
六盘山辽东栎林种群结构和空间分布格局
2021-12-27 04:22:36张中惠郭建斌王彦辉
浙江农林大学学报 2021年6期
张中惠,郭建斌,王彦辉,王 晓
(1. 北京林业大学 水土保持学院,北京 100083;2. 中国林业科学研究院 森林生态环境与保护研究所 国家林业和草原局森林生态环境重点实验室,北京 100091)
种群结构是种群生态学的重要研究内容,其中年龄结构[1]、静态生命表和存活曲线是反映种群现状及存活、死亡过程的重要特征指标[2]。辽东栎Quercus wutaishanica是中国暖温带地带性植被和常见的落叶阔叶优势树种,分布范围较广,具有耐严寒、耐干旱、耐贫瘠的特点,主要分布在中国华北地区,其中,辽东栎天然次生林是六盘山区的顶级群落,辽东栎作为群落主要树种或优势树种,其生长状况和种群结构特征等可以帮助识别当地自然环境特征,因此,以往关于辽东栎的研究主要集中在辽东栎种群结构和空间分布格局等方面[3-8]。伊力塔等[3]通过分析不同群落类型下辽东栎的点格局发现:在辽东栎种群不断成长过程中,其分布特征由聚集分布变为随机分布。张育新等[4]研究表明:分析种群空间格局分布有助于准确认识种群生物学及生态学特性。MASAKI等[5]对群落中优势种的种群结构及空间分布进行了研究。胡尔查等[6]分析了乌拉山白桦Betula platyphylla的种群结构,认为在生存环境相似时,自身生物学和生态学特性是影响分布格局的最主要因素。王道亮等[7]通过研究黄龙山林区辽东栎天然次生林发现:格局分布与尺度大小密切相关。研究种群空间结构能够反映群落的生境条件和种群本身生物学特性之间的相互影响,同时揭示森林的天然更新机制以及发展方向[6]。以往只有对六盘山辽东栎林的空间分布特征、林分结构、生长发育以及天然更新情况有研究[8],缺少对六盘山辽东栎天然林种群结构以及种内各龄级之间的关系研究。鉴于此,本研究对六盘山区辽东栎天然林的径级结构以及不同径级阶段个体的空间分布以及相关性进行分析,研究其动态特征和影响因素,有助于掌握六盘山辽东栎的生长发育现状并深入认识其群落发展过程,为维护六盘山及周边地区辽东栎林的稳定空间分布格局提供理论基础。
1 研究地区与研究方法
1.1 研究区概况
研究区位于宁夏六盘山南部的香水河小流域 (35°27′22.5″~35°33′29.7″N,106°12′10.6″~106°16′30.5″E),该区处于暖温带半湿润区向半干旱区过渡带,土壤以灰褐土为主,水分条件较好。小流域森林覆盖率高达82.91%,天然次生林占小流域面积的58.51%,主要树种有华山松Pinus armandii、辽东栎和白杨Populus alba等;人工林占小流域面积的24.00%,主要树种有华北落叶松Larix principis-rupprechtii等;灌木林占小流域面积的12.01%,灌木主要有峨眉蔷薇Rosa omeiensis、虎榛子Ostryopsis davidiana、野李子Prunus salicina等;草地零星分布在沟底,草本主要有狼针茅Stipa baicalensis、早熟禾Poa annua、艾蒿Artemisia argyi等;草甸占小流域面积的4.64%,草甸组成植物主要有蕨Pteridium和紫苞风毛菊Sanssurea iodostegia等[9]。从表1可见:辽东栎林分布在海拔1 900~2 280 m的山坡上,分布坡向主要为半阴坡、半阳坡和阴坡;坡度主要为陡坡;坡位上、中、下均有分布,常伴生有少量山杨Populus davidiana等。
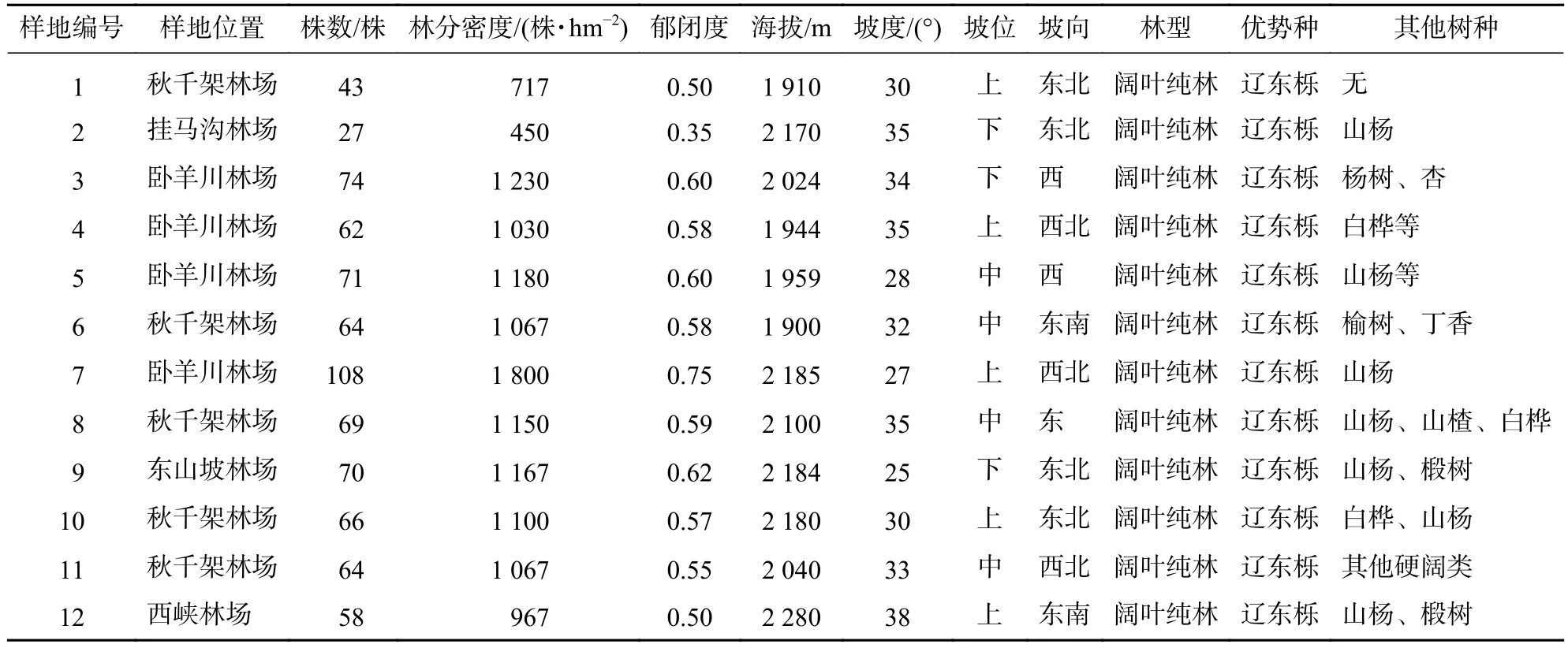
表1 六盘山辽东栎样地信息Table 1 Location factors of Q. wutaishanica in Liupan Mountain
1.2 研究方法
1.2.1 样地概况 本研究采用2015年调查的12块样地,面积为24 m×24 m。调查样地内辽东栎及其他树种胸径DBH≥5 cm的树高、胸径、冠幅及枝下高、郁闭度、林分密度、坐标位置等指标,样地内辽东栎多有丛生,株数按1丛为1株,并从基部往上数确定枝条数。标准地的边界使用罗盘仪测角、皮尺量距,用布鲁莱斯测高仪测得树高,用胸径尺测量胸径,用测距仪确定每株树的坐标位置,并用手持全球定位系统(GPS)记下每个样地的经纬度、海拔等,林分密度通过株数除以样地面积得出,郁闭度采用步测法[10]。
1.2.2 种群径级结构划分 根据样地调查及文献[11-12],确定起测胸径为5 cm,径级划分为:Ⅰ级(5 cm≤DBH<10 cm)、Ⅱ级 (10 cm≤DBH<15 cm)、Ⅲ级 (15 cm≤DBH<20 cm)、Ⅳ级 (20 cm≤DBH<25 cm)、Ⅴ级(25 cm≤DBH<30 cm)、Ⅵ级(30 cm≤DBH<35 cm)、Ⅶ级(35 cm≤DBH<40 cm)、Ⅷ级(DBH≥40 cm)。并以径级为横坐标,各径级内个体所占比例为纵坐标,绘制辽东栎径级结构图。
1.2.3 种群静态生命表及存活曲线的绘制 统计各径级内辽东栎个体数,根据函数关系式计算各函数值,绘制六盘山辽东栎种群的静态生命表。然后,根据静态生命表,以径级为横坐标,绘制辽东栎种群的存活系数(l nlx)、死亡率(qx)和消失率(Kx)图[13]。

式(1)~(7)中:x为径级,取值1、2、 ·· ·、8;lx为标准化存活个体数;ax为辽东栎存活株数;a1为辽东栎第1径级存活株数;dx为径级间隔期内的标准化死亡数;qx为径级间隔期内的死亡率;Lx为区间寿命;T为总寿命;Ex为期望寿命[14],Kx为消失率。
1.2.4 生存分析 以径级为横坐标,分别绘制辽东栎种群存活率S(x)和积累死亡率F(x)、死亡密度f(x)和危险率 λ (x)图[15]。


式(8)~(11)中:x为径级;Sx为存活率;hx为径级宽度。
1.2.5 种群分布格局分析 对辽东栎纯林的空间格局,使用成对相关函数g(r)进行描述,用尺度r作为横坐标,上下包迹线作为纵坐标,进行绘图。即:

式(12)中:函数K(r)是从种群内随机抽取的个体落在以定点为圆心、r为半径的圆内期望值;g(r)函数是基于成对个体间距离的关联性函数,用种群实际分布数据计算不同研究尺度下的g(r)值,若在上下包迹线之间,符合随机分布;若在上包迹线以外,则呈聚集分布;若在下包迹线以外,则呈均匀分布[16]。采用先决条件零假设模型,进行双变量空间关联分析,依据Monte Carlo模拟结果,如g(r)在上下包迹线之间,表明两者空间上相互独立;如 g (r)在上包迹线以上,则两者空间上为正相关;如在下包迹线以下,则两者空间上负相关。
数据分析使用Programita软件完成,通过99次Monte Carlo模拟,得到99%的置信区间。数据处理采用Excel以及Origin软件完成。
2 结果与分析
2.1 径级结构分布特征
由图1表明:从第Ⅰ径级到第Ⅷ径级,辽东栎各径级所占比例逐渐减小,依次为46.48%、24.12%、13.91%、10.92%、3.36%、0.88%、0.18%、0.18%,从第Ⅲ到第Ⅵ径级所占比例减少,速度减慢。种群径级结构分布比例整体上为倒J型。
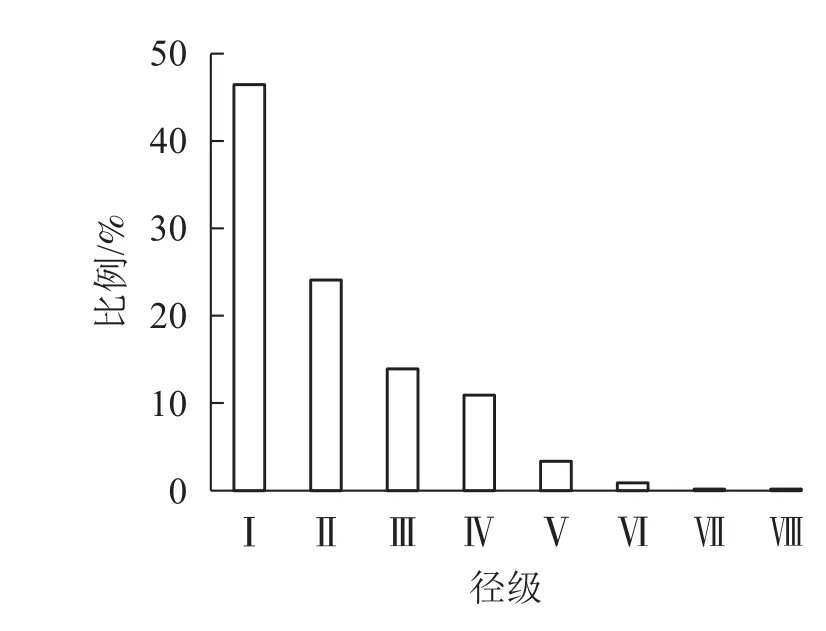
图1 辽东栎径级结构的分布Figure 1 Distribution of diameter class structure of Q. wutaishanica
2.2 静态生命表
从表2可见:辽东栎种群的个体数量随径级增大而减少,同时标准化存活个体数(lx)也随径级增大而减少。辽东栎种群期望寿命(Ex)在第Ⅱ径级最高,为1.72;而在第Ⅰ径级个体数量最大,在第Ⅵ到第Ⅶ径级,Ex又开始增大至1.50。在第Ⅷ径级,Ex突然降到0.50,说明这时辽东栎种群已进入衰退阶段。辽东栎种群的存活率(Sx)在第Ⅲ径级达到最高,而在第Ⅶ径级又达到存活高峰。
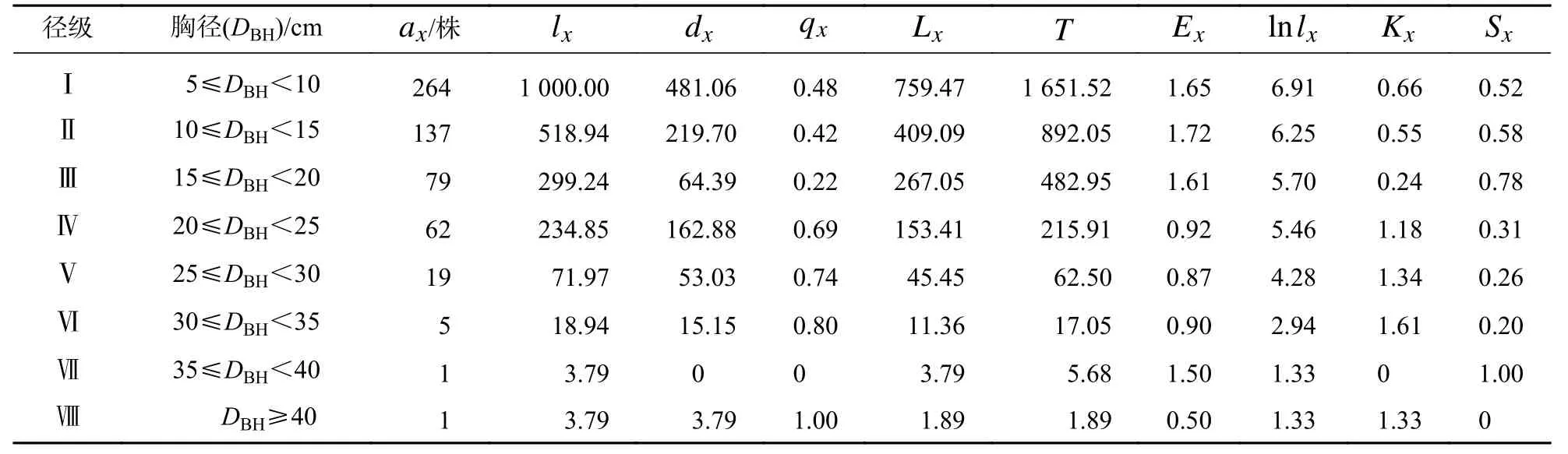
表2 辽东栎种群静态生命表Table 2 Static life table of Q. wutaishanica population
2.3 存活系数
根据Deevey的划分,存活曲线被分为3种类型:Deevey-Ⅰ型、Deevey-Ⅱ型、Deevey-Ⅲ型[17]。从图2可看出:六盘山辽东栎存活趋势线趋于Deevey-Ⅱ型(斜线型),因第Ⅰ到第Ⅳ径级的存活系数逐渐下降,第Ⅳ第Ⅶ径级的存活系数近似相等,所以辽东栎种群存活趋势线在径级Ⅶ前大致呈直线。在第Ⅶ径级,存活系数突然下降,仅有33.36%的第Ⅵ径级的辽东栎个体可通过此筛选进入第Ⅶ径级。
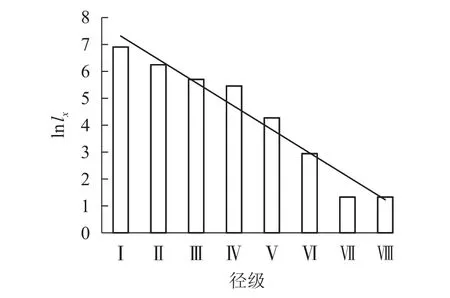
图2 各径级辽东栎的存活系数(ln lx)Figure 2 Survival coefficient (ln lx) of Q. wutaishanica under different diameter classes
2.4 死亡率和消失率
由图3可看出:种群的死亡率和消失率总体上均随径级增加先减后增,变化趋势一致,两者仅在第Ⅲ和Ⅶ径级出现波动,呈下降趋势,第Ⅶ径级死亡率和消失率突然降至为0。均出现了2个最大值,第1个最大值在第Ⅵ径级,这时死亡率为0.80,由于此阶段种内竞争加剧,环境压力下消失率达到顶峰,为1.61;第2个最大值出现在第Ⅷ径级。
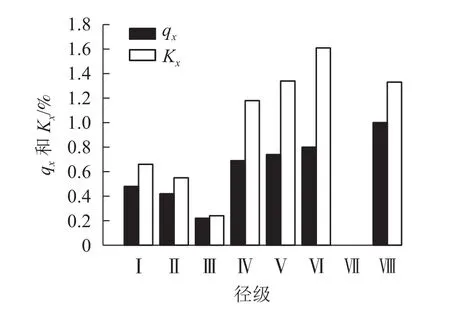
图3 各径级辽东栎的死亡率(qx)和消失率(Kx)Figure 3 Mortality (qx) and disappearance rate (Kx) of Q. wutaishanica under different diameter classes
2.5 生存分析
由图4可看出:在第Ⅰ到第Ⅵ径级阶段,随龄级增大,辽东栎种群的生存率逐渐减小,而积累死亡率逐步增大;在第Ⅵ到第Ⅷ径级阶段,生存率与积累死亡率呈平稳状态。

图4 辽东栎种群的生存率[S(x)]与积累死亡率[F(x)]Figure 4 Survival [S(x)]and accumulated mortality [F(x)]of Q.wutaishanica population
由图5可看出:辽东栎种群的死亡密度在第Ⅲ径级出现第1个峰值,在第Ⅶ到第Ⅷ径级阶段,死亡密度达到最高并不再变化。辽东栎种群的危险率在第Ⅷ径级达到最大值,此时的死亡密度也最大。
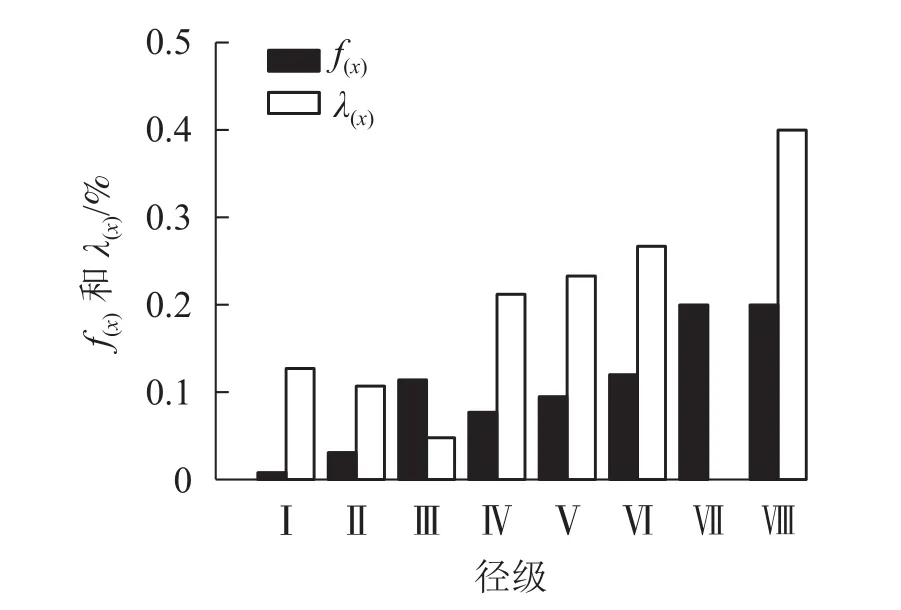
图5 辽东栎种群的死亡密度[f(x)]和危险率[λ(x)]Figure 5 Death density [f(x)]and risk rate [λ(x)]of Q. wutaishanica population
2.6 辽东栎种群的点格局
从12个样地中选择有代表意义的3个典型样地(第9、10、11号样地),把辽东栎分为4个生长阶段:幼龄阶段(5 cm≤DBH<10 cm)、中龄阶段(10 cm≤DBH<20 cm)、近熟龄阶段(20 cm≤DBH<30 cm)、成过熟龄阶段(DBH≥30 cm)。因为样地中辽东栎处于成过熟龄阶段的个体数特别少,因此本研究只考虑前3个生长阶段,从图6可以看出:典型样地9的辽东栎个体数从大到小依次为幼龄树、近熟龄树、中龄树,多为聚集分布;典型样地10的个体数从大到小依次为幼龄树、中龄树、近熟龄树,各生长阶段个体分布均匀;典型样地11的中龄树最多,个体呈随机分布。
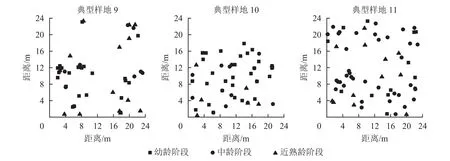
图6 典型样地位置空间分布图Figure 6 Location and spatial distribution of typical plots
取尺度r的间隔为1~14 m,对3个典型样地的幼龄、中龄、近熟龄辽东栎生长阶段的点格局进行了分析。由图7看出:典型样地9中,幼龄阶段在0~1 m和9~10 m尺度上为聚集分布,在其他尺度上为随机分布;中龄、近熟龄个体大致都为随机分布。典型样地10的幼龄和中龄个体在小尺度上为聚集分布,随着尺度变大,幼龄个体呈随机性聚集分布;近熟龄阶段在小尺度上出现均匀分布趋向,中龄个体在尺度14 m以内为随机分布。样地9和10内胸径大于20 cm的近熟龄个体,在尺度1~14 m都为随机分布。典型样地11内所有个体在1~14 m都为随机分布,在0~10 m上都呈聚集分布。
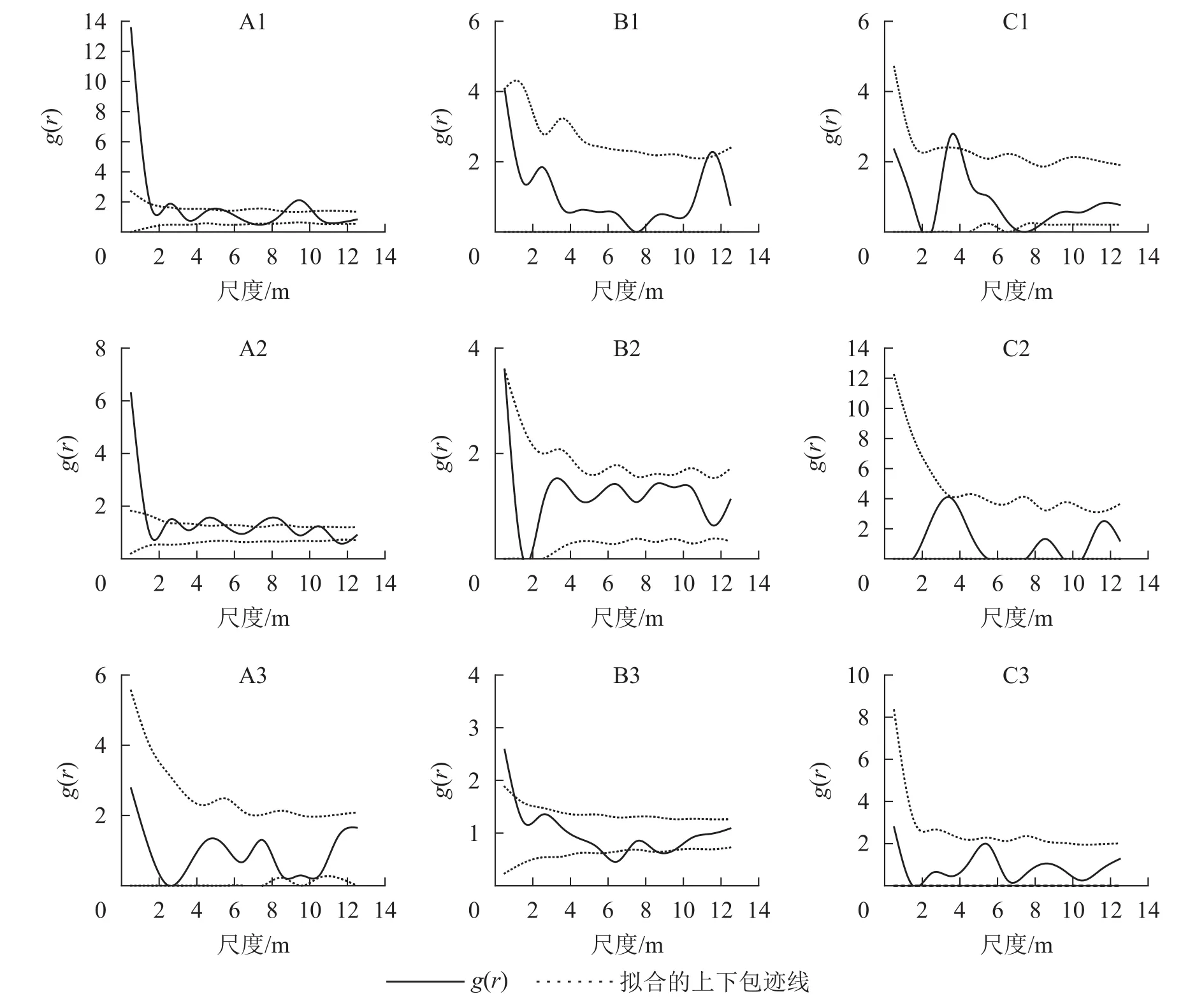
图7 典型样地各龄级点格局Figure 7 Patterns of point at each age level of typical plots
对各典型样地各年龄阶段进行双因素分析(图8)表明:典型样地9的幼龄与中龄个体间在小尺度上呈正相关,幼龄与近熟龄个体在尺度5~10 m呈负相关,中龄和近熟龄个体在0~14 m相关不显著。典型样地10的幼龄与中龄个体在2~6 m尺度上呈正相关,幼龄与近熟龄个体在0~14 m尺度上无相关,中龄和近熟龄个体之间无相关。典型样地11的辽东栎各生长阶段之间在0~2 m尺度上呈负相关,随尺度增大的相关性越来越不明显,呈相互独立状态。
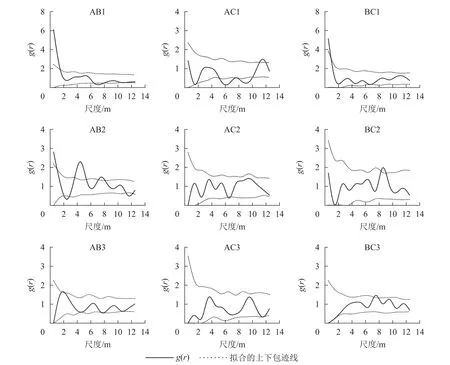
图8 典型样地各龄级间空间关联性Figure 8 Spatial correlation among age levels of typical plots
3 结论与讨论
种群结构是物种自身生物学特征、环境因素及植物种内和种间关系综合作用的结果[11]。静态生命表和生存分析可准确地反映种群在特定时间和空间范围内的动态规律[12]。种群的分布格局是由植物自身的生物学特性和环境因子共同决定的,可表明种群变化特征和群落的发展走向[18-19]。以往的辽东栎种群点格局研究[17,20]认为:辽东栎种群中幼树主要呈聚集分布,随生长发育趋向于随机分布。
本研究得出:辽东栎种群在前期时,死亡率随龄级增加而减小,这是因为种群顺利度过了幼苗到幼树的瓶颈期,使辽东栎个体对环境的适应能力不断加强;在中期时,死亡率随龄级增加而升高,并趋于稳定,辽东栎种群的期望寿命在第Ⅱ径级达到最高,而第Ⅰ径级的个体数量最大;原因可能是第Ⅰ径级的辽东栎种群竞争强度大,个体间竞争增强,产生“自疏”,为第Ⅱ龄级的辽东栎生长提供了更多水热资源[21],使辽东栎在此期间生长旺盛,种群存活率最高。在第Ⅵ到第Ⅶ径级,期望寿命又开始增大,表明这个阶段辽东栎种群竞争强度逐渐减小。在第Ⅷ径级,期望寿命突然下降,表明辽东栎种群已进入衰退阶段。本研究生存分析发现:辽东栎群落生存曲线基本属于Deevey-Ⅱ型,这是因随径级增大,生存能力逐渐下降。辽东栎是地带性植被,各地的辽东栎并没固定的生存线型,凸线型[22]和凹线型[23]在以往研究中都存在。六盘山地区的辽东栎种群具有前期和中期稳定、后期衰退的生存过程。
本研究表明:在辽东栎种群中,幼龄和中龄个体在小尺度范围内呈聚集分布,随尺度不断增大,由聚集分布转为随机分布,这与伊力塔等[3]、王道亮等[7]的研究结论大致相同。在0~14 m尺度内,辽东栎种群中的幼树在小尺度上主要呈聚集分布,这是因辽东栎常出现幼苗簇生群落,且林内生态环境有利于幼苗发育[24]和辽东栎萌蘖更新、桩蘖更新。中龄树或近熟龄树占辽东栎种群较大比例时呈随机分布,这是因为随着径级增大,种群个体因争夺光照、土壤养分和水分等资源,出现恶性竞争,个体数量减少,聚集程度不断减弱,所以逐渐表现为随机分布格局。对3个典型样地的幼、中、近熟龄个体进行的双因素分析表明:小尺度上,样地9和10的幼龄和中、近熟龄个体都有一定正相关,这是因为在样地9和10的辽东栎个体之间存在良好竞争;大尺度上样地9、10、11的辽东栎个体之间大致都为无明显相关,这与王道亮等[7]的研究结果一致。由于种群空间格局是由个体生物学特性和环境条件相互作用所决定的[6],研究尺度也会影响研究结果,在未来的研究中可以扩大样地面积范围,进行六盘山辽东栎纯林的种群格局分析。
本研究认为:六盘山辽东栎林乔木种群处于相对稳定状态,更新状态良好,为保持辽东栎纯林结构稳定,应加强人为保护,维持辽东栎林种群更新的稳定环境条件,提高第Ⅱ径级的存活率。同时加强人工正面干预,使种群个体间保持良性竞争。
猜你喜欢
农业科技通讯(2023年1期)2023-02-12 07:10:26
广西林业科学(2022年4期)2022-11-07 01:50:34
热带作物学报(2022年9期)2022-10-17 05:49:06
林业调查规划(2022年3期)2022-06-13 09:31:04
广西林业科学(2021年6期)2022-01-21 07:39:10
老年教育(2018年12期)2018-12-29 12:43:56
云南档案(2016年11期)2016-12-13 07:15:22
中国果菜(2016年9期)2016-03-01 01:28:45
西北植物学报(2015年9期)2015-07-05 07:03:42
长江蔬菜(2013年23期)2013-03-11 15:09:46
