
基于WinRHIZO的川中紫色土坡地冬小麦根系生长发育
2021-12-31 05:23柯尊嵩
西南农业学报 2021年11期
柯尊嵩,冯 波*,李 帆
(1.吉林大学新能源与环境学院,吉林 长春 130021; 2.长安大学水利与环境学院,陕西 西安 710054)
【研究意义】四川历来是中国重要的粮食生产基地和国家农业经济体的重要组成部分,小麦和水稻在川中粮食产量上占到了60%。然而四川多山地丘陵,使得多种高效的机械化麦稻轮作系统不能得以应用,所以在川中坡地上最常用到的依旧是人力锄具耕作,但这种耕作方式效率低下,同时对土壤的耕作侵蚀效果非常严重[1]。土壤侵蚀会对耕地上植物上部产生直接影响,本研究对紫色土坡耕地上小麦根系的研究,对完善耕地侵蚀模型、保证国家粮食安全、提升农副产品经济价值都具有重要意义。【前人研究进展】储栎泉等[2]研究发现,在四川耕地中主要以坡耕地为主,坡耕地上壤中流造成的养分重分布具有时空性。苏正安等[3]研究表明,紫色土具有土层浅薄、结构性差和有机质含量低的缺点,同时又是极不稳定的土壤型。在长期的风吹日晒下极易发生风蚀,并且土壤内部养分会随水流发生较大重分布。在线性坡面肥料一般是从坡耕地上部流失,在下部堆积[4]。国内在紫色土区的土壤“浅薄化”对水肥影响方面也进行了大量的研究。如马胜兰等将紫色土层厚度分为5个不同的级别进行研究,指出60 cm以上厚度的紫色土可以维持基本稳定的生产力水平,土层厚度对玉米影响格外显著,超过60 cm厚度的土壤对小麦的生物量及根重差异不显著[5]。以上结果对紫色土的土壤生产力就行了研究,但目前针对川中丘陵地区坡地紫色土层对于小麦根系的生长研究仍然较为匮乏。小麦属于禾本科植物,拥有丰富的根系[6]。根系是植株最重要的是营养器官,在土壤中承担了水肥吸收、植株无氧呼吸等重要过程[7]。近年来,大量的学者采用直接或者间接观察的方法描述植株的根系生长,但大多数实验是在实验室内多种条件固定的环境下进行的,同时对植株的培养也都选择以管栽、盆栽的方式进行[8]。但作物在自然条件下的生长状态与实验室内有着明显差异,实验室内根系实验忽视了植物根系对环境变化的自适应性,使得作物根系形态研究具有很大的局限性。加之作物地下部分难以测量,使得植株根系发育特点以及可视化的研究一直严重滞后于地上植株[9]。另外,许多研究将注意力集中在无人扰动的自然土体上的乔灌草根系上,而关于坡耕地尤其是紫色土区的坡耕地上农作物的根系研究报道较少。【本研究切入点】本研究以四川丘陵地区紫色土坡地上的小麦为研究对象,分析了小麦根系在不同坡位上的生长状况,结合机械化与传统耕作模式,探讨适合坡地耕作的形式。研究了在不同坡位处小麦根系所表现的根系表型性状及不同直径小麦根系对不同坡位的响应。【拟解决的关键问题】利用根系表型性状对川中丘陵区紫色土坡地上的小麦作生长发育综合性评价,揭示小麦根系在坡地上的发育规律,为川中丘陵区紫色土坡地上农作物种植理论进行理论补充。
1 材料与方法
1.1 实验材料
本实验材料选用市售蜀麦830,于2018年11月中旬播种,种植在四川遂宁水土保持实验站内。
1.2 研究区概况
试验区选择在四川农业大学的水土保持实验站内,该站位于四川省遂宁市,地处北纬30°10′50″~31°10′50″,东经105°03′26″~106°59′49″。该实验区域以亚热带的典型湿润季风气候为主,不存在冻土现象。试验区的坡耕地具有坡度大,耕地少而散,土壤为碱性,农机难以进入,水力侵蚀严重等特点。在该地区,传统的铁农具(镐头,镰刀,铲子等)是该地区的主要农具。为了方便耕作,经常使用下坡耕作。
1.3 实验设计
前人的研究中,为了减小不可控因素的影响同时方便观察,大多对作物进行管栽、盆栽及温室栽种[8]。但容器和比较理想条件下的自然条件是无法完全替代农田土壤水库以及农田养分库对水肥的缓冲作用,同时较小的种植空间对小麦的根系生长发育起到了抑制作用,无法达到完全自然条件下小麦根系生长过程中的性状,本研究结合遂宁本地的气候、工业经济发展水平,考虑到,不同耕作模式对团聚体的扰动效果不同[10],采取传统农作与机械犁耕作的对比试验。在设置实验组时进行了机械犁作组与传统犁作组的对照设置,试验场从北向南定向,长度约为10 m,土壤深度大于0.8 m,并且碎石大部分位于0.8 m以下,两种种植方式的对照组平行排列。同时集中种植小麦,籽粒平均尺寸小于5 cm。进行采样时,所采样的小麦处于乳熟期后期,根系保持完整。试验田周边设置有1 m保护行,试验田内进行箱式种植,带有0.3 m排水沟系用以排水防涝。播前施加农家肥4 cm,播种完成后取压碎后的紫色土盖籽。为确保采样根系的完整性,方便对根系分布规律的研究,我们借鉴农淦[11]等人的环式取土法(图1)。同时使用铁楸对半径20 cm的土样进行整体采样。采样前,将坡地按景观位置分为五个部分,分别为坡顶、坡肩、坡中、坡脚、坡趾,分别距离坡顶0、5、10、15、20 m,根据不同的景观位置对小麦根系采样,在每一个景观位置上取其几何中心(图2)。每个区域,在几何中心点采用农淦的方法,在被取植物的中心画一个半径为10 cm的圆,并对被圆覆盖的整个土壤体进行环刀采样。为了确保采样不会互相影响,需要使用位于相同景观位置的植物,相距20 cm以上。同地多次取样,分别测算相应数据。

图件引自参考文献[11]图1 小麦根系环刀取土方式Fig.1 Soil sampling method of wheat ring cutter

图2 坡地取样点设置Fig.2 Sampling point setting of slope land
1.4 指标测定
1.4.1 单根抗拉强度测量 植株收获后,对小麦根系进行清理后,将同一植株相同直径的根系归类,将要测试的单个根放在拉力测试仪的两个夹具之间,启动应变控制式直剪仪(图3),然后以手动匀速向上移动上夹具,直到根断开。数据采集系统每0.01秒自动记录一次拉伸和位移,以分析单根的变形特性;实验后,计算机系统自动生成根部拉伸时的极限抗力F和断裂时根系的伸长率△L值,并通过计算机计算出单根拉伸强度(应力)和应变值,其中单根抗拉强度计算公式为:
P=4F/ΠD2
式中,P为抗拉强度(MPa);F为抗拉力(N);D为根系断裂处直径(mm)。在本研究中将根系根尖、根中、根末三处的直径测得取均值作为最终的实验根径。

图3 ZJ型(EDJ-1)应变控制式直剪仪Fig.3 ZJ (EDJ-1) strain controlled direct shear apparatus
1.4.2 单根轴向抗拉应变计算公式
ε=△L/L
方法3:蒸发鉴别法。取一干净、干燥的玻璃片,然后分别在不同位置滴等量水样,待其完全蒸发后,白色残留物多的水样为硬水,反之为软水。
式中,ε为应变;△L为试验根在均匀拉力下导致的变形量(mm);L为试验根初长(mm)。
1.4.3 根系体积密度计算的公式
RVD=Vd/Vs
式中,RVD为根体积密度(cm3/125 cm3);Vd为根系体积(cm3);Vs为土壤积(cm3),其中根钻取土体积为125 cm3。
1.4.4 根系拓扑指数计算公式
Fitter[12]等和Bouma[13-14]等提出根系拓扑结构以鱼尾状分支和叉状分支为主要分型模式。并提出拓扑指数表达式:
Ti=lgA/lgM
1.4.5 根系指标测定 在将根系洗净去除泥沙之后,采用数字化扫描仪器(Lenovo M7605D)对根系形态进行扫描,扫描前小麦根系根系需分离压实,以白色面板做底板(图4)。并采用WinRHIZO (version4.0b,Rengent Instruments Inc.,Canada)根系分析系统对根系生物特性进行测定,应用Origin9.1对测得的数据进行分析。
2 结果与分析
2.1 不同耕作模式下小麦根系拓扑指数对景观位置的响应
不同耕作模式下,小麦根系的拓扑指数Ti呈现出差异。如表1所示,机械耕作组均值为0.947,传统耕作组均值为0.948,两组样本Ti总体接近1,机械耕作组拓扑指数与传统耕作组差异不显著,表明在12°坡地上,机械耕作与传统耕作对小麦根系总体不构成显著影响。机械耕作方式得到的小麦根系内部数平均为45.2,外部数均值为55.6,传统耕作方式下的小麦根系内部数平均为54.8,外部数均值为 67,机械耕作方式小麦根系的内部数与外部数均少于传统耕作方式,表明在12°坡地上,传统耕作下的小麦根系发育情况要更好于机械耕作。顺坡方向上,机械耕作组小麦根系内部数在坡顶与坡趾分别达到了最大值46和56,外部数分别达到了最大值56和69,传统耕作组小麦根系内部数在坡顶与坡趾分别达到了最大值72和55,外部数分别达到了最大值89和68,表明在顺坡方向上,小麦根系在坡顶与坡趾分型发育最为明显,传统耕作模式下与机械耕作模式下小麦分叉数数量均表现出凹状抛物线趋势。顺坡方向上,传统耕作组中不同坡位小麦根系Ti保持,0.966~0.915,机械耕作组中不同坡位小麦根系Ti保持在0.954~0.935,两个实验组Ti均更加靠近1,表明不同景观位置的小麦根系存在较多的外部连接,根系近似于鱼尾状分支模式,主根明显且分支较少。机械耕作组中小麦qa与qb略小于传统耕作组,说明传统耕作组的小麦根系拓扑结构比机械耕作组复杂,有相对更多的次级分支。

图4 小麦根系样本Fig.4 Wheat root samples

表1 两种耕作方式不同坡位处小麦根系拓扑指数
2.2 不同根径域的根系抗拉力与拉裂距离的相关性研究
如图5所示,0~0.1 mm根系抗拉强度抗拉力主要处在0.25~1.5 N之间,断裂距离主要在0.25~1.25 mm之间,0.1~0.2 mm根系抗拉力大都处在0.5~4 N之间,断裂距离大多在0.5~7 mm之间。0.2~0.3 mm根系抗拉力大多处在2~7 N之间,断裂距离大多在1~4 mm之间。0~0.3 mm组根系中0.1~0.2 mm根系较0~0.1 mm根系在抗拉强度和断裂距离方面都提高了2~4倍,0.2~0.3 mm总体的抗拉强度断裂距离比0.1~0.2 mm增加了2~3倍,说明0~0.3 mm组小麦根系抗拉力、拉伸距离均与直径成正相关关系。0.3~0.4 mm根系抗拉强度大多在2~6 N之间,断裂距离断裂距离大多在2~7 mm之间,0.4~0.5 mm根系抗拉强度大多处在2~6 N之间,断裂距离大多在1~6 mm之间。两组变化趋势总体比较平稳,但0.3~0.4 mm组,变化值域明显加大。>0.5 mm根系抗拉强度大多在4~8 N之间,断裂距离在3~7 mm之间,变化范围最不稳定,说明在>0.3 mm直径域的小麦根系,根系抗拉力、断裂距离与根径没有直接相关性,其中每个根径域内的根系在根系抗拉力、断裂距离方面差异不显著。

图5 不同根径域的根系抗拉力与断裂距离Fig.5 Tensile strength and fracture distance of roots with different diameters
通过对不同直径根系抗拉力与断裂距离图进行匹配性分析, 0.3 mm以上根系的匹配性明显降低,表明直径增加匹配性降低,较细小的根系抗拉力与断裂距离匹配性良好,具有较好的线性关系。0.3 mm及以上小麦根系抗拉强度均值为6 N,0.3 mm以下小麦抗拉强度均值为3 N,0.3 mm及以上小麦根系断裂距离均值为3.5 mm,0.3 mm以下小麦断裂距离均值为2 mm,表明在土体中0.3 mm及以上直径的小麦能承担更大的抗拉强度,其提供了植株对土体锚固作用的绝大部分。相关性方面,0.3 mm以上根系中存在抗拉力与断裂距离线性关系差,表明较粗小麦根系的外层根皮与内层细根具有明显差异化的抗拉强度,小麦根系的外层根皮能够对内部细根起到的保护作用有限。
2.3 不同景观位置小麦根系的生物表征比较
小麦根系的根系表面积、覆盖面积与坡位正相关性明显。在川中丘陵区紫色土坡地进行机械种植条件下,小麦单株根系总表面积不超过10 cm2,均值不超过5 cm2,并且在顺坡方向上,小麦根系的表面积逐渐增大,在坡脚位置达到最大值3.95 cm2,而坡趾处较坡脚处小麦根系表面积有所降低(图6-d)。小麦单株根系覆盖面积均值13 cm2,在坡脚位置达到最大值12.5 cm2(图6-a),在坡趾也比坡脚处小麦单株根系覆盖面积更小,说明在川中丘陵地区12°紫色土坡上,采用机械耕作后,小麦根系有利的生长位置在坡肩以下,并且坡脚位置为最优生长位置,坡趾生长较优,但并不是最优生长位置。
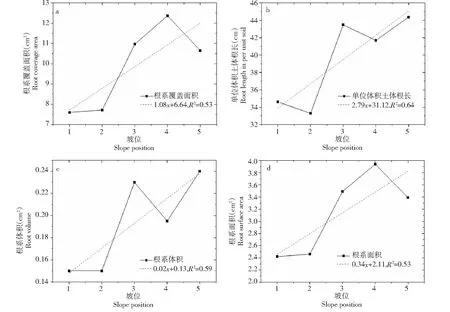
图6 不同景观位置小麦根系生物表征与坡位拟合图Fig.6 Biological characterization of wheat roots and slope fitting map at different landscape locations
小麦根系在单位体积土体中的根长、根系体积与坡位也有明显正相关性。单位体积土体中根长最长为44.5 cm,坡肩下部根长均值为坡肩上部的1.3倍(图6-b)。小麦体积在坡趾、坡中达到最大值0.235 cm3,坡肩与坡顶为0.15 cm3(图6-c),说明坡地下部更加肥沃蓬松的土壤对小麦根系延伸与根系根径的生长更加有利。
2.4 小麦根系多因素相关性分析
对小麦根系各参数进行多因素相关性分析,结论如表2所示:坡位与小麦根系面积、根系体积、不同根径域的小麦根系抗拉强度均有显著相关性(P<0.05),且坡位与小麦根系面积、根系体积表现出正相关关系(r>0),坡位与不同根径域的小麦根系抗拉强度表现出显著负相关性(r<0),坡位与拓扑指数无明显相关性(P>0.05),说明顺坡方式向上,反应根系面积与根系体积的指数逐渐增高,而不同根径域的小麦根系抗拉强度呈现降低,根系的分型强度对坡位反应不明显。根系面积与根系体积、拓扑指数、>0.3 mm小麦根系抗拉强度有显著相关性,与0~0.3 mm根系抗拉强度则无明显相关性,且根系面积与根系体积呈正相关关系,与拓扑指数和>0.3 mm小麦根系抗拉强度呈负相关关系,说明根系表面积与根系体积为直接相关因素,体积越大表面积一般也越大,而根系表面积的扩大会抑制根系分型的发生,并且降低>0.3 mm小麦根系的抗拉强度。根系体积与根系拓扑指数和不同根径域的小麦根系抗拉强度无显著相关性,拓扑指数也与不同根径域的小麦根系抗拉强度无显著相关性。
3 讨 论
Niu等[15]的研究表明,植株对水肥吸收主要受到植株根系的空间分布与形态差异的影响。作物的根系会在土壤中随壤中流造成的水肥重分布变化而主动发生适应性的改变[16]。本实验中,在川中12°紫色土坡地上以机械耕作和传统耕作两种方式进行小麦种植,发现以两种耕作方式种植的小麦根系在紫色土坡地的极端条件下均呈鱼尾状分支结构,顺坡方向上,小麦根系在坡顶与坡趾分型发育最明显,小麦分叉数量均呈凹状抛物线趋势,在不同景观位置的小麦主根系明显且分支较少,而对比两种耕作模式下小麦根系的分型发育结果,传统耕作模式下小麦根系拓扑结构更加复杂,这是由于鱼尾状的根系分支结构能够有效提高根系的覆盖面积,扩大有效营养吸收利用的空间,从而高效的在贫瘠且水土流失严重的紫色土坡地上获取养分,根系的鱼尾状拓扑结构特征,有利于为拔节或灌浆提供物质基础[13,17]。
同时根系体内部也拥有叉状分支结构,能在土层中形成比较牢固的土壤-根系复合体。在在坡地坡趾处土层疏松,更有利于小麦侧根发育,而坡顶处水土流失严重,小麦根系受环境胁迫,被迫进行主根供养,造成两个不同景观位置小麦主根上的分支少且主根明显的特点。综上所述,小麦根系所表现出的特征,与紫色土坡地的所处环境、土壤性质密切相关。作物地下竞争环境的不均匀性,土壤理化状态的空间变异性,或者水肥供应的空间方向性,种子自身的品种特性等,均能造成根系在土体空间中的不均匀分布[18-20]。

表2 小麦根系形态学特征与力学特征相关性表
不同直径小麦根系具有不同的力学特性。厉翔等[21]研究表明,植株在土体中的根系对于水土流失和土壤侵蚀的防治有很大的作用,在载荷相同的情况下,有根系存在的土块会产生远小于无根土块的位移,并且小麦在达到成熟期以后根系对土体的固持能力明显要好于其他生育期,同时越大的根土接触面积会进一步的提高水土保持效果。本实验研究发现,0~0.3 mm根径的小麦根系抗拉力、断裂距离与根径成正比,>0.3 mm根径的小麦根系抗拉力、断裂距离与根径没有直接相关性,并且>0.3 mm根径域的小麦根系在根系抗拉力、断裂距离方面几乎差异不显著,0~0.3 mm根径的小麦根系抗拉力与断裂距离的匹配性要好于>0.3 mm根径域的小麦根系,但>0.3 mm根径域的小麦根系抗拉力为0~0.3 mm根径的小麦根系的两倍。这是由于小麦根系的特定结构决定的。小麦根系可以区分为较细根系和较粗根系两类,较粗根系相较于较细根系拥有很厚的根皮,这层根皮保证了内根的较大延展性,当进行抗拉测试时断裂的外皮会顺着内跟进行滑移,从而防止了内根的直接断裂,这对小麦根系形成牢固的根土复合体起到了极其重要的作用[22]。
本实验中,小麦根系的根系表面积、覆盖面积、单位土体根长、根系体积与坡位正相关性明显,均在坡脚处达到最大值;小麦根系在斜坡上的坡肩以下位置土体中,有更长的单位土体根长与根系体积。这是因为顺坡方向上水肥会有向下堆积的趋势,越向下部景观位置,土壤肥力蓬松度越高,但坡趾处接近于排水沟渠,水土流失也比较严重[23]。
诸多研究表明,根系是植株最重要的营养器官,作物根系的生长发育受到多种因素的综合影响[24-27]。本实验对坡位、小麦根系面积、根系体积、拓扑指数、不同根径域的小麦抗拉强度进行相关性分析,发现顺坡方式向上,反应根系面积与根系体积的指数逐渐增高,而不同根径域的小麦根系抗拉强度呈现降低,根系拓扑指数对坡位反应不明显。根系面积与根系体积呈正相关关系,与拓扑指数和>0.3 mm小麦根系抗拉强度呈负相关关系,说明根系表面积与根系体积为直接相关因素[28-29],体积越大表面积一般也越大,而根系表面积的扩大会抑制根系分型的发生,减小体积的进一步扩大,同时降低>0.3 mm小麦根系的抗拉强度。这与王宇蕴等[30]人和陈杨[31]对小麦~蚕豆间作的研究结论也具有一致性,小麦可以显著减少根系平均直径,增加小麦根系表面积,以提高小麦产量。
4 结 论
12°紫色土坡地上,相同景观位置的小麦根系拓扑指数不受耕作模式的显著影响,传统耕作模式更有利于小麦根系分型发育;两种耕作模式下的小麦根系均呈鱼尾型分支结构,且在顺坡方向上呈凹状抛物线趋势,传统耕作模式下的小麦根系拓扑结构较机械耕作更加复杂。不同根径域的小麦根系抗拉力、断裂距离有差别,0~0.3 mm根系根系抗拉力、断裂距离与根径成正相关,>0.3 mm根系差异不显著;>0.3 mm根系为小麦根系中承担“锚固作用”的主要部分,小麦根系较厚的外层表皮,能对内层根系起到一定程度的保护作用。景观位置与小麦根系生物表征有明显的正相关关系,小麦根系有利的生长位置在坡肩以下,并且坡脚位置为最优生长位置,坡趾生长较优,但并不是最优生长位置,坡肩以上不适于小麦根系生长。多因素相关性分析显示,坡位与小麦根系面积、根系体积正相关,与拓扑指数无明显相关性,与不同根径域的小麦抗拉强度为负相关;根系表面积与根系体积为直接相关因素,体积越大表面积一般也越大,而根系表面积的扩大会抑制根系分型的发生,减小体积的进一步扩大,同时降低>0.3 mm小麦根系的抗拉强度。通过对川中丘陵区紫色土坡地上小麦根系生长发育进行研究,可作为坡耕地上作物研究的重要组成,一定程度对坡耕地上农作物种植、管理提供指导。
猜你喜欢
现代园艺(2022年7期)2022-11-19
林业科技(2022年5期)2022-10-08
农业技术与装备(2022年3期)2022-05-31
数学大王·低年级(2022年5期)2022-05-21
山东农机化(2022年1期)2022-03-05
腐植酸(2021年2期)2021-12-04
湖北农业科学(2019年22期)2019-12-23
高中生学习·高二版(2017年9期)2017-10-25
幼儿智力世界(2016年11期)2017-02-21
中学生数理化·高二版(2008年10期)2008-06-17
