
低温胁迫对拔节期小麦光合色素、光合参数及叶绿素荧光特性的影响
2021-12-17 11:04:18葛君,刘震
山西农业科学 2021年11期
葛 君,刘 震
(商丘市农林科学院,河南商丘476000)
光合作用是植物进行生长发育的物质基础,但植物的光合作用对低温、高温、干旱等在内的非生物逆境胁迫较为敏感[1-2],因此,光合特性常被用作反映植物对逆境胁迫响应程度,判断植株生长状况及抗逆性强弱的重要指标[3]。不适宜的温度往往会通过影响植物的光合能力进而对植物各个生育阶段产生不利影响,最终导致作物产量减少和品质降低。受厄尔尼诺等现象的影响,低温等极端天气发生的频率、程度不断增加[4],小麦越冬期遭遇的冻害、返青期和拔节期受到的冷害等低温灾害,会严重制约小麦品质和产量的提高[5]。因此,加快研究低温胁迫对小麦形态特征和生理特性方面的影响,对灾害天气小麦防灾减灾具有重要意义。
有研究表明,叶绿素荧光动力学技术在测定植物对光能的吸收、传递、耗散、分配等方面具有独特的作用[6],它可以在不对植物叶片细胞造成破坏的前提下便捷、可靠、快速地反映植物光合系统的内在特性与光合作用的变化过程,因此,在植物胁迫生理学方面得到广泛应用。其中,Fo为固定荧光,能够反映光系统Ⅱ反应中心处于完全开放状态下的荧光产量;Fv/Fm为PSⅡ最大光化学量子产量,能够反映光系统Ⅱ反应中心的光能转换效率;qP为光化学淬灭,表示PSⅡ天线色素吸收的光能占光化学电子传递的份额[6]。因此,采用叶绿素荧光研究植物逆境下的生理变化是当前生理研究的重要内容之一,对研究逆境条件下小麦生理变化机制具有重要意义。
研究表明,植物叶片叶绿素荧光参数的变化受温度的影响[7],目前针对低温胁迫对不同小麦品种光合色素含量方面的研究报道较多[8],但这些试验大都是在恒定低温胁迫下对小麦叶片光合速率变化情况及叶片受害症状进行研究[9-10],而有关采取渐进式降温—恒温—升温动态变温模式研究低温胁迫小麦光合特性等生理指标的研究尚未见报道。
本试验采取渐进式降温—恒温—升温动态变温模式研究低温胁迫对拔节期小麦光合色素、光合参数及叶绿素荧光特性的影响,以期为小麦的防灾减灾、安全生产、优质高产提供理论指导,为小麦抗逆丰产育种提供数据参考。
1 材料和方法
1.1 试验材料
供试小麦品种为济麦22,由山东省农业科学院作物研究所提供。
1.2 试验方法
将小麦种子播种于直径30 cm、高35 cm花盆中,底部有排水孔,播种后将花盆埋于田间,高度与大田土壤平齐,按照大田管理方法进行管理。花盆内装入20 cm耕层大田土(土质为沙壤土,碱解氮93.00 mg/kg、速效磷30.07 mg/kg、速效钾87.36 mg/kg、有机质15.49 g/kg),共播种45盆,待麦苗长至3片叶时进行定苗,每盆留苗20株。
试验设置5个温度水平,包括10℃对照(CK)以及0、-2、-4、-6℃等4个低温胁迫处理(分别记作T1、T2、T3、T4)。每3盆作1个重复,每处理设3次重复,即每处理9盆。在小麦拔节期第7天,将各处理分别置于相应温度的人工气候室内进行12 h低温处理,其处理方法为:采用渐进式降温和升温方式进行低温处理,以接近外界环境的降温—升温规律,即用3 h的时间将温度从对照温度10℃分别降至每一个处理温度,并在该处理温度下胁迫6 h,然后再用3 h升温至10℃,总计12 h。空气相对湿度为60%~65%。胁迫结束后将花盆置于大田,并于第2天9:00—10:00进行叶绿素含量、叶绿素荧光参数、光合参数的测定。
1.3 测定项目及方法
利用FMS-2便携式荧光仪(Hansatech,UK),参照GENTYB等[11]的方法测定小麦植株倒3叶的初始荧光(Fo)、最大光化学效率(Fv/Fm)、PSⅡ实际光化学效率(ФPSⅡ)、光化学淬灭(qP);采用美国Li-COR公司生产的Li-6400XT型便携式光合仪测定小麦植株倒3叶的净光合速率、蒸腾速率、胞间CO2浓度、气孔导度;采用丙酮浸提法[12]测定小麦植株倒3叶的叶绿素含量。
1.4 数据分析
利用Excel 2007进行数据处理和作图;采用SPSS 25.0软件进行数据统计分析。
2 结果与分析
2.1 低温胁迫对拔节期小麦叶片叶绿素含量的影响
由表1可知,不同低温处理对拔节期小麦叶绿素含量的影响存在差异,4个低温处理下的小麦叶片叶绿素a含量均显著低于对照(P<0.05),较对照分别降低12.80%、22.27%、42.18%、50.32%;随着胁迫温度的降低,叶绿素b、叶绿素(a+b)含量均呈逐渐下降的变化趋势,至T4处理达最小值,较对照分别降低40.14%、47.73%,且与对照差异均达显著水平(P<0.05)。在T1处理下,小麦叶片类胡萝卜素含量与对照差异不显著(P>0.05),说明0℃低温对拔节期小麦叶片类胡萝卜素含量的影响较小;在T2、T3、T4处理下,类胡萝卜素含量均显著低于对照(P<0.05),较对照分别降低27.87%、40.98%、49.18%。

表1 低温胁迫对拔节期小麦叶片叶绿素含量的影响 mg/g
2.2 低温迫胁对拔节期小麦叶片光合特性的影响
由表2可知,不同低温处理对拔节期小麦叶片光合参数的影响不同。4个低温处理下的小麦叶片净光合速率均低于对照,且随温度的降低净光合速率逐渐下降,与对照相比分别显著降低10.10%、19.59%、47.69%、59.33%(P<0.05);4个低温处理下的小麦叶片蒸腾速率和气孔导度的变化规律与净光合速率一致,且在T4处理下达最小值,较对照分别降低55.43%和63.52%,且均与对照间差异达显著水平(P<0.05)。
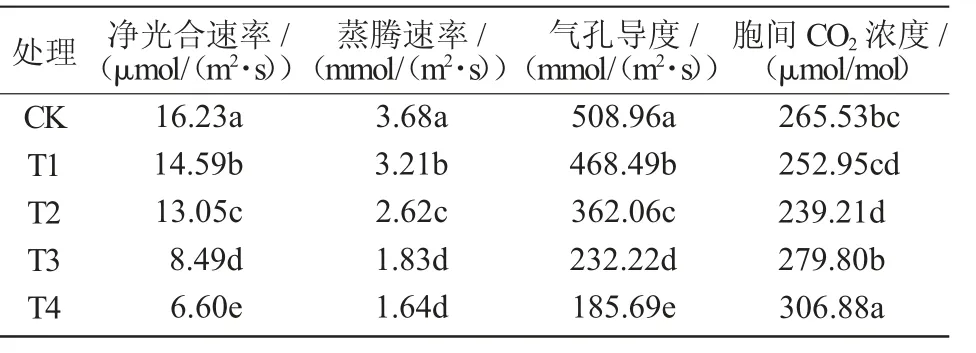
表2 低温胁迫对拔节期小麦叶片光合特性的影响
随着胁迫温度的下降,拔节期小麦叶片胞间CO2浓度表现出先降低后升高的变化趋势,且在T3、T4处理时高于对照,较对照分别增加5.10%和15.57%,其中,T3处理与对照间差异不显著(P>0.05),而T4处理与对照间差异达显著水平(P<0.05)。
2.3 低温胁迫对拔节期小麦叶片叶绿素荧光特性的影响
由图1可知,随着胁迫温度的降低,拔节期小麦叶片Fo逐渐升高,T1处理下与对照差异不显著(P>0.05),T2、T3、T4处理较对照分别增加18.52%、45.45%、56.52%,且与对照差异显著(P<0.05);随着胁迫温度的降低,叶片Fv/Fm表现出逐渐下降的趋势,T1处理下与对照差异不显著(P>0.05),T2、T3、T4处理与对照差异显著(P<0.05),表明随着胁迫温度的降低拔节期小麦叶片PSⅡ反应中心的光能转化效率下降;ФPSⅡ和qP的变化趋势相似,均随胁迫温度的降低而降低,但在T1处理下与对照差异不显著(P>0.05),其余3个处理与对照差异均达显著水平(P>0.05);且在T4处理下达最低值,较对照分别降低38.60%、25.00%。
综上可见,T1处理下各指标与CK差异不显著,T2、T3、T4处理下与对照间差异均达显著水平,说明低温胁迫达到-2℃及以下时会对拔节期小麦叶片光合器官造成一定程度的损伤。

3 结论与讨论
叶绿素是绿色植物进行光合作用的重要场所,在植物光合过程中起着捕获光能、吸收光能的重要作用,其含量通常会对植物的光合能力产生影响[13]。和红云等[14]研究得出,低温胁迫能够降低甜瓜幼苗叶片叶绿素含量,且随温度的降低胁迫加剧。陆新华等[15]研究认为,低温胁迫能够降低菠萝幼苗叶片叶绿素含量,且随着胁迫时间的延长,叶绿素含量呈逐渐降低趋势。本试验结果表明,低温胁迫能够降低拔节期小麦叶片光合色素含量,且叶绿素a、叶绿素b、类胡萝卜素含量随胁迫温度的降低而降低,试验结果与前人研究结论[13-15]一致,这可能是因为低温降低了酶的活性,从而减少叶绿素生物合成酶的合成[16];另外,低温通过影响δ-氨基乙酸丙酸(ALA)向单卟啉胆色素原(PBG)的转化和Mg-卟啉IX(Mg-Proto IX)向Mg-原卟啉甲酯(Mpe)的转化,降低了PBG、尿卟啉原Ⅲ(UrogenⅢ)、粪卟啉原Ⅲ(CoprogenⅢ)、原卟啉Ⅸ(ProtoⅨ)、原叶绿素酸酯(Pchlide)、叶绿素酸酯(Childe)的积累量,降低叶绿素a和叶绿素b的产量,并最终使叶绿素(a+b)含量减小[17]。
本研究结果表明,在T1、T2处理下拔节期小麦叶片净光合速率与气孔导度、胞间CO2浓度变化趋势一致,随胁迫温度的降低而降低,可能是由于低温胁迫使小麦叶片气孔收缩,导致气孔导度下降,胞间CO2浓度降低,进而使叶片的光合作用受到抑制[18]。在T3、T4处理下,拔节期小麦叶片净光合速率、气孔导度降低,而胞间CO2浓度却升高,表明此时净光合速率的降低为非气孔限制因素所致,可能是由于低温胁迫下酶活性降低,使光合碳同化过程对CO2浓度变化不敏感,从而使植物的光合能力降低[19];也可能是由于过低的温度胁迫使小麦叶片组织受到破坏,进而降低了小麦的光合速率[20]。
Fo随低温胁迫程度的加剧而升高,在温度达到-2℃及以下时与对照差异显著,而在0℃时Fo与对照差异不显著,说明-2℃及以下的低温已对拔节期小麦叶片光合器官造成了损伤,且这种损伤随Fo的增加而增加[21]。Fv/Fm随温度的降低而降低,试验结果与胡春梅等[22]、王兆等[23]的研究结论一致,这可能是因为低温胁迫能够降低Rubisco酶活性,从而使植株叶片对CO2的同化力降低,进一步导致卡尔文循环过程中对ATP和NADPH需求量减少,进而引起反应中心光能转换效率降低[24]。
本研究采用盆栽法对济麦22拔节期进行0、-2、-4、-6℃不同低温处理,结果发现,光合色素含量随温度降低而减少,光合速率随温度降低而减慢;叶绿素荧光特性结果显示,0℃处理下Fo、Fv/Fm、ФPSⅡ、qP值与对照差异不显著,而更低温度下均与对照差异显著,可能是由于0℃时光合器官未造成损伤,而低于-2℃时会对光合器官造成损伤。本试验虽然研究得出了拔节期小麦不同生理指标对低温胁迫的响应状况,但还需要在后期进一步研究低温胁迫对拔节期小麦细胞生物学及分子生物学水平上的变化机制,并找出抵御逆境低温胁迫伤害的防御措施,为应变小麦防灾减灾提供技术支持,为小麦高产栽培技术体系的进一步完善提供资料参考,为小麦的抗逆育种提供理论依据。
猜你喜欢
真空与低温(2022年6期)2023-01-06 07:33:20
今日农业(2021年6期)2021-11-27 08:05:59
现代塑料加工应用(2021年5期)2021-02-28 08:18:04
阅读(科学探秘)(2020年8期)2020-11-06 06:22:48
中国果业信息(2019年1期)2019-01-05 17:41:42
劳动保护(2018年8期)2018-09-12 01:16:18
生物学教学(2017年9期)2017-08-20 13:22:32
动物营养学报(2015年10期)2015-12-01 06:45:19
发明与创新(2015年30期)2015-02-27 10:39:49
食品工业科技(2014年6期)2014-05-10 06:04:50
