
植物响应非生物胁迫的分子机制
2021-12-20 10:53陈柯岐邓星光林宏辉
生物学杂志 2021年6期
陈柯岐, 邓星光, 林宏辉
(四川大学 生命科学学院 生物资源与生态环境教育部重点实验室,成都 610065)
非生物胁迫严重危害农业生产,同时导致生态环境极度恶化。植物遭受逆境胁迫后,发生一系列从形态到分子水平的变化,这些变化直接或间接影响植物的生长和发育。极端温度、干旱、水淹和盐害等逆境往往相互关联,在造成特殊影响的同时,也造成相似的细胞损伤。为了生存,植物在遭受非生物胁迫时需要在形态和生理生化代谢上进行一系列调整,以适应逆境。植物响应非生物胁迫一般从信号感知开始。细胞外膜受到非生物胁迫的刺激后,产生第二信号分子,如活性氧(ROS)和肌醇磷脂等。第二信号分子刺激细胞内膜,通过调节胞内Ca2+水平,启动蛋白磷酸化级联反应,产生磷酸化蛋白分子,以直接参与细胞保护的蛋白或调控特异性胁迫调节基因的转录因子为目标。有些基因的产物参与产生脱落酸(ABA)和乙烯(ETH)等调节因子,从而激活转录因子的表达。转录因子与靶基因启动子序列特异性结合,进一步激活或抑制下游功能基因的表达,最终起到对非生物胁迫的调控作用。揭示植物非生物胁迫响应机制,培育抗逆作物新品种,有助于实现农业的可持续发展,并保障日益增长的世界人口的粮食安全[1]。本文结合极端温度、干旱、水淹和盐害对植物生长发育的影响,从信号感知、信号转导、基因表达调控和重要次生代谢产物变化等方面,综述植物响应非生物胁迫的分子机制,展望未来的研究方向。
1 植物温度胁迫响应机制
随着全球范围内异常天气发生规模扩大和频率增加,各种植区作物生长和发育都受到影响。在极端温度下,作物的产量和品质显著降低,种植地点也被严重限制。因此,要想在当前气候变化的情况下维持农业可持续发展,了解植物温度胁迫响应机制至关重要[2]。
1.1 低温
低温主要通过细胞质脱水和细胞壁结冰引发细胞死亡。温带地区的植物暴露在不结冰的低温环境中,会变得更加耐寒,该过程被称为冷驯化。冷驯化过程中会发生许多生理、生化和分子变化,包括抗氧化系统的激活、低温保护剂的合成和积累以及保护和稳定细胞膜的变化[3]。为了保持细胞膜在低温胁迫下的稳定性,膜脂组分中不饱和磷脂的含量上升。同时,细胞积累渗透分子和抗冻蛋白,这些蛋白富含蔗糖和脯氨酸,它们通过产生氢键来捕获水分子。
1.1.1 信号感知
膜流动性的变化与温度传感有关。冷、热胁迫改变细胞膜的流动性,影响膜定位蛋白的结构和活性,从而触发Ca2+内流,该过程对诱导温度响应性基因的表达具有重要意义[4]。
有研究证实,水稻低温感受器(COLD1)与水稻G蛋白α亚基1(RGA1)结合,介导水稻的冷感知和低温诱导的胞外Ca2+内流[5]。除了COLD1,水稻CBL互作蛋白激酶7(OsCIPK7)被认为通过调节其自身激酶结构域的构象和Ca2+内流来感知低温信号[6]。然而,目前还不清楚COLD1和OsCIPK7如何在低温胁迫下调控Ca2+内流。拟南芥基因组包含40多个编码Ca2+通道的基因,其中许多都定位于质膜。环核苷酸门控离子通道(CNGCs)作为植物生长、发育和胁迫响应的重要调节因子[7],为拟南芥和苔藓等提供耐热性[8]。然而,目前的研究还不清楚CNGCs是否具有在低温下感知Ca2+通道的功能。
1.1.2 信号转导和基因表达调控
丝裂原活化蛋白激酶(MAPK)级联参与调控冷信号和低温耐受性。植物受冷胁迫后,MKK2磷酸化并激活MPK4和MPK6。在MKK2过表达的植物中,C-重复结合因子2/脱水应答元件结合蛋白1C(CBF2/DREB1C)的表达水平上调。因此,低温信号通路可能通过蛋白磷酸化而被激活[9]。
拟南芥中的3个CBF/DREB1基因在冷驯化中发挥着核心作用。在冷驯化过程中,CBF2/DREB1C负调控CBF1/DREB1B和CBF3/DREB1A的表达,而CBF1/DREB1B和CBF3/DREB1A没有参与调节其他CBF/DREB1基因[10]。此外,通过RNAi降低CBF1和CBF3转录本的植物表现出较低的冷驯化能力,这表明CBF1/DREB1B和CBF3/DREB1A协同诱导CBF/DREB1-调控基因和冷驯化[10]。CBF基因受到低温的诱导,激活低温调节基因(COR)的表达[11]。CBF/DREB1A的过表达可以诱导COR基因的表达和拟南芥、番茄和水稻等的低温耐受性[12]。
CBF基因表达受多种类型转录因子的调控。拟南芥中CBF蛋白的稳定性受到蛋白质翻译后修饰(PTMs)的调节。CBF表达诱导因子1(ICE1)及其同系物ICE2正向调节CBF表达和低温耐受性。泛素化、SUMO化和磷酸化对ICE1在拟南芥中的功能发挥起着重要的作用[13]。MYB15转录因子与CBF/DREB1启动子中的MYB顺式作用元件结合,负向调控后者的表达[14]。低温诱导的C2H2锌指转录因子(ZAT12)也是CBF/DREB1的负调控因子。有研究表明,CBF/DREB1的表达在ZAT12过表达植株中降低[15](图1)。

箭头代表促进作用;双竖线代表抑制作用。图1 植物响应冷胁迫的简化模型Figure 1 Simplified model of plant in response to cold stress
1.2 高温
全球变暖导致包括极端高温在内的气候变化,这些变化对作物生产造成毁灭性的破坏。当高温胁迫发生在发育的关键阶段时(生殖期、开花期等),对植物的生存非常不利:高温会抑制花粉活力、受精能力、籽粒灌浆和果实形成[16]。同时,高温胁迫会抑制光合作用,由高温引起的水分含量降低也会对植物细胞分裂和生长造成负面影响[17]。
1.2.1 信号感知和信号转导
最近的研究表明,Ca2+对热胁迫响应(HSR)的触发不可或缺。Ca2+进入细胞质以响应热胁迫引起的质膜流态化。Ca2+渗入由CNGC介导。到目前为止,CNGC2、CNGC4、CNGC6和CNGC16被证实为与HSR相关的CNGCs[18]。在热胁迫过程中,环腺苷酸(cAMP)和环鸟苷酸(cGMP)的水平升高,CNGCs在这些环核苷酸的作用下打开[19]。因此,有研究人员提出,质膜流态化可以通过激活核苷酸环化酶打开CNGCs。
ROS也被认为是热胁迫信号传感器。ROS的积累是触发HSR的必要条件。在热胁迫过程中,叶绿体是产生ROS的主要场所。此外,NADPH氧化酶、呼吸爆发氧化酶同系物B(RbohB)和RbohD也能产生ROS。虽然目前还不清楚ROS信号如何被感知和转录调控,但ROS信号已被证实可触发与一氧化氮(NO)和活性短链叶挥发物(RSLVs)相关的两条信号通路。遗传分析表明,RbohB和RbohD产生的ROS可引起NO的积累[20]。
钙依赖蛋白激酶(CDPK)级联参与调控热信号和高温耐受性。高等植物主要有3类Ca2+结合蛋白:CDPK、钙调蛋白(CaM)和类CaM蛋白,以及类钙调神经磷酸酶B[21]。CDPKs通过Ca2+与类钙调蛋白结构域的结合直接激活,进一步调节下游靶位点。在热胁迫下,CaM3充当Ca2+信号转换器[22]。NO信号激活CaM3,导致热激转录因子(HSF)与DNA结合[23],进而激活下游热胁迫相关基因的表达。
1.2.2 基因表达调控
HSF家族成员HsfA1通过激活HSR基因从而调控植物对热胁迫的耐受性。在热胁迫下,HsfA1的表达上调,而HsfA1的活性恰恰受高温的调控[24]。热激蛋白(HSP)HSP70和HSP90在一定温度范围内抑制HsfA1的活性和核定位,而热胁迫诱导HsfA1从HSP70、HSP90复合体中分离,进而激活HsfA1[25]。
研究人员推测HsfA1s直接调节编码重要热胁迫应答转录因子基因的表达,包括HsfA3、DREB2C、HsfA2、HsfBs和多蛋白桥联因子1C(MBF1C)等[26]。HsfA3是一种重要的热胁迫反应因子,HsfA3的缺失会导致热胁迫过程中靶基因表达下调[27]。DREB2C(DREB2A同源基因)的过表达能激活HsfA3,该过程在没有热胁迫时也能实现。因此,DREB2C在热胁迫过程中参与调节HsfA3基因[28]。HsfA2是HsfA1的直接靶基因,在植物中对HSR起关键作用。研究人员发现,hsfa2敲除突变体对热胁迫具有高敏感性,许多热胁迫诱导基因的表达减少[29]。HsfBs(HsfB1和HsfB2b)也是拟南芥中HsfA1的下游靶基因。热胁迫诱导的MBF1C表达受HsfA1调控。有研究表明,在热胁迫期间,hsfa1缺失突变体中MBF1C的表达受到抑制[26]。
一些HSF家族蛋白可以独立于HsfA1途径参与HSR。HsfA4a受氧化应激(包括热胁迫)诱导,调控抗坏血酸盐过氧化物酶(APX)的表达。HsfA9在种子中特异性表达,其过表达可诱导多种HSP基因。bZIP28也是一个重要的转录因子,它独立于HsfA1途径调控HSR[30]。
1.2.3 调节错误折叠蛋白和ROS稳态
热胁迫下产生的错误折叠蛋白和过量ROS对植物有毒害作用。因此,植物必须对这些错误折叠蛋白进行复性或降解,并清除ROS以减轻细胞损伤。在拟南芥和水稻中,HSPs由HSFs诱导,它们编码的蛋白作为分子伴侣,使错误折叠蛋白重新复性,该过程在植物对热胁迫的耐受性中发挥了重要作用。在拟南芥中,HSP100通过分解蛋白聚集体来增强耐热性[31]。除了HSPs,26S蛋白酶体α2亚蛋白OsTT1通过清理热诱导的细胞毒性变性蛋白,增强水稻的耐热性[32]。
热胁迫还会激活抗氧化系统,以清除多余的ROS。负责清除ROS的APX和过氧化氢酶(CAT)突变体对热胁迫敏感[33]。其他重要的ROS清除剂,如铜/锌超氧化物歧化酶1(CSD1)、CSD2和SOD铜伴侣1(CCS1),对拟南芥的耐热性也非常重要[34](图2)。
2 植物水分胁迫响应机制
干旱是对农业影响最大的非生物胁迫之一,与其他非生物胁迫相比,干旱严重损害植物生长发育,限制作物生产。目前,农业生产消耗了全世界70%以上的淡水。随着全球气候变暖,这一消耗量估计还会增加。近年来的全球水灾发生频率逐渐增高,该趋势与气候变化相关[35]。洪水引发水淹胁迫,对植物多样性、物种分布及粮食生产构成严重威胁。
2.1 干旱
干旱胁迫诱导植物产生一系列生理生化反应,包括气孔关闭、光合抑制和呼吸激活。植物也会在细胞和分子水平上对干旱做出一系列响应,如积累渗透调节物质和特异性胁迫耐受蛋白。期间,干旱胁迫诱导或抑制各种不同功能基因的表达。干旱诱导ABA的合成,而ABA导致气孔关闭,并激活胁迫相关基因的表达。已有证据表明,在调控干旱耐受基因表达过程中,存在ABA依赖性和ABA非依赖性信号系统[36]。

箭头代表促进作用;双竖线代表抑制作用。图2 植物响应热胁迫的简化模型Figure 2 Simplified model of plant in response to heat stress
2.1.1 ABA依赖性途径
ABRE是ABA响应基因表达的主要顺式作用元件,bZIP转录因子ABRE结合蛋白/ABRE结合因子(AREB/ABF)与ABRE结合,从而激活ABA依赖性基因表达。在拟南芥中,过表达ABF3或AREB2/ABF4引起植株对ABA超敏,蒸腾速率降低,并增强耐旱性[37]。干旱诱导基因RD22的激活由ABA介导,MYC转录因子AtMYC2(RD22BP1)和MYB转录因子AtMYB2被证实可以结合RD22启动子的顺式作用元件并一起激活RD22的表达[38]。这些MYC及MYB蛋白在内源ABA积累后被激活,进一步发挥在后期胁迫反应中的作用。
研究人员鉴定出干旱诱导编码NAC转录因子的RD26基因[39]。干旱、高盐、ABA和茉莉酸(JA)处理均可诱导RD26的表达。RD26过表达的植株对ABA敏感,热胁迫诱导基因的表达上调;而RD26功能被抑制后植株则表现出相反的表型。
2.1.2 ABA非依赖性途径
一些干旱诱导基因不受ABA信号的调控,这说明植物存在不依赖于ABA的信号通路来响应干旱胁迫。其中,ERD1是一个重要的ABA非依赖性基因。该基因不仅受脱水诱导,而且在自然衰老和黑暗诱导的衰老过程中表达上调。对转基因植物ERD1启动子的分析表明,ERD1启动子中含有顺式作用元件,不仅参与了依赖胁迫响应的基因表达,还参与了衰老诱导的基因表达[40]。
低温、干旱和高盐诱导基因LTI78、RD29A和COR78的启动子包含两个主要的顺式作用元件,即ABRE和脱水应答元件/C-重复(DRE/CRT),它们也参与了胁迫诱导的基因表达。与DRE/CRT结合的ERF/AP2家族转录因子被分离并命名为CBF/DREB1和DREB2。许多CBF/DREB1靶基因已经通过cDNA和基因芯片微阵列技术被鉴定出来[41]。其中,大多数CBF/DREB1靶基因在其启动子区域包含具有保守(A/G)CCGACNT序列的DRE序列。这些蛋白的靶基因产物参与胁迫耐受性的建立。DREB2基因受脱水胁迫诱导,可能激活其他与干旱胁迫耐受性相关的基因[42](图3)。
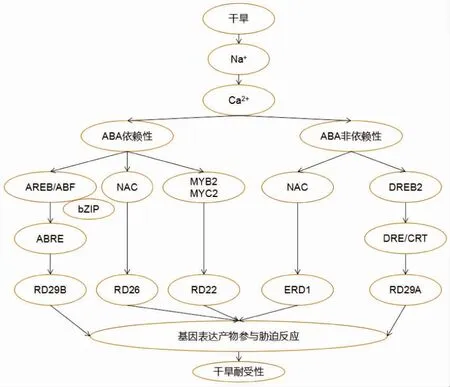
箭头代表促进作用。图3 植物响应干旱胁迫的简化模型Figure 3 Simplified model of plant in response to drought stress
2.2 水淹
氧气在水中的扩散比在空气中慢104倍。因此,对适应能力较差的植物物种来说,水淹胁迫会导致淹没在水里的组织氧气供应不足。除了缺氧的威胁,过多的水分还会导致土壤和植物的其他变化:土壤微生物厌氧代谢的产物积累,乙烯的水平上升。此外,洪水导致完全水淹以及通常被水淹的水生植物中,二氧化碳、光和氧气的获取受限[43]。
2.2.1 通气组织
植物通过改善与环境的气体交换避免体内缺氧,为此,植物发育出通气组织,形成一种纵向连接的气体空间,为植物体内长距离气体交换提供了一条捷径[44]。通气组织通常与根系结构改变相结合,以减少空气表面和被水淹根尖之间的距离,从而减少扩散阻力[45]。例如,通过不定根产生一组从下胚轴或茎进入厌氧基质的导气通气管。通气组织与防止氧气泄漏到周围厌氧土壤的屏障结合,极大地提高了植物对水淹的耐受性[46]。
通气组织系统能使植物有效地适应水淹胁迫,在水淹条件下,地上组织保持与大气接触,可以将空气输送到地下组织。然而,在完全水淹的过程中,地上部分不能与大气接触,它们将空气输送到地下部的效率降低。在这种情况下,为了恢复与大气的接触,一些湿地植物表现出叶片、节间或叶柄的快速垂直伸长,以便浮潜换气[47]。
2.2.2 细胞内高浓度乙烯的感知
在水淹胁迫下,植物体内乙烯含量快速上升,诱导植株形态发生改变。作为气体,乙烯在水淹条件下很难脱离植物,导致其在体内快速积累。因此,乙烯是检测植物是否处于水淹胁迫的可靠指标[48]。
乙烯通过调节ABA、赤霉素(GA)和生长素(IAA)的积累,诱导一些植物形成不定根。然而,根的形成也需要乙烯诱导表皮细胞产生ROS,促使细胞死亡,从而使根的通透性增强。同样,溶生性通气组织由皮层中特定细胞的凋亡形成,涉及乙烯依赖性的抗氧化活性下降,随后,ROS增加导致细胞死亡[49]。
在水淹胁迫下,高浓度乙烯抑制根的伸长,有些植物通过形成通气组织,很轻松地清除过量乙烯。然而,无法产生通气组织的植物物种根系生长受到显著抑制。在这些植物不同的发育过程中,对高浓度乙烯信号的依赖可能发挥重要作用。为了避免与高浓度乙烯有关的不利影响,一些持续占据水生环境或易受洪水影响的植物物种已经丧失或降低了它们产生、感知和响应乙烯的能力[50]。
2.2.3 细胞内低浓度氧的感知
植物在水淹胁迫下,氧含量的改变是一种重要的感知信号。低氧诱导的胁迫不仅发生在水淹条件下,还会在具有快速代谢特征的组织中出现,如根分裂组织和韧皮组织等。虽然植物通过光合作用产生氧气,但是缺乏有效的系统将氧气运输到非光合器官,如果一些器官的解剖结构限制了氧气的扩散,这些器官可能会处于低氧状态[51]。当氧气限制呼吸时,植物会出现缺氧,而完全无氧对植物的生存更不利。缺氧和完全无氧都能触发基因的大量重编程,并诱导发酵代谢,从而使植物利用糖酵解产生ATP。
虽然拟南芥对水淹的耐受性不是很高,但它作为模式植物的特性,使氧传感和信号转导的发现成为可能[52]。除了已知的厌氧基因外,还鉴定出几种缺氧应答未知蛋白(HUP),它们可能代表新的厌氧反应途径。VII类乙烯反应因子(ERF-VIIs)在水稻中的作用促进了在拟南芥中对该基因家族的研究[53]。RAP2.12是一组ERF-VIIs,它并非由缺氧诱导,但能激活乙醇脱氢酶(ADH)[54]。RAP2.12在蛋白水平上受氧气调控,氧气促进其降解。只有在低氧条件下,RAP2.12和其他两个组成性表达的ERF-VIIs,即RAP2.2和RAP2.3,才能稳定地激活厌氧反应[55]。
缺氧反应衰减器1(HRA1)是一种三螺旋转录因子,它能抑制RAP2.12的表达。在缺氧条件下,HRA1基因表达本身被RAP2.12稳定激活,表明厌氧反应的调节存在一种动态平衡机制[56]。最近的研究鉴定出一种连接氧感应机制和ROS产生的蛋白。低氧应答通用应激蛋白1(HRU1)由氧应答型N端规则通路诱导,可能通过与膜定位的RbohD及其调控因子ROP2的相互作用影响ROS的合成[57]。这些发现表明,缺氧依赖性信号通过一个紧密连接的网络,被各种信号和蛋白严格控制。
3 植物盐胁迫响应机制
土壤盐渍化是限制土地使用和作物生产的全球性问题之一,据估计,世界上大约五分之一的灌溉土地受到盐害的影响[58]。此外,还有大面积的边缘土地,如沿海地区,不适合种植作物。利用这些盐碱地生产作物可以缓解人口不断增长带来的粮食需求问题。
盐胁迫显著抑制植物的种子萌发、根长、株高和结实[59]。在盐渍土壤中,渗透胁迫是植物最早经历的胁迫,会立即影响植物的生长。当盐浓度达到一个阈值,植物会遭受离子毒害,超过这个阈值,植物便不能维持离子稳态和正常生长。离子毒害和渗透胁迫是引起氧化胁迫和一系列次生胁迫的主要原因。
3.1 信号感知和信号转导
越来越多的证据表明,植物细胞壁参与盐胁迫的感知和响应。植物细胞壁不仅保护细胞免受生物和非生物胁迫,还是第一个感知和响应逆境胁迫的细胞结构[60]。活性细胞壁完整性(CWI)的维持和感知对应激保护和胁迫响应至关重要。除了CWI状态的感知外,细胞壁调节的相关过程也参与了对盐胁迫的反应。木质素含量和组成在生物和非生物胁迫中会发生变化[61]。木糖半乳糖转移酶参与细胞壁的生物合成和内膜组织的维持,通过限制细胞内ROS的浓度来减少细胞损伤,并调节盐胁迫下应激相关基因的表达[62]。因此,研究人员推测出一种可能的盐敏感机制:在盐胁迫下,细胞壁的损伤和成分变化触发细胞壁维持和CWI感知机制,进而激活植物盐胁迫的响应和回避机制,调控根的生长和结构改变[63]。
维持细胞内离子稳态是耐盐植物响应过量离子的重要适应性特征。通过减少细胞质中Na+并增加K+,可以使植物获得合适的细胞质K+、Na+浓度比,从而防止细胞损伤和营养缺乏。减少细胞质中Na+的机制包括限制Na+摄取、促进Na+外排和液泡中分隔Na+。基于对盐超敏感基因(SOS)的发现,研究人员建立了一种离子胁迫信号转导途径。SOS1是Na+细胞质-质外体运输的关键元件。SOS2和SOS3能调节SOS1的表达。在盐胁迫下,SOS3-SOS2复合物正向调节液泡Na+/H+交换器NHX和H+-ATPase的活性[64]。NHX是一种潜在的Na+/H+交换器,它将Na+从细胞质运输至液泡,该过程由液泡H+-焦磷酸酶和H+-ATPase建立的H+梯度驱动[65]。
3.2 活性氧清除
氧气作为植物生存不可缺少的物质之一,参与新陈代谢、线粒体呼吸和氧化磷酸化以产生能量。然而,在代谢过程中,氧气被激活为ROS。ROS具有较强的氧化能力,可引起细胞质膜损伤、不可逆代谢功能障碍和细胞死亡。
超氧化物歧化酶(SOD)是植物抗氧化酶系统的第一道防线,能清除细胞内多余的超氧化物阴离子。SOD能使O2歧化形成H2O2,消除超氧阴离子的毒性[66]。在高盐胁迫下,植物中ROS的增加导致细胞膜中脂质的过氧化。丙二醛(MDA)是植物膜脂过氧化的主要产物,其含量的变化能反映细胞膜受损的程度。因此,可以利用MDA的含量指示植物的盐胁迫和耐盐性[66](图4)。
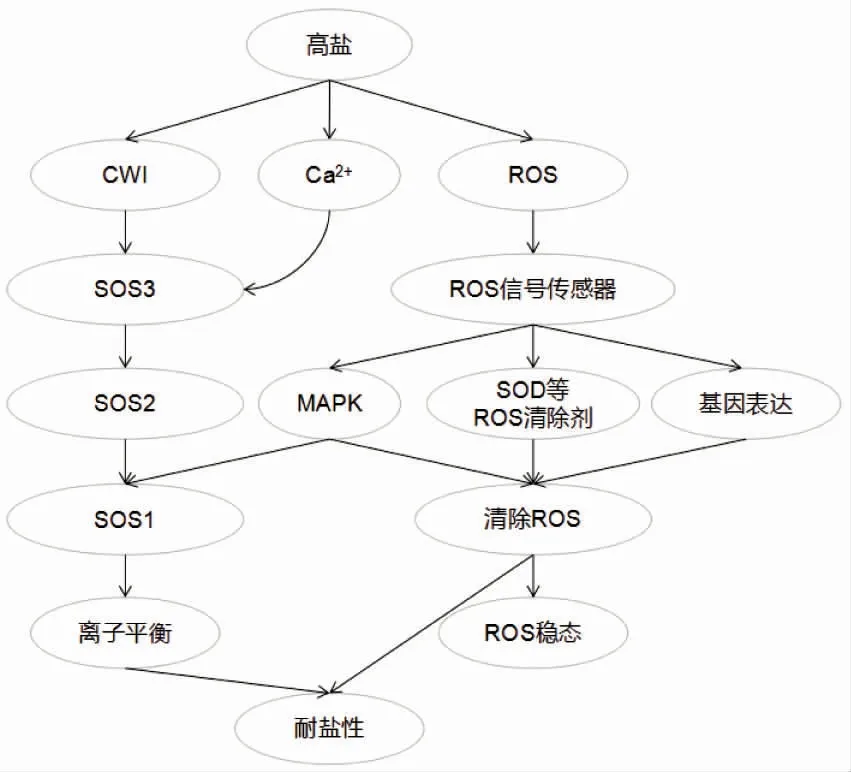
箭头代表促进作用。图4 植物响应盐胁迫的简化模型Figure 4 Simplified model of plant in response to salt stress
4 总结与展望
目前,植物响应非生物胁迫的分子机制已经逐渐清晰(图5)。近年来,研究者对主要非生物胁迫应答过程中的信号转导和基因表达调控途径开展大量的研究,许多激素和转录因子在植物抗逆中的作用也被揭示。但现阶段的研究进展大多以未知转录因子的发现、基因结合位点的完善和生物化学水平的细节为主,新的研究方向、研究工具和研究实践仍有待开发。
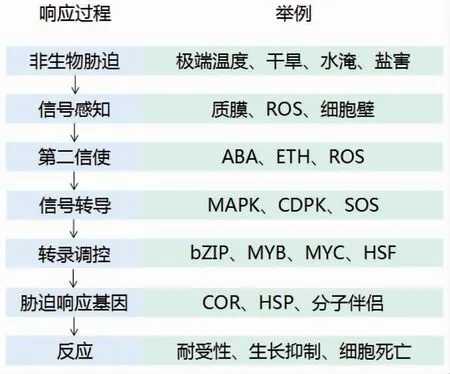
图5 植物非生物胁迫响应过程及举例Figure 5 The response process of abiotic stress in plants and examples
多数胁迫信号的感知都与Ca2+内流有关,但具体哪些调控因子参与感应Ca2+,以及这些调控因子的调节方式还不够清楚;虽然有研究表明,质膜、ROS和细胞壁等能作为胁迫信号的传感器,对它们的研究大多还停留在生物化学水平和未知蛋白方面。这些传感器对信号的具体感知方式以及信号感知后,传感器下游的信号通路还没有明确。在信号转导方面,MAPK、CDPK和SOS途径已经趋于完善,尤其是MAP、CDPKs和SOS这些激酶的连续激活过程和蛋白修饰方式;研究人员利用基因功能缺失或过表达植株,对大多数激酶的正/负向调控作用有了深刻认识;同时,大量研究表明,ABA等植物激素在信号转导过程中发挥非常重要的作用。然而,迄今为止的试验仅集中在ABA和ETH中,研究人员对水杨酸(SA)和JA等的关注较少。虽然有证据表明,SA和JA等也参与信号传递,而且它们可能有独立的调控途径,但具体的机制还不了解。利用反向遗传学手段,对非生物胁迫转录调控调节因子的探究非常有效,目前的研究已经鉴定了大量的转录因子,它们与靶蛋白启动子的顺式作用元件结合,激活特异基因的表达。研究人员在拟南芥和水稻等模式植物中,对这些转录因子的结合位点及其与靶基因启动子的结合方式作出了详细的解析。然而,涉及多个转录因子协同激活抗逆信号,仍有很大的研究空间。总的来说,想要揭示植物非生物胁迫响应的机理,在分子水平上还需综合利用基因组学、转录组学和蛋白质组学等研究方法。在细胞学层面,随着显微镜技术的快速发展,利用冷冻透射电镜、高分辨率荧光显微镜等,观察植物细胞膜、细胞器和细胞骨架的变化,有助于更好地理清和解析有关非生物胁迫响应的线索。
由于全球气候变化的影响,各地区作物遭受环境胁迫的事件愈演愈烈,持续时间越来越长,作物产量和品质受到严重影响。不断增长的人口、有限的耕地面积和退化的土壤条件已经成为限制我国农业现代化和农业可持续发展的瓶颈。因此,当务之急是将非生物胁迫响应机制的研究引入作物体系。要实现增强作物对环境胁迫的抗性,目前还有诸多问题需要解决。其一,多数实验室使用水稻、番茄和拟南芥等作为研究材料,利用它们是模式植物的优势,能更加高效地开展试验。然而,温室条件和控制变量等研究环境或分析方法会忽略生物胁迫和非生物胁迫以及不同非生物胁迫之间的相互作用。例如,大田水稻长期处于高温、干旱和高盐逆境下,环境因子对作物生长和发育的限制具有多元性。作物所处的环境具有多变性,不同发育时期的逆境也并非一成不变。因此,探索植物对环境变化的精准识别和协同应答显得尤为重要。其二,作物在生长和发育的同时,还要适应极端环境。在异常气候下,作物常常受到非连续性逆境胁迫的影响,这些胁迫在不同生长时期都会出现,植物往往快速响应逆境并恢复生长。目前,还缺乏对植物平衡生长和抗逆的理论研究。最后,相比作物病虫害抗性种质资源,逆境适应性种质资源的获取显得尤为缓慢。主要作物响应逆境涉及多基因复杂性状位点的调控。同时,由于作物的育种年限较长,不确定性因素较多,它们的育种效率被严重限制。因此,还需综合运用基因组学、代谢组学和表观组学等组学研究手段,引入单倍体育种和基因编辑等新兴技术,以提高遗传位点选择的准确性,加快逆境适应性种质资源的培育效率。
猜你喜欢
汽车实用技术(2022年16期)2022-09-03
理财周刊(2022年4期)2022-04-30
昆明医科大学学报(2022年1期)2022-02-28
园艺与种苗(2020年12期)2021-01-08
学苑创造·A版(2020年12期)2020-01-07
汽车实用技术(2019年20期)2019-11-26
中国外汇(2019年15期)2019-10-14
中国石油大学胜利学院学报(2019年2期)2019-07-08
飞碟探索(2015年9期)2015-11-05
红领巾·探索(2015年9期)2015-09-10
