
35 日龄撒坝仔猪盲肠发育及微生物多样性研究
2021-12-17 03:37杨玉婷冉津铭沈丽艳蒋恒鑫李雪艳潘洪彬
中国畜牧杂志 2021年12期
杨玉婷,高 欢,冉津铭,沈丽艳,蒋恒鑫,李雪艳,杨 光,潘洪彬*
(1.云南农业大学动物科学技术学院,云南省动物营养重点实验室,云南昆明 650201;2.达州职业技术学院,四川达州 635000;3.云南文化艺术职业学院,云南昆明 650201)
我国是养猪大国,拥有丰富的猪品种遗传资源,地方品种猪达100 多种[1]。撒坝猪主要分布于滇中楚雄彝族自治州的禄劝、昆明、武定、楚雄等县、市,以禄劝县撒营盘(以前称撒坝)为中心而得名,是云南省“六大名猪”之一,具有耐粗饲、肉质优良、适应性强、抗逆性好等特性[2]。我国自1964 年开始饲养长白猪,其生长速度快,屠宰率和胴体瘦肉率高,是我国生猪饲养利用的主要猪种之一[3]。
猪肠道中栖息着大量的微生物,肠道微生物及其代谢产物与宿主营养物质的消化吸收、肠道健康状态和免疫反应等密切相关,因此,肠道微生物也被认为是动物的重要器官之一[4]。研究显示,单胃动物大肠中的微生物数量显著高于小肠,盲肠、结肠和直肠是微生物聚集的主要区段,并且盲肠核心菌群有助于维持肠道的正常功能和黏膜的完整性[5]。研究发现,国外商品猪种和中国地方猪种的肠道微生物区系差异较大[6]。然而,目前关于撒坝猪肠道形态学和微生物多样性的研究鲜见报道。本试验通过比较35 日龄撒坝和长白仔猪盲肠组织形态学和微生物多样性的差异,为合理开发利用撒坝猪及其肠道微生物资源提供基础数据。
1 材料与方法
1.1 试验动物与试验设计 选取均为第三胎的撒坝和长白母猪各3 头,同一环境下单栏饲养,饲喂NRC 无抗日粮,日粮原料组成及营养成分见表1。分娩后,母猪与其分娩的仔猪在同一栏中饲养,母乳饲养至35 日龄。每头母猪随机选取仔猪各3 头屠宰,每个猪种各9 头,共18 头仔猪;迅速取盲肠食糜于液氮保存;并取盲肠中段组织于4%甲醛固定液固定。

表1 日粮原料组成及营养成分
1.2 盲肠组织形态学测定 固定24 h 的盲肠组织,依次进行石蜡包埋、切片、苏木精-伊红(HE)染色,染色后用二甲苯透明5 min,用中性树胶加盖玻片封片,并利用Image-proplus 6.0(Media Cybernetics,Inc.,Rockville,MD,USA)软件观察盲肠形态,测量隐窝深度和肌层厚度。
1.3 16S rDNA 测序 18 个仔猪盲肠食糜样本,根据制造商说明,使用QIAamp®Fast DNA Stool Mini Kit(Qiagen,Cat No.:19593)试剂盒提取总DNA。扩增16S V3+V4 测序区域,引物序列Forard Primer(5'→3'):CCTACGGGRSGCAGCAG(341 F);Reverse Primer(5'→3'):GGACTACVVGGGTATCTAATC(806 R)。纯化后的PCR 产物文库用Qubit® dsDNA HS Assay Kit试剂盒进行定量检测,按照盲肠食糜样品的测序量要求,进行适当比例混合。利用Illumina Miseq(Miseq 2500 PE250)测序。撒坝和长白仔猪各有一个样品建库失败。
1.4 生物信息学分析 在Illumina Miseq 平台上,针对16S rDNA 的V3+V4 区进行扩增,利用双末端测序(Paired-End)方法,通过Pandaseq 软件利用重叠关系将双末端测序得到的成对Reads 拼接成一条序列,得到高变区的长Reads[7]。对拼接后的Reads 进行处理,获取Clean Reads。利用Usearch 序列分析软件在97% 相似度下进行聚类,对聚类后的序列进行嵌合体过滤后,得到用于物种分类的OTU。使用Venn 图确定每组中特定OTU 数量。使用QIIME 软件基于OTU结果计算每个样本的物种的Alpha 多样性[8]。通过样品之间的UniFrac 距离矩阵进行Beta 多样性分析。LEfSe(LDA Effect Size,Linear discriminant analysis Effect Size)分析用于确定组间差异最大的OTU[9]。利用KEGG 数据库(http://www.genome.jp/kegg),进行PICRUSt(phylogenetic investigation of communities by reconstruction of unobserved states)基因功能预测分析[10]。
1.5 统计分析 利用SPSS 22.0 软件进行独立样本T 检验,结果以平均数±标准误表示,P<0.05 为差异显著,P<0.01 为差异极显著。
2 结果
2.1 盲肠组织形态学分析 由图1 可知,撒坝仔猪盲肠肌层厚度高于长白仔猪(P<0.01),隐窝深度(P<0.05)和隐窝深度/肌层厚度比值(P<0.01)低于长白仔猪。

图1 撒坝和长白仔猪盲肠形态比较
2.2 OTU 聚类分析 由图2 可知,撒坝和长白仔猪盲肠食糜的OTU 分别为715 和722 个,其中共有OTU 为572 个,分别占到80.00%和79.22%,撒坝仔猪盲肠中特有OTU 为143 个,长白仔猪盲肠中特有OTU 为150 个。

图2 盲肠食糜OTU 组成
2.3 撒坝和长白仔猪盲肠微生物Alpha 多样性分析 由表2 可知,长白仔猪盲肠微生物丰富度指数observed species 和多样性指数PD whole tree 指数均高于撒坝仔猪(P<0.05)。
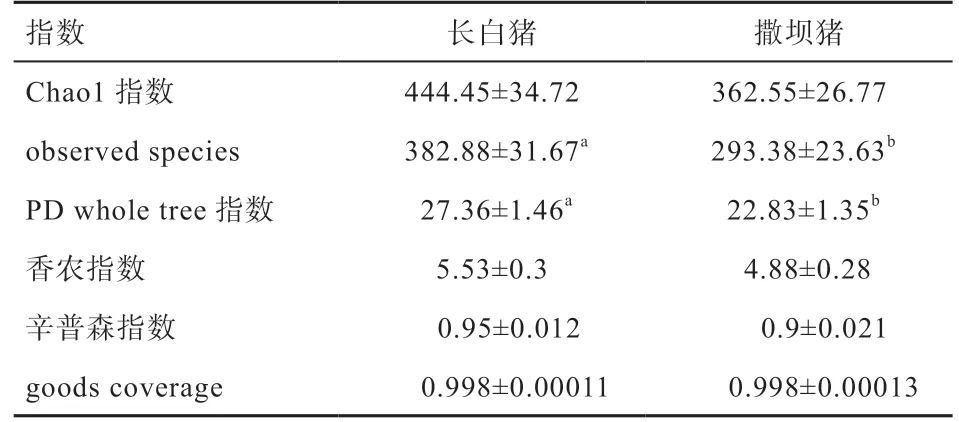
表2 长白和撒坝仔猪盲肠微生物Alpha 多样性
2.4 撒坝和长白仔猪盲肠微生物Beta 多样性分析 由图3 可知,基于样本间OTUs 的未加权unifrac 分析数据结果,绘制成PCoA 图形并在图上显示Adonis 分析结果。长白猪和撒坝猪明显分离,Adonis 分析结果为R2=0.166(P=0.001)。由图4 可知,Anosim 相似性分析发现R2=0.367(P=0.002)。由表3 可知,MRPP 分析显示未加权unifrac A=0.062(P=0.002)。以上结果表明长白和撒坝仔猪盲肠食糜微生物结构存在极显著差异(P<0.01)。

表3 MRPP 组间差异分析

图3 Adonis 分析PCoA 图

图4 Anosim 相似性指数分析图
2.5 撒坝和长白仔猪盲肠门水平微生物组成比较 由表4 可知,在门水平上,撒坝和长白仔猪盲肠食糜菌群相对丰度97% 以上主要由Bacteroidetes、Firmicutes、Fusobacteria 和Proteobacteria 组成;撒坝仔猪盲肠中Bacteroidetes 的相对丰度高于长白仔猪(P<0.05),长白仔猪盲肠中Proteobacteria 的相对丰度高于撒坝仔猪(P<0.05)。
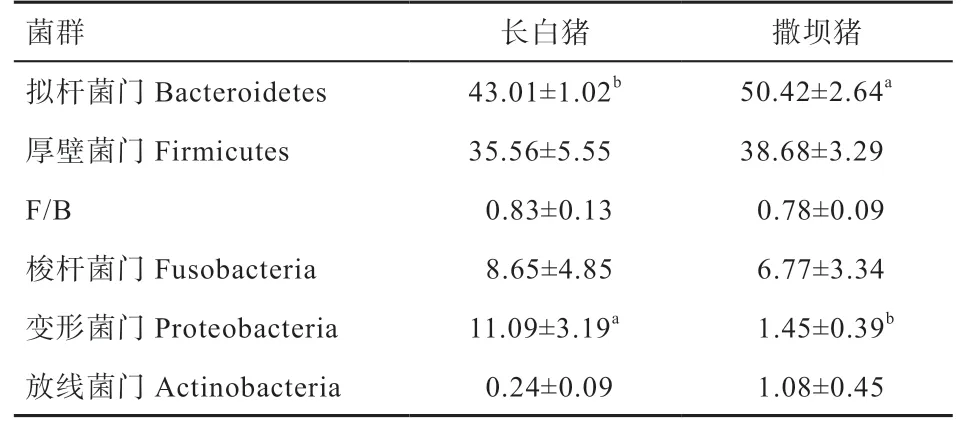
表4 盲肠微生物门水平物种丰度分布图
2.6 撒坝和长白仔猪盲肠属水平微生物组成比较 两猪种盲肠菌群在属水平上明显不同。由表5 可知,撒坝仔猪盲肠菌群相对丰度最高的4 个属依次为Bacteroides、Alloprevotella、Fusobacterium和Prevotella;长白仔猪盲肠中的优势菌群为Prevotella,其次为Bacteroides、Fusobacterium和Alloprevotella;撒坝仔猪盲肠中Bacteroides的相对丰度高于长白仔猪(P<0.05),而长白仔猪中Prevotella高于撒坝仔猪(P<0.01)。

表5 盲肠微生物属水平物种丰度分布图
2.7 撒坝和长白仔猪盲肠差异微生物分析 基于16S rDNA 基因测序所得的物种注释信息,我们进行了线性判别分析效应大小(LEfSe)的分析,以比较撒坝和长白仔猪的盲肠微生物差异,从而确定两猪种显著富集的微生物(图5)。撒坝仔猪盲肠中有1 个纲(class)包括Opitutae,1 个目(order)包括Bifidobacteriales,1个科(family)包括Bifidobacteriaceae,7 个属(genus)包括Megasphaera、ParaBacteroides、Mitsuokella、Acidam inococcus、Bifidobacterium、Gemmiger和Blautia显著富集。

图5 盲肠微生物LDA 值分布柱状图(P<0.05,LDA>2)
在长白仔猪盲肠中有3 个门(phylum)包括Fibro bacteres、Synergistetes 和Proteobacteria,5 个纲(class)包括Subdivision5、Fibrobacteria、Synergistia、Epsilon proteobacteria 和Gammaproteobacteria,4 个目(order)包括Fibrobacterales、Synergistales、Aeromonadales 和Campylobacterales,5 个科(family)包括Fibrobactera ceae、Enterococcaceae、Synergistaceae、Succinivibrion aceae 和Campylobacteraceae,14 个 属(genus)包 括Acetanaerobacterium、Anaerobiospirillum、Subdivision5 genera incertae sedis、Veillonella、Sutterella、Fibrobacter、Schwartzia、Enterococcus、Pyramidobacter、Cloacibacillus、Faecalicoccus、Roseburia、Succinivibrio和Campylobacter显著富集。
2.8 撒坝和长白仔猪盲肠微生物的KEGG 功能预测 由图6 可知,撒坝仔猪盲肠中显著富集了Galactose meta bolism、General function prediction only、Arginine and proline metabolism、Lysine biosynthesis、Glycine serine and threonine metabolism 和Other transporters 共6 个信号通路。
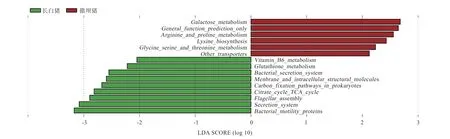
图6 盲肠微生物富集的信号通路LDA 值分布图(P<0.05,LDA>2)
长白仔猪盲肠中显著富集了Membrane and intracellular structural molecules、Secretion system、Carbon fixation pathways in prokaryotes、Citrate cycle TCA cycle、Bacterial secretion system、Glutathione metabolism、Vitamin B6 metabolism、Bacterial motility proteins 和Flagellar assembly 共9 个信号通路。
3 讨 论
仔猪肠道健康是影响养猪业发展的重要因素之一,仔猪消化系统发育的成熟程度直接影响仔猪的消化吸收[11]。盲肠是猪进行纤维素发酵和分解的主要场所,盲肠没有绒毛和环形皱襞,但其肠腺发达,肠腺分泌的碱性黏液与食糜发酵产生的酸性产物不含消化酶,但有溶菌酶[12]。肠腺又称肠隐窝,隐窝深度影响肠上皮细胞的生成率,补充肠道上皮中脱落的细胞,深度越浅说明成熟细胞越多,肠道吸收能力越强,另外,肠道的肌层厚度影响肠道蠕动能力,肠道蠕动能力强则对食糜消化能力强[13]。本研究中撒坝仔猪盲肠肌层厚度高于长白仔猪,而隐窝深度和隐窝深度/肌层厚度比值低于长白仔猪,表明撒坝仔猪盲肠发育状况优于长白仔猪。
遗传(品种)是影响猪肠道微生物的重要因素之一[14-15]。研究发现240 日龄金华猪盲肠微生物多样性高于长白猪[6];然而,本研究发现35 日龄长白仔猪盲肠微生物丰富度指数observed species 指数和多样性指数PD whole tree 指数高于撒坝仔猪,即长白仔猪盲肠微生物Alpha 多样性高于撒坝仔猪,这与先前的研究结果不一致,这可能是年龄和品种的差异导致。仔猪的交叉培养模型表明,哺乳母猪会影响仔猪的生长性能[16]和盲肠微生物的组成[17]。本课题组前期研究发现撒坝仔猪的初生重、35 日龄体重、平均日增重和体尺性状均显著低于长白仔猪,35 日龄撒坝仔猪空肠微生物多样性及组成区别于长白仔猪[18];本研究结果显示撒坝仔猪的盲肠微生物组成和肠道形态学存在较大差异。因此,哺乳期撒坝和长白仔猪生长性能、肠道发育和肠道微生物组成的差异与宿主遗传和哺乳母猪相关。
肠道内核心菌群直接影响肠道功能的正常发挥[19]。研究发现,金华猪和长白猪盲肠门水平上的优势菌群为Bacteroidetes 和Firmicutes[6],这与本研究中Firmicutes和Bacteroidetes 是撒坝和长白仔猪盲肠中的优势菌群的结果一致。Bacteroidetes 菌群具有发酵碳水化合物、参与糖类代谢、胆汁酸和类固醇代谢等诸多功能[20]。本研究结果显示,撒坝仔猪的Bacteroidetes 显著高于长白仔猪,且KEGG 功能预测结果显示撒坝仔猪盲肠微生物与半乳糖代谢(Galactose metabolism)相关。提示撒坝仔猪通过盲肠微生物调控糖类代谢的能力可能优于长白仔猪。
肠道微生物存在3 种肠型,分别为以Bacteroides为指示类群的肠型1(ETB);以Prevotella为驱动的肠型2(ETP),其丰度通常与拟杆菌的丰度成反比;Firmicutes(Ruminococcus为主)占比高的肠型3(ETF)[21]。ETB 菌群结构与富含动物蛋白和饱和脂肪酸饮食相关,并且具有Bacteroides特异的碳水化合物酶(CAZymes),与此相反ETP 肠型分解脂肪和蛋白的能力较低[22]。本研究中,撒坝仔猪盲肠的Bacteroides丰度高于长白仔猪,而长白仔猪盲肠的Prevotella丰度高于撒坝仔猪,表明撒坝仔猪为ETB 肠型,而长白仔猪为ETP 肠型,提示撒坝仔猪盲肠微生物对脂肪和蛋白的利用能力可能强于长白仔猪。
猪的盲肠是微生物发酵碳水化合物的主要场所。微生物通过发酵碳水化合物产生短链脂肪酸(SCFAs),主要为乙酸、丙酸和丁酸。Firmicutes 菌群中的大部分细菌属于发酵型细菌,具有更高的从饮食中获取能量的能力,发酵出更多的SCFAs,从而促进脂肪沉积[23]。本研究结果显示富集在撒坝仔猪盲肠中的Megasphaera、Mitsuokella、Acidaminococcus、Gemmiger和Blautia均属于Firmicutes 菌群。其中,Megasphaera和Mitsuokella也能够利用乳酸和琥珀酸酯产生短链脂肪酸(SCFAs)[24-25]。乙酸是胆固醇和脂肪酸的潜在前体物质,丁酸常常作为结肠上皮细胞的能量底物,丙酸是肝脏糖异生的底物[26]。此外,SCFAs 能降低pH 进而维持肠道微生态系统稳定[27]。因此,作为肠道微生物菌群产生的非直接营养物质,SCFAs 在调节能量代谢、免疫、肠道发育方面有着重要的作用。本研究结果表明撒坝仔猪可能通过盲肠的Megasphaera、Mitsuokella等Firmicutes 菌群产生SCFAs 调节机体的能量代谢、免疫和肠道发育。
人们普遍认为地方猪的抗病能力强于外来品种,推测可能与猪肠道中的微生物有关[28]。Proteobacteria 是细菌中最大、最多样化的一门,在动物胃肠道疾病的临床诊断中具有重要的意义,研究发现肠道炎性疾病发生会导致肠道中Proteobacteria 数目增加[29]。而Bifidobacterium是动物肠道内的优势菌群之一,具有免疫调节、抗肿瘤、抗致病、抗炎症等多种生物学功能,是重要的益生菌,Bifidobacterium可通过特殊的双歧支路代谢途径分解糖类碳水化合物最终生成乙酸和乳酸[30-31]。并且Bifidobacterium还可分泌a、β-半乳糖苷酶、α、β-葡萄糖苷酶及人体不能消化的寡糖的肽酶、磷蛋白磷酸酶、胆酸代谢酶、结合胆酸水解酶、降解N-亚硝酸的酶等许多种酶类参与代谢[32]。本研究中长白仔猪盲肠Proteobacteria 丰度高于撒坝仔猪以及撒坝仔猪盲肠Bifidobacterium丰度高于长白仔猪,提示撒坝仔猪的强抗病能力可能与盲肠中低丰度的Proteobacteria 和高丰度Bifidobacterium有关。
Mitsuokellasp.和Megasphaeraelsdenii-like是继代培养中利用赖氨酸、苏氨酸、精氨酸和谷氨酸的优势菌[33]。本研究中,Megasphaera、Mitsuokella显著富集在撒坝仔猪盲肠中,且KEGG 功能预测结果显示撒坝仔猪盲肠微生物主要与氨基酸代谢相关(精氨酸和脯氨酸代谢、赖氨酸的生物合成、甘氨酸、丝氨酸和苏氨酸代谢),因此,撒坝仔猪可能通过Mitsuokella.和Megasphaera促进精氨酸、脯氨酸代谢和赖氨酸等多种氨基酸的合成与代谢,这为进一步解释不同品种猪在氨基酸利用上的差异研究提供了思路。
4 结 论
本试验结果显示,撒坝仔猪盲肠发育较好,食糜消化能力更强;撒坝仔猪盲肠微生物多样性低于长白仔猪,撒坝仔猪倾向于ETB 肠型,撒坝仔猪通过盲肠微生物对能量的吸收和储存能力可能优于长白仔猪并且撒坝仔猪可能通过Mitsuokella和Megasphaera促进精氨酸、脯氨酸代谢和赖氨酸等多种氨基酸的合成与代谢。
猜你喜欢
现代交际(2022年10期)2022-11-01
今日农业(2022年14期)2022-09-15
中老年保健(2022年2期)2022-08-24
中国饲料(2022年5期)2022-04-26
昆明医科大学学报(2022年3期)2022-04-19
江苏农业科学(2019年8期)2019-08-20
意林·少年版(2018年11期)2018-07-04
中外医学研究(2017年15期)2017-06-29
扬子江(2016年1期)2016-05-19
环球慈善(2009年11期)2009-12-27
