
草海国家自然保护区4种森林群落稳定性评价
2021-12-17 02:40:46郭其强罗丝琼周浩然
中南林业科技大学学报 2021年11期
郭其强,罗丝琼,刘 志,周浩然,高 超,杨 瑞
(贵州大学 a.贵州省森林资源与环境研究中心,b.林学院,c.贵州省高原山地林木培育重点试验室,贵州 贵阳 550025)
森林群落稳定性是林分结构与功能的综合体现[1-2],也是森林生态系统维持稳定和发挥良好综合效益的基础[3]。研究如何构建稳定的森林群落对指导植被恢复和生态建设都极具理论和实践价值。以往国内外大多数学者关注物种多样性与群落稳定性的关系较多[4-6],而对群落生态功能与稳定性的关系考虑较少。有研究认为稳定的群落结构应该有相对稳定的年龄结构[7];也有研究认为较高的物种多样性和复杂的结构对群落稳定性有利[8]。近年来,有研究表明土壤理化性质、群落生产力及生境条件等都对森林群落稳定性具有重要影响[9-10]。可见,森林群落的稳定性与其组成特性、维持潜力、生态功能及外界干扰等都存在密切关系。
长期以来,学者们尝试用多种方法评价森林群落稳定性。如,M.Godron 稳定性测度法、数学模型法、转移概率法、主成分分析法及空间结构指数法等[7,11-14],因存在评价指标不够全面、计算过程繁杂及理论晦涩难懂等弊端[15],基层林业工作者理解和运用较为困难。后来,有人采用计算过程相对简便的隶属函数值法,综合运用森林群落物种特性、土壤肥力、林地承载力及外界干扰强度等指标,突出了对群落调查中获取林分数据的应用,评价过程不存在繁琐的数据处理,应用性与操作性强,在我国陕北黄龙山、黑龙江大兴安岭及吉林长白山及等地均有采用且效果较好[9-10,16-17]。
草海国家级自然保护区(以下简称保护区)地处贵州省威宁县城郊,属岩溶湿地类型,在调控局部水量平衡和维持区域小气候作用显著[18]。在保护区120 km2总面积中,森林面积占比约30%,广阔的水域面积使其成为黑颈鹤等多种鸟类的重要越冬栖息地,极具生态和科研价值。然而,在1999—2015年期间林地和灌草地面积占保护区总面积的比例分别下降了4.12%和3.21%[19],原因是频繁的人类活动导致原生植被破坏严重,森林面积锐减,以至于生物多样性降低、水土流失严重及水污染加剧等生态问题日渐突出[20]。
近年来,当地政府和相关部门针对保护区开展了一系列保护性工作。如,划分生态功能区、调整土地利用比例及加强保护区生态监测等[18,21-22]。这些措施虽能暂时缓解区域生态恶化进程,但因缺乏一定规模和质量的植被支撑,并未能有效的提升保护区的整体生态功能与效益[23]。如何营造高效稳定的森林生态系统,遏制环境恶化和维护生态安全是保护区面临的重要问题之一。基于此,本研究选择草海自然保护区现存发育相对较好的森林资源,以开展群落调查与取样分析为基础,通过分析不同群落类型的更新潜力、土壤肥力状况、林地承载力及外界干扰强度等特征,综合评价不同类型森林群落的稳定性,以期为保护区人工林建设与植被恢复提供科学的理论依据。
1 研究区概况
研究区位于贵州省威宁县草海自然保护区周边的人工林区(26°47′~26°57′N,104°9′~104°20′E),海拔2 150~2 300 m,坡度<10°。年平均气温10.6℃,最冷(1月)和最热(7月)平均气温分别为1.6℃和17.8℃;年均降水量约为950 mm,5—10月降水量约占全年总量的85%,年均蒸发量约为550 mm;属亚热带半湿润气候,具有光照充足、长冬无夏和干湿季分明的特点。林区土壤主要为黄棕壤,土层较厚,pH值4.5~6.5。植被类型主要为温带针叶林、针阔混交林、阔叶混交林及稀疏灌丛等,乔木树种以云南松Pinus yunnanensis、华山松P.armandii、旱冬瓜Alnus nepalensis和滇杨Populus yunnanensis等为主;灌木树种主要有平枝栒子Cotoneaster horizontalis、大白杜鹃Rhododendron decorum、滇榛Corylus yunnanensis和威宁小蘗Berberis weiningensis等;草本层以羊茅Festuca ovina、荩草Arthraxon hispidus、车前Plantago asiatica和云南兔儿风Ainsliaea yunnanensis等为主。
2 研究方法
2.1 样地设置与调查
经充分踏查,在研究区选择林分结构完整、林相整齐且外界干扰少的4 种森林群落类型,设置20 m×20 m 典型样地共23 块,每个群落类型的样地概况及编号见表1。每块样地内沿任一对角线设5 m×5 m 的灌木样方3 个,1 m×1 m 的草本及更新幼苗样方各3 个,于2018年7月中旬进行调查和取样。
调查内容:1)生境概况 地形地貌、土壤类型、坡度、坡向及人为干扰强度等;2)乔木调查 以样地一边为X 轴,另一垂直边为Y 轴建立直角坐标系,记录样地内所用乔木个体的种名、坐标值,并测定胸径(DBH)、树高、冠幅和枝下高等;3)灌木和草本调查 分物种测定其多度(簇数)、盖度、高度及冠幅等;4)土壤样品采集 在各草本样方内挖土壤剖面,有地面向下分3 个土层深度(0~20、20~40、40~60 cm)用环刀取样,带回实验室备用;5)乔木及灌草生物量 乔木采用植株地上生物量代替,计算方法依据曾伟生等[24]构建的通用生物量模型(M=a×DBHb,其中M为生物量;a 按针叶和阔叶树种不同分别取0.127 和0.130,b 值为2.33)计算确定;灌草生物量在样方中盖度大于10%的灌草物种,在每个样方内选择3 株标准个体,全株挖出后分根、径、叶取样,分别称鲜质量200 g,室内85℃下烘干至恒质量,求出含水率,进而得出每个物种标准株个体生物量和样方单位面积生物量,据此计算样地灌草生物量。6)林分年龄判定:在纯林中选择3 株径级最大的乔木,混交林中则在每个乔木种类中各选择3 株径级最大的单株,分别用生长锥钻取树芯,最后根据该样地中取样树芯年轮数的最大值来判定林分年龄,以此作为计算林地灌草生产力的依据。
2.2 评价指标及因子
1)种群更新潜力 统计群落内乔木树种的幼苗、幼树比例,作为评价种群更新潜力的依据。按DBH 把乔木个体划分为幼苗(DBH ≤1 cm)、幼树(1 cm<DBH<10.0 cm)和大树(DBH ≥10.0 cm),据此统计其比例。
2)物种多样性 参与评价因子为物种丰富度指数(S)、Simpson 指数、Shannon-Wiener 指数和Pielou 指数[20],公式如下:
物种重要值:(相对高度+相对盖度+相对多度)/3;
丰富度指数(S),等于样地内总物种数;

式中:S为群落总物种数;Pi=Ni/N为第i 个物种的相对重要值,Ni为种i 的重要值,N为群落内种i所在林层所有物种重要值之和。
3)土壤肥力 参与评价因子为土壤有机质、全氮、全磷和全钾含量,其值为3 个土层深度下土壤养分含量的平均值,测定方法依次为重铬酸钾外加热氧化法、半微量凯氏法、酸融钼锑抗比色法和醋酸铵法[25]。
4)林地承载力 参与评价因子为乔木地上生物量、灌木生和草本的年均生物量。
5)外界干扰强度 参与评价因子为外界干扰强度。根据样地调查时的人为赋值确定,范围为-1~1[26]。
2.3 评价方法
为消除不同评价因子间数量级与量纲对评价结果的影响,先对各因子原始数据进行标准化转换,采用公式:
Xik=Xjk/Xkmax×1 000。
式中:为Xjk的标准化数据,Xjk为第j项指标第k个评价因子的实测值,Xkmax为第k个评价因子的最大值。
计算各群落指标的隶属函数值,采用公式:
U(Xijk)=Aj(Xijk-Xkmin)/(Xkmax-Xkmin)。
式中:U(Xijk) 为第i群落类型第j个组织层次(种群更新潜力、物种多样性、土壤肥力、林地承载力、外界干扰)第k项指标隶属度,且U(Xijk)∈[0,1];Aj表示5 个组织层次指标的权重值,在参考前人研究成果[27]及本研究区概况的基础上,对5 个评价层次指标权重分别赋值:种群更新潜力0.30、物种多样性0.20、土壤肥力0.30、林地承载力0.10、外界干扰0.10;Xijk表示第i 群落类型第j个组织层次第k个指标值;Xkmax、Xkmin为所有参试群落类型中第k项指标的极大值和极小值。用每个群落类型各项指标隶属度的平均值作为综合评判的标准,比较不同群落类型的稳定性。
3 结果与分析
3.1 森林群落类型特征
通过对研究区4 种森林类型设置典型样地及调查数据,统计其群落基本特征见表1。
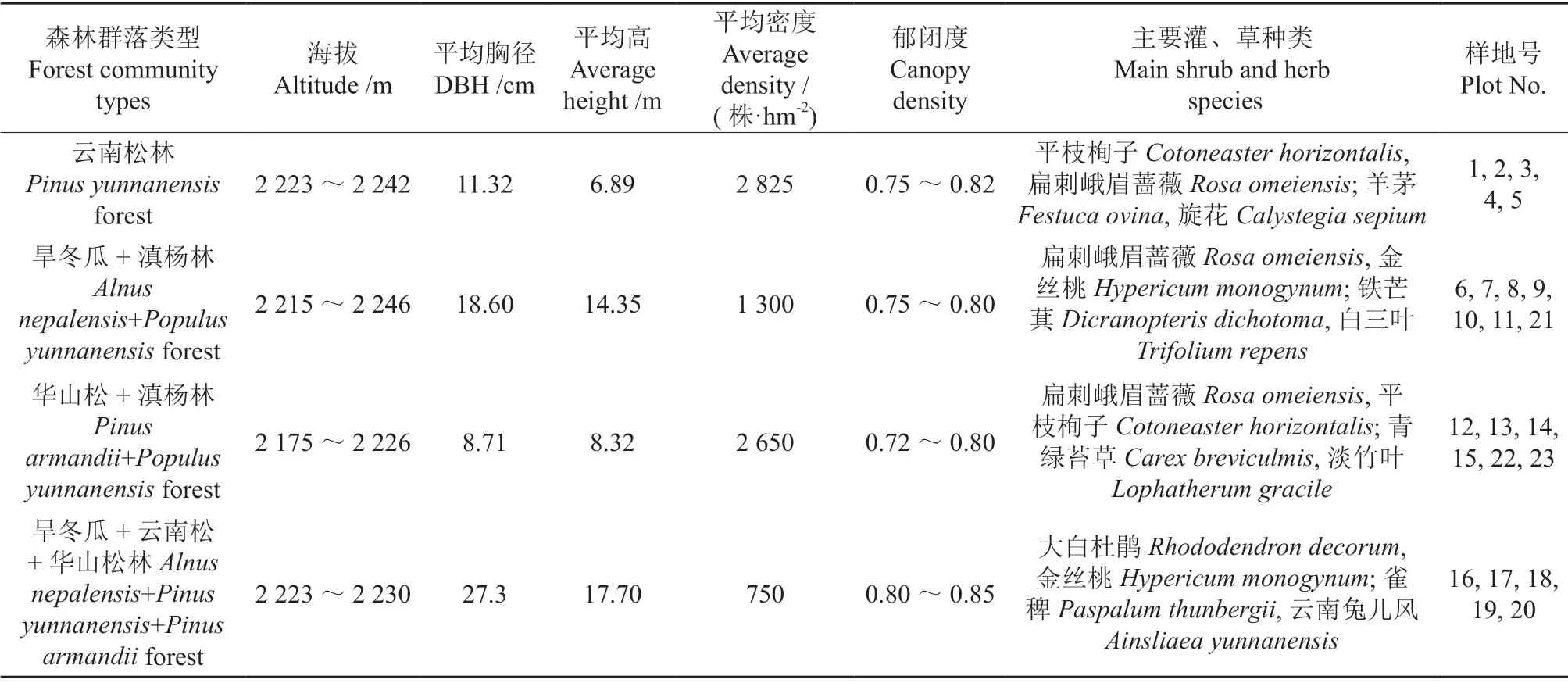
表1 草海国家级自然保护区4 种森林群落基本特征Table 1 Characteristics of four forest community types in Caohai national natural reserve
3.2 种群更新潜力
森林中幼苗、幼树数量及所占乔木树种的数量比是群落稳定性得以维持的重要基础[28]。本研究4 种森林群落中,幼苗、幼树个体数分别占乔木树种个体总数的53.54%、80.01%、50.54%和85.92%,而群落Ⅴ中无幼苗幼树存在。由此可见,由旱冬瓜、云南松和华山松组成的针阔混交林更新潜力最好,其次分别为旱冬瓜+滇杨林和云南松纯林及华山松+滇杨林。这说明不同森林类型在更新能力上多树种组成的针阔混交林具有明显优势,而乔木树种较少或纯林则呈劣势。
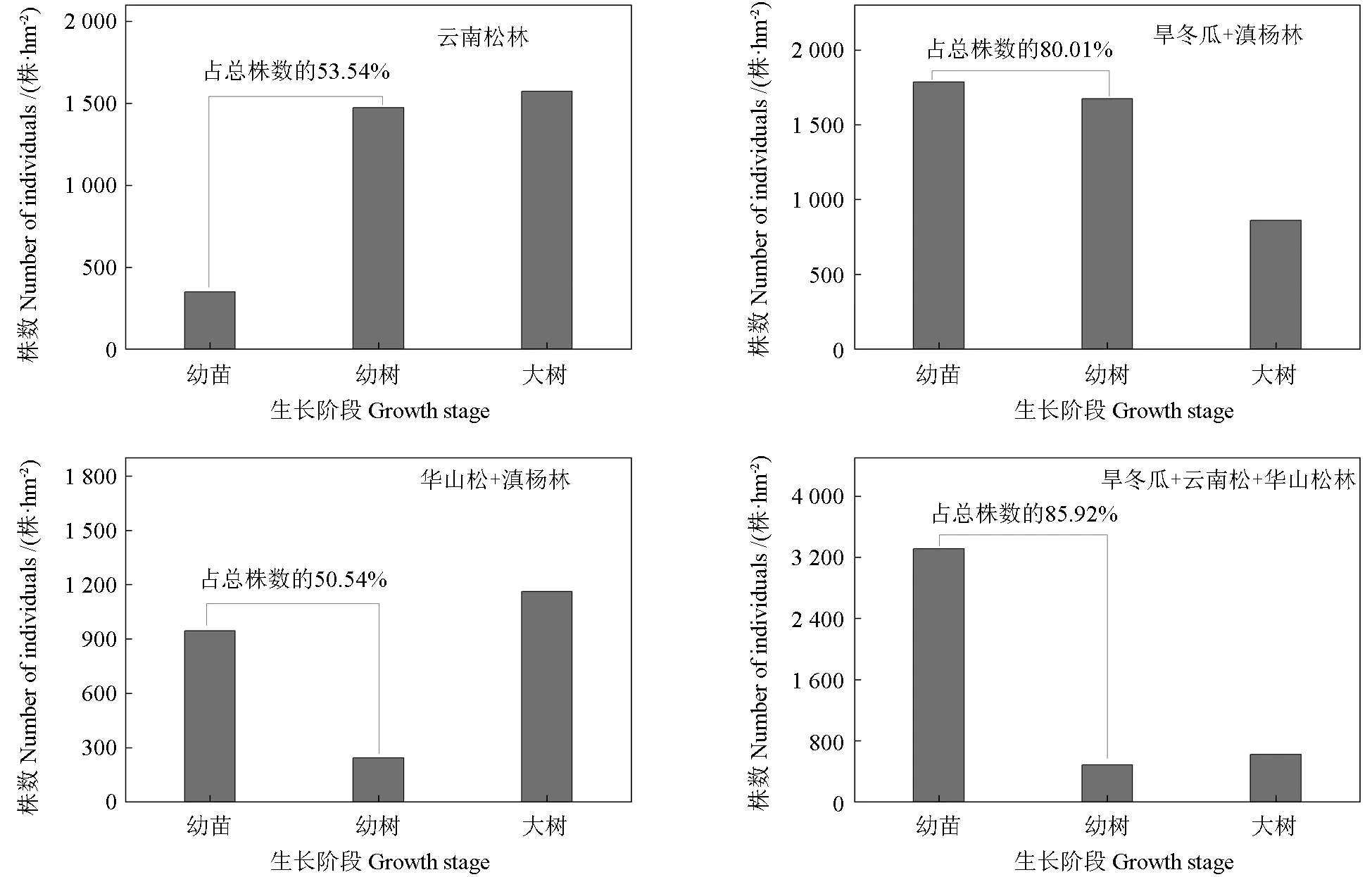
图1 4 种群落类型幼苗、幼苗占群落乔木总株树的比例Fig.1 Proportions of seedlings and seedlings in the total tree of four forest community types
3.3 物种多样性
4 种群落类型中不同物种多样性指数存在显著差异。由表2可知,乔木层物种多样性指数S、D和H′以旱冬瓜+云南松+华山松林和华山松+滇杨林较高,而云南松林中最低;J指数以旱冬瓜+滇杨林中最高而旱冬瓜+云南松+华山松林最低,且与其他各群落均存在显著差异(P<0.05)。在灌木层中,云南松林和旱冬瓜+云南松+华山松林的物种多样性指数均较高,其次为旱冬瓜+滇杨林,而华山松+滇杨林和旱冬瓜+云南松+华山松林中较低;多重比较结果表明云南松林与旱冬瓜+云南松+华山松林、华山松+滇杨林和旱冬瓜+云南松+华山松林两两间均无显著差异(P>0.05)。在草本层中,群落Ⅳ的各物种多样性指数(除J指数外)显著高于其余群落(P<0.05);且华山松+滇杨林和旱冬瓜+云南松+华山松林中J指数均无显著差异(P>0.05)。总体看,由旱冬瓜、云南松和华山松3 种乔木组成的针阔混交林物种多样性指数相对较高,而纯林则相对较低。

表2 不同群落类型物种多样性指数†Table 2 Species diversity indexes of different community types
3.4 土壤肥力及林地生产力
土壤肥力与林地生产力共同反映着林地的生态承载能力[29],其结果可以乔木和灌草年均生物量值为代表[30]。由表3可知,全氮含量以云南松林和旱冬瓜+云南松+华山松林为最高,而全磷和全钾含量分别在旱冬瓜+滇杨林和云南松林中最高,有机质在旱冬瓜+云南松+华山松林中最丰富。方差分析结果表明,以上各群落土壤中除云南松林和旱冬瓜+云南松+华山松林全氮含量差异不显著外,同类养分含量最高者均与其余群落存在显著差异(P<0.05)。各群落乔木和灌、草年均生物量统计结果表明(表3),旱冬瓜+云南松+华山松林的乔木和灌草本生物量均显著高于其余各群落(P<0.05)。总体上看,土壤养分含量在研究区各群落中并无明显规律,但林地生产力在旱冬瓜+云南松+华山松林中具有明显优势。
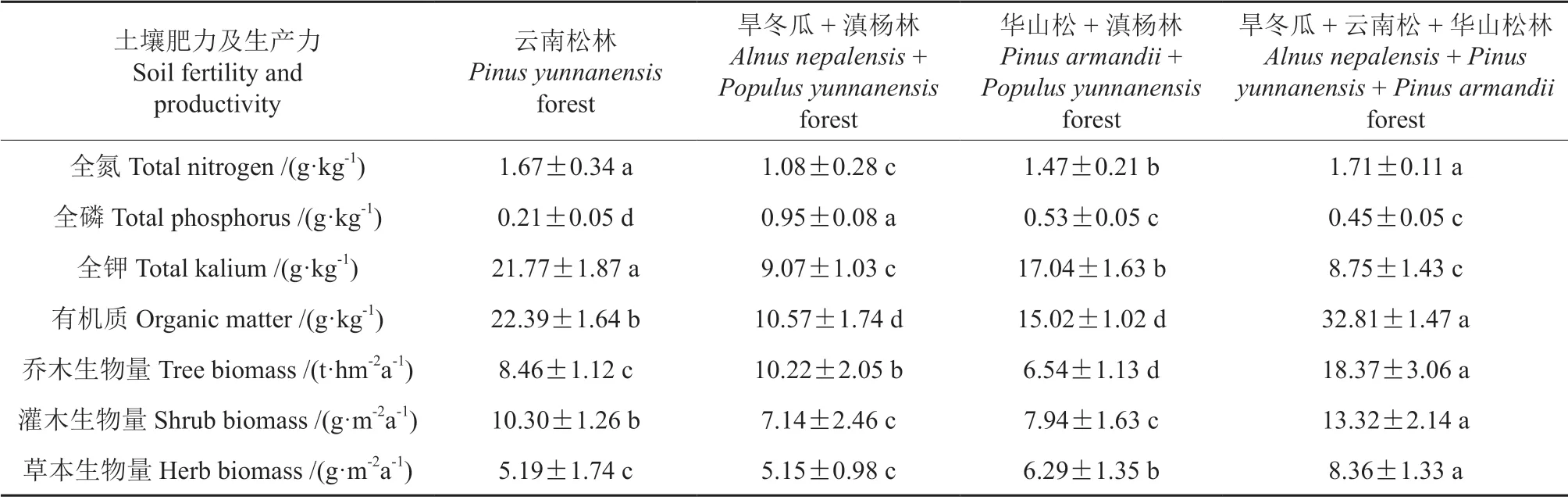
表3 4 种森林类型土壤肥力及林地生产力Table 3 Soil fertilizer and woodland productivity of four forest community types
3.5 外界干扰状况
研究区受外界干扰的形式多样,主要有林地侵蚀沟、人畜踩踏痕迹、放牧及野生动物活动等。野外调查中发现:在云南松林、旱冬瓜+滇杨林和华山松+滇杨林中林地表面均被流水侵蚀和部分踩踏痕迹,这势必影响到林下幼苗定居和灌草发育,尤其是在云南松林和华山松+滇杨林中影响更为明显;在旱冬瓜+云南松+华山松林中放牧活动和松鼠啃食松果,这对林下种子库存留和幼苗更新不利[10]。对上述4 种森林类型外界干扰状况赋值结果见表4。

表4 4 种森林类型中外界干扰形式及强度†Table 4 Forms and intensity of external disturbance in different community types
3.6 群落稳定性评价
将不同评价指标的同类因子数值合并累加,计算汇总结果见表5,并将其作为各评价指标的基础数据。
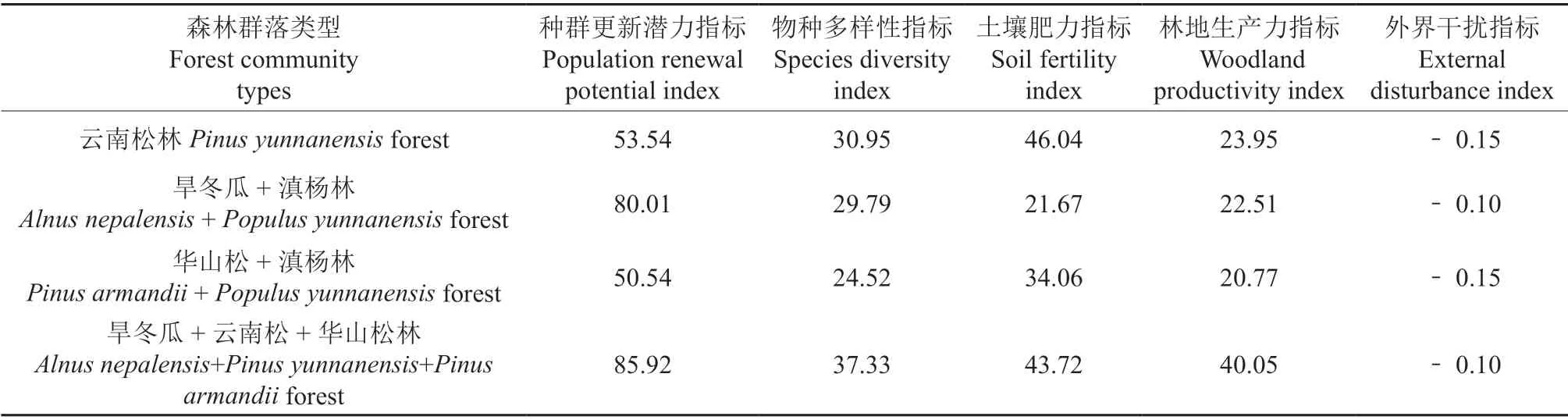
表5 不同群落类型5 项评价指标中各因子汇总值Table 5 Summary values of the five items factors of different community types
对评价指标体系中的5 项指标值进行标准化处理,结果见表6。
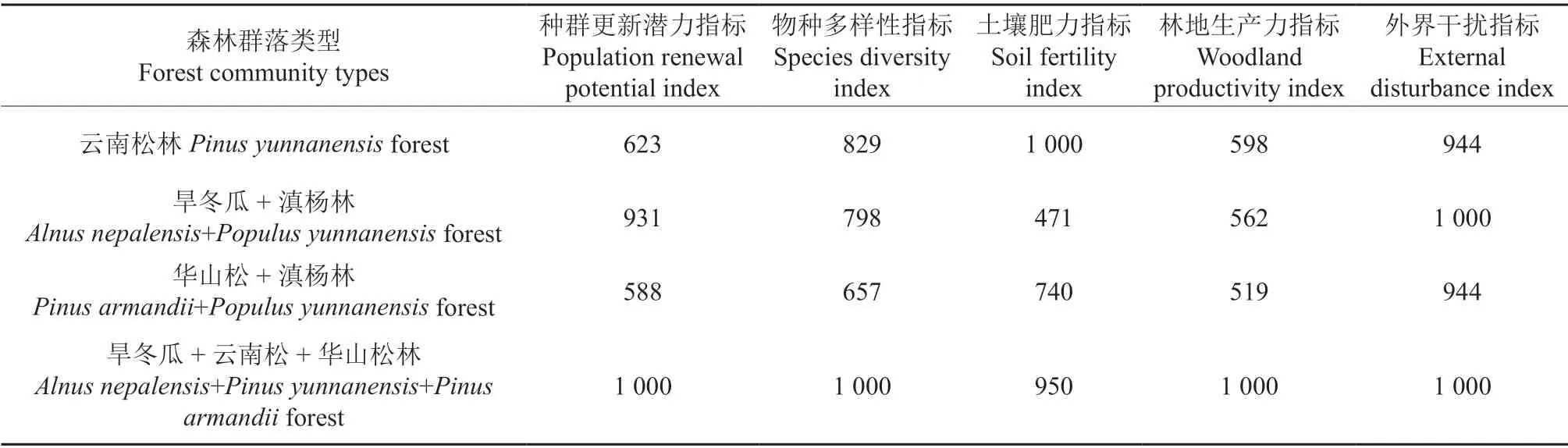
表6 不同群落类型5 项评价指标各因子标准化值Table 6 Standard index of the five items factors of different community types
根据4 种森林类型5 项指标标准化值计算隶属函数值,然后计算出各森林类型中5 项指标的加权隶属函数值,最后根据各森林类型评价指标的加权隶属函数值的和进行排序并比较其稳定性。
由各森林类型稳定性评价结果(表7)可知,威宁草海国家级自然保护区4 种森林群落的稳定性排序表现为:旱冬瓜+云南松+华山松林>云南松林>旱冬瓜+滇杨林>华山松+滇杨林。
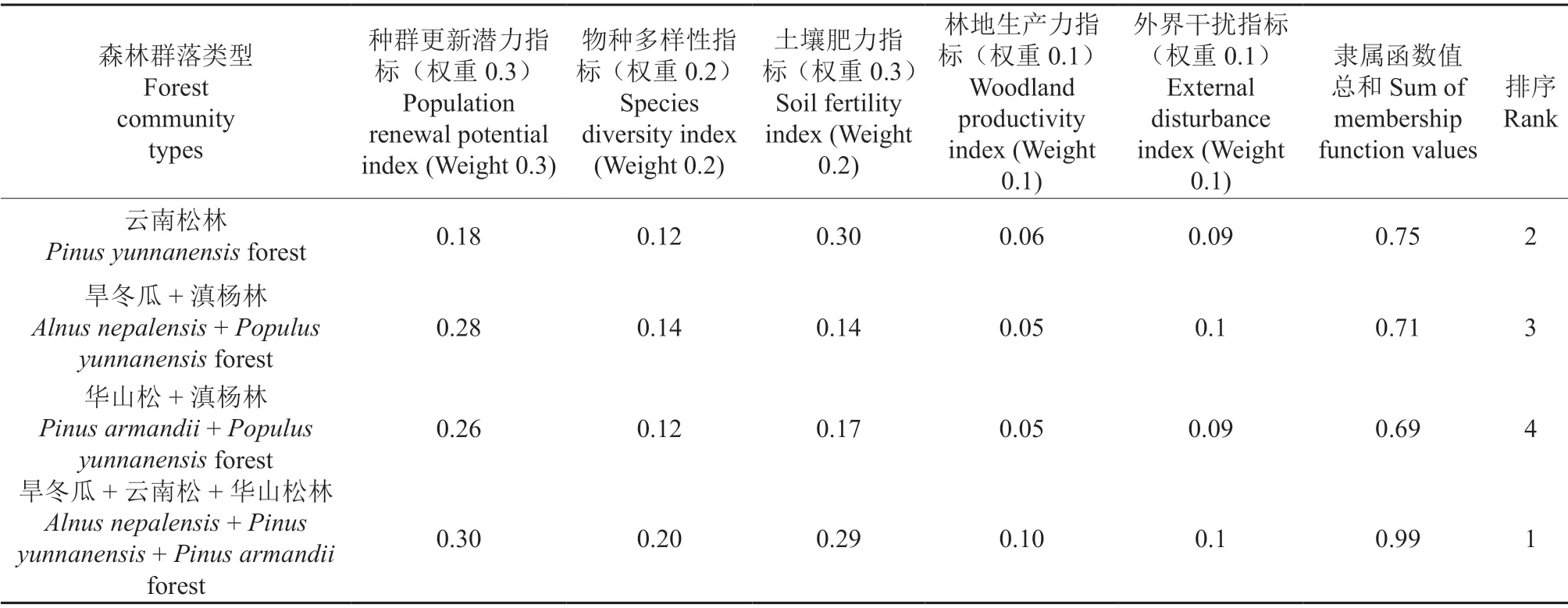
表7 不同群落类型指标加权隶属函数值及稳定性评价Table 7 Weighted subordination function value and stability evaluation of different community types
4 结论与讨论
本研究中,由旱冬瓜+云南松+华山松林组成的针阔多树种混交林具有较强的更新潜力,林中灌木和草本层物种丰富且多样性指数高,同时较高的土壤养分和林地生产力,使得该群落处于较为稳定的发育阶段。在云南松林中,种群更新潜力及物种多样性均较高,虽林地生产力指标偏低,但土壤肥力指标仍较高。在旱冬瓜+滇杨林中,由阔叶树种混交形成的林分中种群更新潜力较强,但林地生产力较低且土壤肥力差。在华山松+滇杨林组成的针阔混交林,因林地生产力较差,加上不利扰动,其群落稳定性相对最差。根据加权的隶属函数值综合评价草海国家自然保护区4 种森林类型稳定性,其排序依次为:旱冬瓜+云南松+华山松林>云南松林>旱冬瓜+滇杨林>华山松+滇杨林。
草海国家级自然保护区森林群落在物种组成、生境条件和林分发育状况均存在明显差异。本研究中,云南松林林相整齐,乔木层树种单一,是当地重要的森林类型之一;其余3 种林分类型主要位于草海水域周边的丘陵坡地中,是当地林业部门在20世纪70—80年代营造的滇杨林和旱冬瓜林,后又在林中补植了部分云南松和华山松,后随着乔木层植株生长和林下灌草发育,逐渐分化形成旱冬瓜+滇杨林、华山松+滇杨林和旱冬瓜+云南松+华山松林,这说明在研究区混交林较纯林具有更强的生存优势。森林群落类型的形成主要是物种对生境及当地气候长期适应的结果,也是森林生态系统自然演化的体现[31]。云南松、旱冬瓜和滇杨等都是分布在我国西南高原山地的广布树种,具有较强的耐瘠薄土壤、耐低温和喜光的乔木树种,能以建群种的形式构建发育良好的森林群落。因此,探究草海国家自然保护区森林群落物种特性及生长环境,不仅有助于明确当地森林的层次结构与物种配比,而且可为保护区植被恢复与重建提供基础资料。
稳定性是森林群落的综合性特征,是由群落更新潜力、物种多样性、林地承载力及外界干扰等因素共同决定[9]。乔木层作为森林生态系统的主体,其稳定存在对于保持群落概貌、维持林下生境和抵御外界不利干扰等方面均有重要作用。在旱冬瓜+云南松+华山松林中,乔木树种有足量的幼苗、幼树,是群落保持稳定的重要保障。较高的物种多样性对于增强森林群落的御逆性、自我恢复力及养分循环等均有显著积极作用,有助于群落长期稳定存在[5,10]。如土壤中的氮、磷、钾及有机质含量是影响林地生产力的最直接因素,最终表现为最高的乔木和灌草生物量[32]。云南松林中土壤较高的养分含量可为林中植物生长提供较好的营养供给,形成较高的林地生产力,使得群落具有较强的稳定性,而旱冬瓜+滇杨林因土壤养分含量低造成林地生产力较低,群落稳定性相对不高。林下扰动,如地表侵蚀、放牧、人为踩踏等都对林下幼苗、幼树及灌、草层植物发育造成危害,不利于森林群落的稳定性存在,这也是华山松+滇杨林稳定性较差的主要原因。
综上分析,针对草海国家保护区的云南松林和旱冬瓜+云南松+华山松林,在维持当前群落结构和生境条件即可长期稳定存在;旱冬瓜+滇杨林可通过人工措施提高土壤养分供给来提高林地生产力;华山松+滇杨林需加强管护措施,避免不利干扰,为植物生长营造有利的生存环境,促使群落生产力提高以维持其稳定存在。
猜你喜欢
科海故事博览(2022年28期)2022-11-25 03:09:48
电影文学(2021年21期)2021-11-14 21:01:24
林产工业(2020年8期)2020-08-31 06:58:26
中国森林病虫(2020年1期)2020-01-18 07:21:52
湛江文学(2019年2期)2019-11-13 19:54:18
金山(2019年9期)2019-10-18 09:37:14
乡村科技(2019年9期)2019-02-22 06:09:22
山西农业科学(2019年10期)2019-02-12 04:15:51
新闻传播(2018年15期)2018-09-18 03:20:12
戏曲研究(2017年4期)2017-05-31 07:39:32
