)的提出是基于对增强子的研究。2013 年,研究人员发现胚胎干细胞(embryonic stem cells,ESC)的主要转录因子在大多数控制多能性的基因上形成了不寻常的增强子域,将其定义为超级增强子[8]。其相对于普通增强子的特性主要包括以下几点(图1):1)超级增强子的转录因子结合程度显著高于普通增强子,其相关基因的表达水平更高;2)超级增强子相对于普通增强子DNaseI超敏反应增加;3)超级增强子具有高密度的组蛋白H3K4me1和H3K27ac修饰,为高密度的Mediator、p300 等辅助因子所占据;4)一些转录因子水平的降低对超级增强子相关基因表达的影响大于普通增强子[10-11];5)超级增强子自身能转录出大量的增强子RNA,且拥有更大密度的RNA聚合酶Ⅱ[12-13]。与普通的增强子相比,超级增强子内的单个组分增强子具有提高转录激活水平的作用[8],同时有证据表明,超级增强子内的组分增强子之间存在加性或协同作用,在基因调控中具有非冗余功能,而组分增强子的缺失可能会损害其他超级增强子组分的活性,最终导致整个超级增强子功能障碍[14-17]。随着超级增强子的发现,另一种能够维持细胞身份(cell identity)类型的转录增强子也被发现。Parker等[18]在试验中发现一种大小在3 kb以上且具有增强子特征的较大基因组区域,将其定义为延伸增强子(stretch enhancer)。超级增强子和延伸增强子都代表一类转录增强子,均被证明可以控制细胞身份基因的表达,并携带与疾病特质相关的变异。不同的是,超级增强子是根据主转录因子、染色质调节因子或染色质标记的结合占位而定义的增强子簇,比后者具有更高的转录活性和细胞类型特异性[19]。
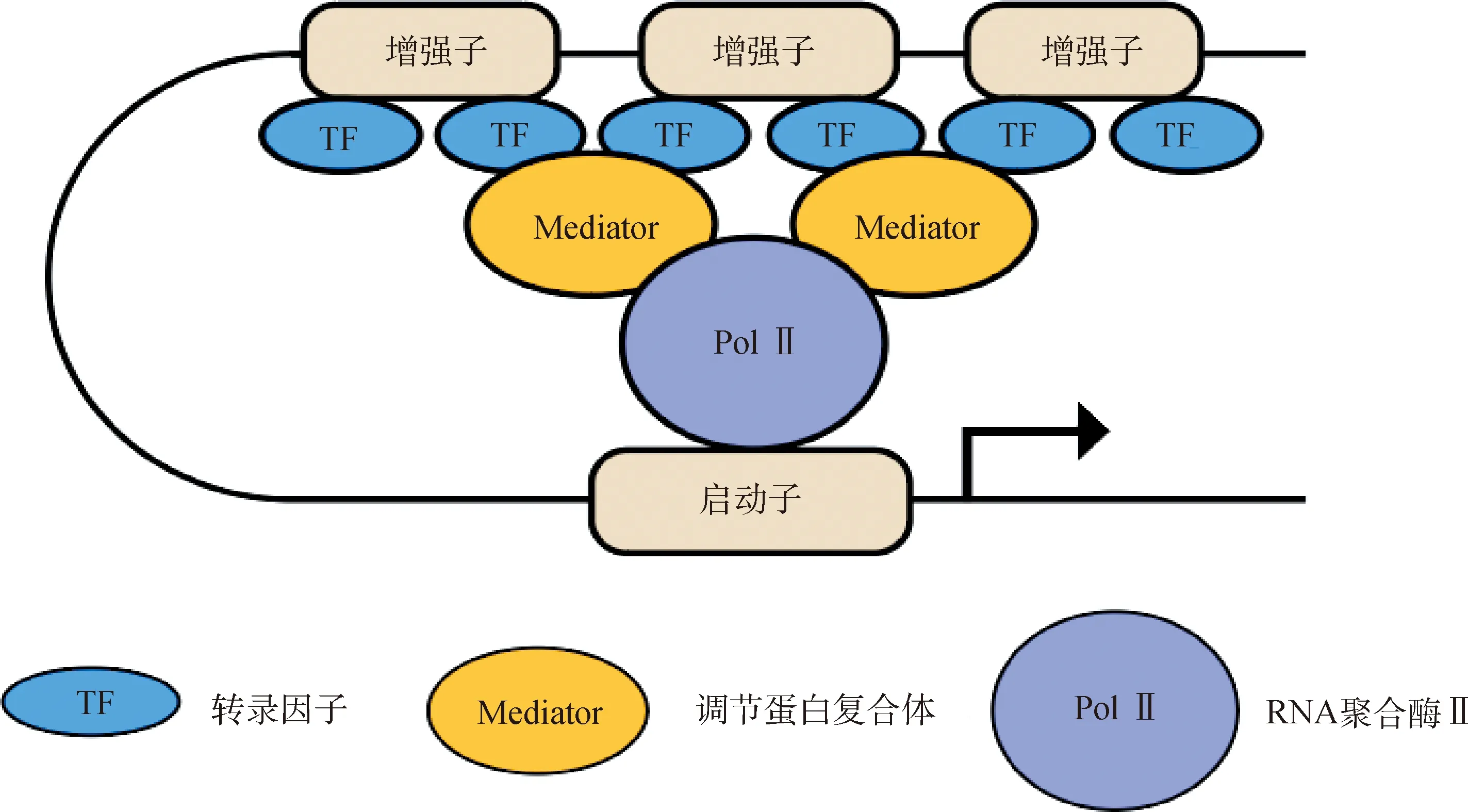
图1 超级增强子的结构Fig.1 The structure of the super enhancers
2 超级增强子的鉴定
超级增强子的鉴定是依据增强子转录活性标记分子结合水平强度的差异,这些分子包括辅因子(如Mediator和Cohesin)、组蛋白修饰标记(如 H3K27ac和H3K4me1)、染色质修饰分子(如p300)及BET家族蛋白(BRD4)[8,10]。当前超级增强子的鉴定主要通过染色质免疫沉淀测序(Chromatin immunoprecipitation followed by high-throughput sequencing,ChIP-seq),主要包括3个步骤(图2A):1)识别活性增强子位点;2)缝合增强子;3)确定超级增强子与普通增强子之间的阈值[20]。首先,利用ChIP-seq检测与活性增强子相关的因子或组蛋白修饰,如转录因子、转录共激活因子(Mediator、p300等)以及组蛋白修饰(H3K27ac、H3K4me1);其次,将所得增强子进行缝合。研究人员将相互连接于12.5 kb范围内的增强子定义为一个单一实体,即缝合增强子(stitched enhancer);最后,通过计算不同增强子区域Med1的ChIP-seq信号水平[21],对缝合增强子和其余的单个增强子进行排序,以获得结果曲线并确定阈值(图2B),以此区分超级增强子与普通增强子。近年来,研究者通过各种方法进行超级增强子的鉴定,如染色体构象捕获技术3C(chromosome conformation capture)[22]及其拓展技术4C(circular chromosome conformation capture)[23]、5C(chromosome conformation capture carbon copy)[24]以及Hi-C(High-through chromosome conformation capture)[25]等。
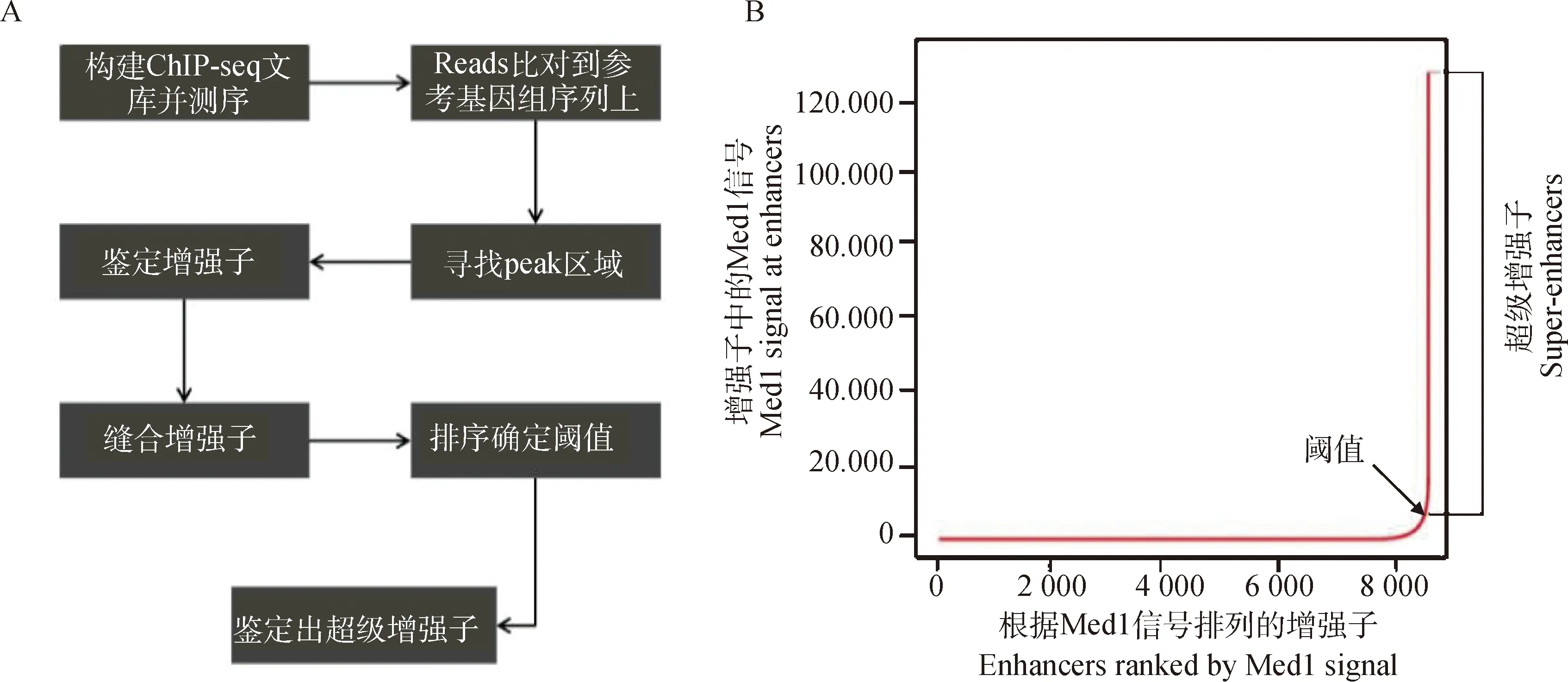
A.超级增强子的鉴定流程图;B.根据ChIP-seq 富集信号强度对增强子进行排序,确定阈值从而鉴定出超级增强子A.Flow chart for identification of super enhancers;B.Ranking of enhancers based on ChIP-seq enrichment signal strength and determination of thresholds to identify super enhancers图2 超级增强子的鉴定Fig.2 Identification of super enhancers
3 超级增强子的作用方式
3.1 超级增强子的基本调控机制
在真核生物中,增强子-启动子的相互作用是基因转录的重要环节。超级增强子作为基因转录过程中的正调节因子,在基因转录过程中扮演着重要角色,对基因的转录调控作用紧密依赖于其靶向转录因子、辅助因子和介质的募集,以及增强子-启动子相互作用的形成[26],这是超级增强子作为细胞中的一种三维结构体系调控启动子的作用模型,也与以往的多数研究结果相符。超级增强子作为一个大的增强子簇,它所包含的每个组分增强子,均可独立结合转录因子、辅助因子,共同调控同一启动子的转录活性,因此超级增强子的基本调控机制也依赖于其组分增强子的功能,最终通过增强子和启动子间的互作发挥功能。大量研究发现,增强子与启动子之间的相互作用是随着基因表达而建立的,但无法理清增强子与启动子的接触是基因激活的原因还是结果。对此有研究表明,增强子和启动子之间的物理接触传递了转录过程所需的相关调控信息[27];另有研究结果表明,诱导小鼠β-珠蛋白(β-globin)启动子与其位点控制区域增强子接触,会导致Hbb基因极显著的转录激活,即使在缺乏关键转录因子GATA1的情况下也是如此,直接证明增强子-启动子互作可以诱导基因转录[28]。在增强子和启动子互作过程中,拓扑关联结构域(topologically assocaited domain,TAD)起到重要作用。TAD是一种首先发现于哺乳动物细胞中的染色质结构单元[29],是真核生物基因组的基本成分[30]。多种证据表明,TAD的边界能起到绝缘体的作用,防止不适当的增强子-启动子相互作用。对此,相关研究能够证明:结构蛋白为TAD构建染色质并充当边界[31-32],而超级增强子边缘又广泛检测到结构蛋白[33],由此确定CTCF(CCCTC binding factor,在多种病毒中调控转录的蛋白)是TAD和超级增强子的边界。例如在T细胞急性淋巴细胞白血病(T-All)中,当CTCF介导的绝缘体缺失时,相关TAD融合,导致超级增强子与MYC基因启动子之间的异常接触,致使癌基因表达上调[34]。一项有关超级增强子在乳腺癌中异常调控的研究显示,关键转录因子RUNX3占据调控RCAN1.4基因表达的远端超级增强子,该转录因子在乳腺癌中的低表达,致使该超级增强子驱动异常,从而导致RCAN1.4表达减少。在进一步探讨RCAN1.4基因在癌组织中低表达的可能原因时,发现RCAN1.4的核心启动子区和远端超级增强子区存在于同一个TAD[35],一定程度反映了TAD与超级增强子在调控机制上的关联性。总而言之,在不同的研究背景下,TAD对超级增强子的影响机制不尽相同,但似乎都能表明超级增强子可能需要依赖于特定的TAD来实现一些转录激活的功能。
3.2 超级增强子介导非编码RNA调控靶基因表达
非编码RNA(non-coding RNA)是指不翻译成多肽的一类RNA,越来越多的证据表明非编码RNA在剂量补偿、细胞分化、细胞增殖、组织再生和适应性免疫等方面发挥着关键作用[36-37]。近期的研究表明,超级增强子不仅激活编码基因的转录表达直接调节生物功能,还通过驱动非编码RNA的转录表达间接调节生物功能。一方面,它们能促进小RNA(microRNA,miRNA)和长链非编码RNA(long noncoding RNA,lncRNA)的转录和成熟;另一方面,超级增强子本身的转录产物eRNA(enhancer RNA,eRNA)在调控基因表达方面也起着协同作用。这些非编码RNA主要包括miRNA、lncRNA、环状RNA(circRNA)和增强子RNA(eRNA)[38]。
miRNA是最早被识别的非编码RNA[39-40],Suzuki等[41]报道了超级增强子是组织特异性miRNA网络的核心,通过调节miRNA的产生影响多种肿瘤的进展。其实质是超级增强子通过增强转录,促进DROSHA/DGCR8招募和初级转录物(pri-miRNA)加工来驱动对细胞身份至关重要的主要miRNA的生物发生过程,之后再由miRNA调控相应的靶基因,从而实现超级增强子通过miRNA间接调控靶基因表达的过程。例如Ng等[42]发现,超级增强子通过miR-32抑制靶基因Tob1活性来上调FGF21分泌因子的表达,进而激活p38/MAPK信号通路,促进小鼠棕色脂肪组织的发生和白色脂肪褐变。超级增强子通常驱动组织特异性基因的高水平表达,包括具有进化保守的细胞类型特异性功能的miRNA基因。Sin-Chan等[43]报道了一种超级增强子衍生的大型miRNA簇C19MC,C19MC可被TTYH1通过其上游的超级增强子区域调控,过表达的C19MC驱动多层菊形团胚胎性肿瘤(embryonal tumorswith multilayered rosettes,ETMRs)中的C19MC -lin28a -mycn回路,促进ETMR细胞的生长;同时,Anandagoda等[44]发现,在调节性T细胞中,转录因子FoxP3可与pri-miR-142的超级增强子区域结合,促进其转录,从而抑制其下游靶基因PDE3b的表达。这些结果提示,超级增强子在基因表达调控中与miRNA存在密切的互作关系,从而在靶基因的转录活性及表达水平方面发挥重要的生物学功能,但它们详细的互作机制需要更为深入的研究来揭示。
lncRNA在超级增强子的调控机制中也发挥重要作用。研究表明,超级增强子衍生的lncRNA可以与外显子敏感的lncRNA发生互作,从而激活启动子和增强子,促进靶基因染色质环整合和核拓扑结构域的形成,促进其表达[45]。值得一提的是,一种与超级增强子密切相关的lncRNA——超级lncRNA。超级lncRNA(super-lncRNA)是指与超级增强子区域形成RNA∶DNA∶DNA三螺旋的lncRNA,可招募调控因子至超级增强子区,从而影响染色体结构,并作为空间放大器,促进超级增强子相关的组织特异性基因表达[46]。超级lncRNA最主要的作用机制之一是通过顺式作用调控邻近基因的表达。如Hand2是控制成纤维细胞向心肌细胞分化的核心转录因子之一,其表达受到邻近一种名为UPH的SE-lncRNA 的调节[47]。在人肝星状细胞(human hepatic stellate cells,HSCs)鉴定出许多lncRNAs,且发现转化生长因子β(transforming growth factor β,TGF-β)直接调控的lncRNA在超级增强子中富集[48]。考虑到肝星状细胞是肝纤维化的主要细胞类型,这一发现提示,TGF-β通过调控这些超级增强子相关lncRNAs参与肝纤维化。Fan等[49]在小鼠肝中发现一个命名为lnc-Crot的昼夜节律lncRNA,在昼夜节律波动基因Crot上游的一个超级增强子上表达,其作用受到转录因子BMAL1和REV-ERBα的影响;进一步的试验表明,lnc-Crot位点具有独立于lnc-Crot转录的增强子功能,lnc-Crot位点与昼夜节律波动基因相互作用,通过长期相互作用促进昼夜节律转录因子对附近基因的调控。
关于超级增强子所衍生的circRNAs形成机制的研究相对缺乏,且与超级增强子相关的circRNAs数据库同样不足。目前,Tang等[50]开发了circRNAs数据库TRCirc,便于检索、浏览以及可视化circRNA的转录调控信息,其中包括circRNA相关的超级增强子。Han等[51]发现,转录因子YY1与QKI(Quaking)的超级增强子和启动子结合,再经p65与启动子结合、p300作为中介,引起DNA环(DNA loops)的形成以及QKI的异常激活,异常激活的QKI会导致肝癌组织中circRNA的形成,再由circ-RNA吸附相应的miRNA,促进癌基因的表达。另有研究表明,超级增强子调控的circRNA Nfix缺失可诱导成年小鼠心肌梗死后的心组织再生,在这一过程中,转录因子Meis1与circNfix位点上的超级增强子结合可以调控其表达,而circNfix通过促进Ybx1泛素依赖性降解和抑制miR-214活性来抑制心肌细胞增殖,因此诱导circNfix的下调可促进成年心肌细胞增殖和心组织再生,显著减少心肌梗死后的纤维化区域,促进功能恢复[52]。该结果支持超级增强子通过招募主转录因子参与了诱导细胞分化和增殖的因果机制。由此,可以大体推断出超级增强子通过非编码RNA实现其对相应靶基因的调控,主要通过形成超级增强子——转录因子——非编码RNA——靶基因调控网络来实现。
3.3 超级增强子通过超级增强子RNA参与基因表达调控
研究表明,RNA聚合酶Ⅱ在超级增强子上的聚集密度大于典型的增强子,从而产生了更高水平的eRNA,即超级增强子RNA(super enhancer RNA,seRNA)[13],其实质是超级增强子自身转录形成的一种增强子RNA,但由于seRNA与超级增强子的特殊关系,对于超级增强子的影响机制与其他非编码RNA有所不同。seRNA的功能可分为顺式(cis)调控和反式(trans)调控两种。顺式调控是seRNA从其合成位点上招募蛋白质复合物来激活相邻基因,例如位于Nanog基因位点上超级增强子的seRNA缺失,会导致其相邻基因Dpp3的表达显著降低,从而通过顺式调控对相邻基因进行差异调控。同时通过染色体构象捕获技术(3C),作者证明了DPPA3启动子处远端超级增强子的环变小约50%,表明远端seRNA稳定了顺式结构中环和染色质的相互作用,从而调控了DPPA3的表达[53]。类似的,另一种命名为CARMEN的seRNA转录被发现可以直接激活下游基因的表达,调节心组织细胞分化[54]。反式调控则表明,seRNA也可能在染色体间发挥作用,从而引导靶基因的表达,这无疑扩大了SE的功能范围,例如MYOD上游非编码RNA(MNUC)是一种eRNA,可诱导形成特定的肌源性转录本,但MNUC在C2C12细胞完全不含MyoD蛋白的情况下,依旧可诱导产生多种肌源性转录本,这不同于许多基因是由MUNC以一种依赖于MyoD的方式进行调节,从而验证了MNUC是一种独立于MyoD的反式作用非编码RNA[55]。有关seRNA的研究更多报道于癌症或肿瘤相关研究中,如seRNA能够通过招募辅助因子、RNA聚合酶Ⅱ和介质,构成并稳定超级增强子和启动子区的染色质环(图3),从而调控肿瘤相关靶基因转录[56]。总之,seRNA的主要功能与eRNA类似,对于超级增强子的调控机制起到促进作用,但二者的调控机制需要更多的研究报道来揭示与阐述。
3.4 超级增强子通过相分离调控基因表达
相分离(phase separation)又称相变(phase transition),通常作为一种物理现象为人所熟知。Brangwynne等[57]在关于线虫P颗粒的研究中,发现P颗粒(一种蛋白质)并非是一种固体,而是像液滴一样,相互碰撞融合,剧烈摇晃后分散成很小的液滴,而后又很快地融合形成大液滴,该研究首次证实了生物体内的相变过程。随后的研究也表明,相分离现象不仅普遍存在于细胞内,如核仁[58]、蛋白[59]等,还存在于DNA修复[60]、神经信号传递[61]等生命活动过程,这些研究使生命科学领域的研究者认识到相分离现象可能是一种普遍的生物学现象,这为揭示真核生物体内不同数量级的DNA调控元件、RNA调控分子以及多种蛋白调控因子如何相互协调,进行复杂的基因调控提供了新的理论模型。由此,相分离成为生命科学领域的研究热点,同时也为解释超级增强子的作用机制提供了新的启发。
2017年Hnisz等[62]首次以相分离理论解释超级增强子如何参与基因调控,虽然该观点没有充足的试验证据,但为进一步探索哺乳动物基因控制原理搭建了一个基本框架。随后,该作者团队的研究结果表明,转录共激活因子BRD4和MED1可以结合在超级增强子处发生相分离并形成液滴,通过相分离从细胞核中隔离出转录相关组分,从而实现转录过程的区室化(compartmentalization)反应,促进转录过程的正常进行,而固有无序区域(intrinsically disordered regions,IDRs,是内在无序蛋白的主要功能位点,能促进多种细胞功能)在其中发挥关键作用,该研究为超级增强子通过相分离调控基因表达的模型提供了试验证据[63]。同时,有研究证明超级增强子内部,转录因子激活域通过与共激活剂形成相分离的凝聚物来影响基因表达[64],相关研究结果奠定了相分离的理论基础,为进一步阐明相分离的独特功能和实现过程提供了重要基础,且超级增强子和相分离的联合研究为未来人类重大疾病的攻克、哺乳动物重要表型性状的调控,从基因水平提供了全新方法与思路。
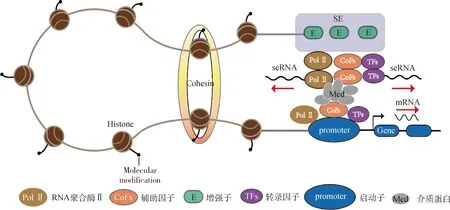
seRNA招募RNA Pol Ⅱ、CoFs和Med,形成和稳定SE和启动子的染色质环,调控靶基因转录seRNA recruits RNA Pol Ⅱ,CoFs and Med to form and stabilize the chromatin loops of SE and promoters,regulating target gene transcription图3 超级增强子通过seRNA参与基因表达调控Fig.3 Super enhancers participate in the regulation of gene expression through seRNA
3.5 超级增强子参与表达调控的其他可能方式
由于研究材料的不同,超级增强子在不同生物细胞中的调控方式具有较大差异,但可以确定的是,超级增强子可以通过重要信号通路来参与细胞发育或肿瘤的发生发展过程中的基因表达[15,65]。信号通路作为机体内实现诸多细胞功能的信号转导途径,已被证明可以通过控制活性增强子转录的起止驱动特定细胞生长[66]。这在一定程度上提示具有更高转录活性的超级增强子可能与信号通路存在某种关联。近来Sun等[67]在研究多能干细胞分化作用机制时,发现Hippo信号通路对超级增强子具有调控作用。在该机制当中,Hippo信号通路的效应因子YAP发挥关键作用,表现为能协同多能干细胞的核心转录因子和其他超级增强子结合蛋白共同作用超级增强子(图4),实现对关键基因表达的调控。在YAP富集增加的位点上,干细胞转录因子的富集也相应增高,导致一些普通增强子转变为超级增强子,使其调控的基因表达增强。同时YAP也会引导激酶Mst(Kinase)缺失的胚胎干细胞中新的超级增强子形成,以及在相分离过程中发挥重要作用的Med1组分蛋白在谱系相关基因上的凝聚增强。该研究是在胚胎干细胞中所揭示的一种通过调节YAP结合超级增强子形成的谱系分化的新机制,同时该机制也体现了Med1作为转录共激活因子所具备的相分离的生物学功能。需要强调的是,上述信号通路对于超级增强子的调控,是一种基于特定组织细胞中发现的新型超级增强子调控机制,至于在其他组织细胞中是否也存在信号通路对于超级增强子的调控需要更多研究结果来说明。
超级增强子在各种特定组织、疾病中的调控机制虽略有差异,但基本都是在各种因素的影响下,形成一个与启动子实现物理接触的大型复合体。但相分离模型的提出使得这种以往的假设模型无法解释染色质相分离凝结水的形成,因此超级增强子更可能是通过空间接近来实现转录激活[68]。
4 超级增强子在动物重要表型调控中的应用前景
由于超级增强子本身具有极高的转录激活能力,且多富集于基因组的变异区,在以往研究中,主要报道了其在维持细胞身份、细胞发育以及控制疾病相关基因方面具有重要作用。通过借鉴当下超级增强子在模式动物和一些疾病中的研究成果,如人类癌症相关,小鼠模型研究中脂肪形成、乳腺发育以及肌源性分化等,发现超级增强子在家养哺乳动物表型调控的研究中具有重大应用前景,这些研究结果能够为未来农业动物的研究应用提供启发和方向。例如在乳腺组织中的研究表明,富含乳腺转录因子(如STAT5)的超级增强子可以控制与乳腺组织分化相关基因的转录[69],在对泌乳期的乳腺组织进行分析时,鉴定出440个乳腺特异性超增强子,其中一半的超级增强子相关基因是由转录因子STAT5在孕期诱导,且在乳腺组织的表达高度丰富[17]。而在肌源性分化的研究中,有研究表明,超级增强子在肌源性分化中的作用主要通过增强子RNA来实现,例如超级增强子产生的Serna-1和Serna-2在体内外均能促进小鼠肌源性分化,这一过程是由主转录因子MyoD诱导Serna与核不均一核糖核蛋白L(heterogeneous nuclear ribonucleoprotein L,HnRNPL)相互作用,在肌源性分化过程中激活靶基因转录[70]。另外使用小鼠模型进行肌源性分化研究时发现,转录因子PAX7通过与一个或多个肌肉相关的转录因子协作来组装超级增强子,从而对肌肉中关键靶基因进行调控[71]。
同样,超级增强子也参与了脂肪的形成、分化以及代谢过程,并在其中发挥重要作用。例如在棕色脂肪形成过程中,研究者利用甲基转移酶MLL4和乙酰转移酶CBP在小鼠脂肪细胞中鉴定出一些超级增强子,结合脂肪发生过程中的转录组数据,发现其中一部分超级增强子与一般脂肪生成基因相关,其他则为棕色脂肪特异性(brown-specific)超级增强子[72],此外,BET含溴结构域蛋白4(bromodomain-containing protein 4,BRD4)可以通过形成新的超级增强子来传导脂肪生成程序,从而驱动PPARG和CEBPA转录,促进脂肪细胞分化[73]。在脂质代谢方面的研究发现,超级增强子所驱动的转录因子KLF6支持脂质代谢基因的表达,促进PDGFB的表达,PDGFB激活mTOR信号和下游脂代谢调节因子SREBF1和SREBF2的表达[74]。在肥胖治疗方面,相关研究也能证实超级增强子所发挥的重要作用,例如在小鼠体内存在一种棕色脂肪组织(brown adipose tissue)特异性的超级增强子能通过相关miRNA,促进小鼠棕色脂肪组织的发生和白色脂肪褐变,可以被用于治疗肥胖[42]。这些研究结果充分说明超级增强子在哺乳动物重要表型调控中的关键作用,对未来解析家养动物的营养物质吸收、脂肪沉积以及能量利用等生物学过程提供了研究思路。
近来,超级增强子在哺乳动物中的研究有了最新进展。Luan等[75]在猪肝中鉴定出了1 711个超级增强子,而肝在新陈代谢中起重要作用并能够影响猪的生产,该研究既为未来超级增强子在猪的基因调控提供了参考,同时也为下一步超级增强子在猪的表型调控研究奠定前期基础。另外一项研究通过对斑马鱼与小鼠和人类的超级增强子进行序列和功能比较分析,发现超级增强子与物种基因组大小成正比;且42%的斑马鱼超级增强子位于与小鼠和人类超级增强子相关的同源序列附近,揭示了哺乳动物间超级增强子在功能上存在高度保守性[76]。此外,研究人员开发了包括绵羊、鸡等家养哺乳动物在内的超级增强子数据库,将极大促进家养哺乳动物超级增强子的研究进展[77]。当前家畜超级增强子相关研究主要是基于表型差异背景的超级增强子测序分析,对于其参与表型变异调控的分子功能验证仍有欠缺。
超级增强子在哺乳动物重要表型的基因表达调控过程中发挥重要作用,主要表现在人或小鼠等模式动物上。尽管相关研究在家养哺乳动物中报道较少,但当下科研人员不断开始尝试进行家养哺乳动物超级增强子的鉴定分析,基本证实了可以通过超级增强子调节特定基因的表达活性来调控相关机体表型,这对于揭示表型性状背后的分子机理、开展家养动物主要表型调控具有重要的参考价值,同时也可为家养哺乳动物分子育种提供一种新的研究思路与方法。
5 展望
超级增强子能驱动控制细胞身份基因的表达,在机体发育和疾病发生过程中起到重要作用。基于超级增强子在基因表达调控中显著的正向调节作用,如促进动物脂肪分化、乳腺组织分化以及肌源性分化相关基因的表达,这对于揭示哺乳动物此类表型性状背后的分子机理具有重要意义。随着生物学技术的发展,对于超级增强子的认识正在逐渐拓宽,如超级增强子参与细胞代谢重编程[78]等。随着分子生物学研究从组织样(bulk cell)到单分子(single molecular)及单细胞(single cell)水平的转变[79],结合CRISPR/Cas9[80]、空间转录组、染色质可及性等新兴技术将为进一步了解超级增强子的生物学作用提供空间,也为详细探究超级增强子功能机制提供了更多可能,从而期望对超级增强子在哺乳类动物的相关疾病或性状解析中提供参考与启示。
猜你喜欢
生命科学研究(2022年3期)2022-09-13 08:15:26
生命科学研究(2020年6期)2021-01-09 11:34:24
学苑创造·A版(2020年9期)2020-10-13 09:41:02
学苑创造·A版(2020年12期)2020-01-07 14:07:23
中国外汇(2019年15期)2019-10-14 01:00:34
遗传(2019年1期)2019-01-30 06:39:30
生物技术通讯(2017年4期)2017-11-06 01:24:56
小学生学习指导(低年级)(2017年10期)2017-10-10 01:00:05
作文教学研究(2016年1期)2016-07-05 12:22:47
湖北师范大学学报(自然科学版)(2015年2期)2016-01-10 08:41:55
