
龙竹人工林的含水率分析及地上生物量回归模型构建
2021-12-11 04:36:48王柯人罗文秀舒清态罗洪斌赖虹燕赵洪莹
西南林业大学学报 2021年6期
王柯人 罗文秀 舒清态 罗洪斌 赖虹燕 山 丹 王 强 赵洪莹
(西南林业大学林学院,云南 昆明 650233)
龙竹(Dendrocalamus giganteus)为禾本科(Gramineae)竹亚科(Bambusoideae)牡竹属(Dendrocalamus),分布于云南南部,包括红河州、玉溪市、保山市等地。除此之外,龙竹在缅甸、泰国、越南也有分布,其适应生长海拔为1 000~2 200 m[1]。龙竹因材质耐久性好,杆型通直、生长周期短、生产量较高等特点被广泛用于竹产区房屋的建造以及竹类产品和工艺品的编制[2]等方面;同时,龙竹还是很好的纸浆用材[3]。目前,龙竹在云南地区已被大面积栽培[4-6]。龙竹作为我国重要的森林资源,有着不可代替的作用,是解决木材供需矛盾的关键。2001年张齐生教授最先提出“以竹代木”和“以竹胜木”的理论[7]。龙竹不仅具有速生丰产、用途广泛等经济价值,龙竹林作为我国重要的森林资源之一,在固碳减排和促进山区经济发展等方面也有着重要的生态和社会价值[8]。在全球气候变化背景下,森林碳汇的计量和监测是当前研究的核心内容之一。
估算碳库和森林经营管理的关键在于构建适用于某一研究区域尺度的单木生物量模型,从而估测不同时期的森林生物量[9-11];同时,单株生物量模型的建立也是进行区域尺度森林生物量监测的基础。目前,国内外对龙竹单株生物量模型构建的研究较少,姚文静等[12]采用Logistic模型和多项式函数对淡竹(Phyllostachys glauca)生物量进行数学模型拟合,分析了多个构件在不同发育阶段的生物量变化、生物量因子之间的相互关系及生物量分配;郭子武等[13]研究了钩梢对雷竹(Phyllostachys praecox)叶生物量及其分配比例、出叶强度及异速生长均有明显的负面影响,不利于雷竹林光合能力的发挥。唐建维等[14]对西双版纳地区不同龄级人工龙竹的生物量及其模型进行了研究,模型精度较高,各器官及总生物量模型决定系数均在0.8以上,但所用的建模样本数量较少,各龄级的株数均小于30株,未达到小样本数量的要求,其生物量模型可推广性较弱;付小勇等[15]使用108株龙竹样竹建立了德宏州龙竹单株生物量模型,虽然建模样本数量有一定提高,但并未对不同龄级以及各器官生及物量特征进行分析,不同龄级的龙竹生物量变化并不相同,例如1年生的龙竹没有枝和叶,生物量主要集中在杆,因此对龙竹生生物量模型研究时时,需考虑对1年生和多年生进行划分。曾伟生[16]根据生物量建模标准指出样本数据的采集是建立生物量模型的基础,每个建模总体的样本数应大于150个,并且达到10个径阶,且考虑5 cm以下幼树生物量的测定。Huy等[17]在越南中部的17个样地通过收获法获取了83个竹杆地上生物量及其组件的样本,验证了可同时估算地上生物量及其组分的建模系统。Macdicken[18]研究表明如果生物量模型只应用于某一特定区域,建模样本数最少12个,若要建立一个区域性生物量模型样本数需30~100个,样本径阶分配均匀则至少需30个。综上可知,现阶段龙竹单株地上生物量模型有待进一步研究。本研究以大型丛生人工龙竹为研究对象,采伐不同龄级、不同径阶的龙竹样本151株,对龙竹地上部分各器官生物量配比及含水率分析,并建立大型丛生人工龙竹地上部分各器官及总生物量模型,为大型丛生人工龙竹林地上生物量的估算及经营管理提供参考。
1 研究区概况
研究区位于云南省临沧市沧源县,地处东经98°52′~99°43′,北纬23°04′~22°40′,该区位于横断山脉南部帚形扩大部位。其地势中部和北部高,东、西、南三面低,海拔为800~ 2000 m。由于陆地的抬升运动以及流水侵蚀还有河流的切割作用,形成了山峦起伏的谷地相间地貌。由于地处低纬度地区,气候以热带、亚热带气候为主,立体气候明显,全境气候温暖,干湿两季显著,极端最低气温1.3 ℃,极端最高气温33.8 ℃,多年平均气温18.5 ℃,日照时数2 110.7 h,年均降雨量1 337 mm。在复杂的地形地势和多样的气候条件,形成了种类繁多、类型多样的植被特点。全县森林面积为179 421.1 hm2,主要植被类型有季雨林、常绿阔叶林、落叶阔叶林、暖性针叶林、竹林、灌丛等。
2 研究方法
2.1 标准竹的选择及样本采集
样竹数据采集于2018年11月—12月在云南省临沧市沧源县进行[19],具体地理位置和实测样点的空间分布见图1。根据沧源县2017年森林资源二类调查数据,结合实地调查,在龙竹分布范围的34个样地内随机选取不同龄级、不同径阶、生长良好且无病虫害的龙竹151株作为标准样竹。其中:I龄级(竹龄≤1 a)31株,Ⅱ龄级(1 a<竹龄≤3 a)58株,Ⅲ龄级(竹龄>3 a)62株,龄级的划分参考唐建维等[14]以及林场管理人员多年管理经验。由于1年生龙竹无枝、叶,仅有竹杆,所以还可分为1年生(竹龄≤1 a)龙竹31株和多年生(不分竹龄)龙竹151株。
图1 实测样点的空间分布Fig. 1 Spatial distribution of measured sample points
2.2 样竹各指标的测定
将选取的样竹从基部砍伐,测量其地径、胸径(DBH)、地壁厚、胸壁厚、胸径节长和竹高,并对竹杆、枝、叶进行取样,各器官分别获取200~300 g。竹杆样本分别在竹杆的1/4、1/2、3/4处截取,获取长度约3 cm的竹杆圆环,称其鲜质量。竹枝和竹叶的样本分别从样竹上、中、下3个部位获取,选取不同粗细程度的枝条和不同大小的叶片。将采集的样品放入烘箱内,先将竹杆置于105 ℃下杀青2 h,再在85 ℃恒温下烘10 h;枝、叶样本先置于75 ℃恒温下烘8 h,然后冷却至室温称质量[19-20],计算出各器官的含水率;再根据含水率计算样竹的各器官干质量和总干质量。龙竹各项测量指标见表1。
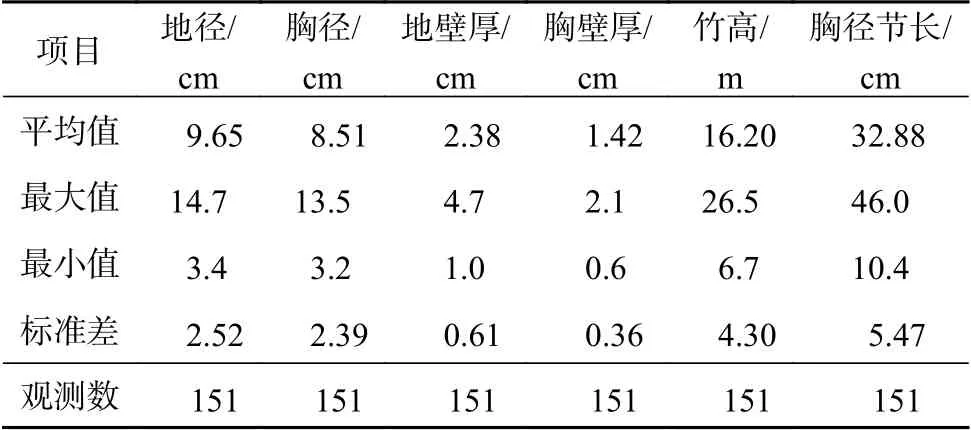
表1 龙竹各测量指标Table 1 D. giganteus various measurement indicators
2.3 建模方法
本研究以121株龙竹样本数据为建模样本,30株龙竹样本数据为检验样本,以DBH为自变量,各龄级杆生物量、枝生物量、叶生物量和总生物量数据为因变量,采用直线、幂函数、指数函数、多项式等对不同龄级的龙竹各器官生物量和单株地上生物量进行回归分析,根据模型评价指标,选择最优生物量模型。
2.4 分析方法
利用SPSS 22.0软件,采用皮尔逊(Pearson)相关性分析法进行显著性检验和相关性分析,以确定各器官生物量之间的相关性,选取相关系数最高的调查因子,建立龙竹各器官地上生物量和单株地上生物量的估测模型。采用决定系数(R2)对估测模型进行精度检验,精度决定系数的变化范围在0~1,值越大,表明自变量与因变量之间的相关性较好,生物量模型精度高。
3 结果与分析
3.1 龙竹各器官含水率分析
龙竹各器官组织结构和功能不相同,其含水率也存在一定差异。随着龙竹龄级的增长,各器官的含水率也会出现相应的变化;同时,龙竹各器官含水率的变化也从侧面反映出生物量的累积程度,不同龄级龙竹各器官含水率见表2。

表2 不同龄级龙竹各器官的含水率Table 2 Moisture content of different organs of D. giganteus at different ages %
由表2可知,Ⅰ到Ⅲ龄级龙竹随龄级的增加,各器官生物量的逐渐累积,含水率逐步下降,单株龙竹地上部分平均含水率分别为75.01%、49.35%、47.97%。从各器官含水率变化可知,Ⅰ龄级龙竹枝叶未长开且竹杆幼嫩生物量累积较少,Ⅱ和Ⅲ龄级竹杆含水率有极显著差异(P<0.01);其次,Ⅱ和Ⅲ龄级龙竹含水率呈杆>枝>叶的规律。
3.2 龙竹单株地上生物量分配占比
龙竹地上部分生物量由杆、枝、叶构成,根据不同龄级龙竹地上部分各器官生物量配比分析,结果见表3。
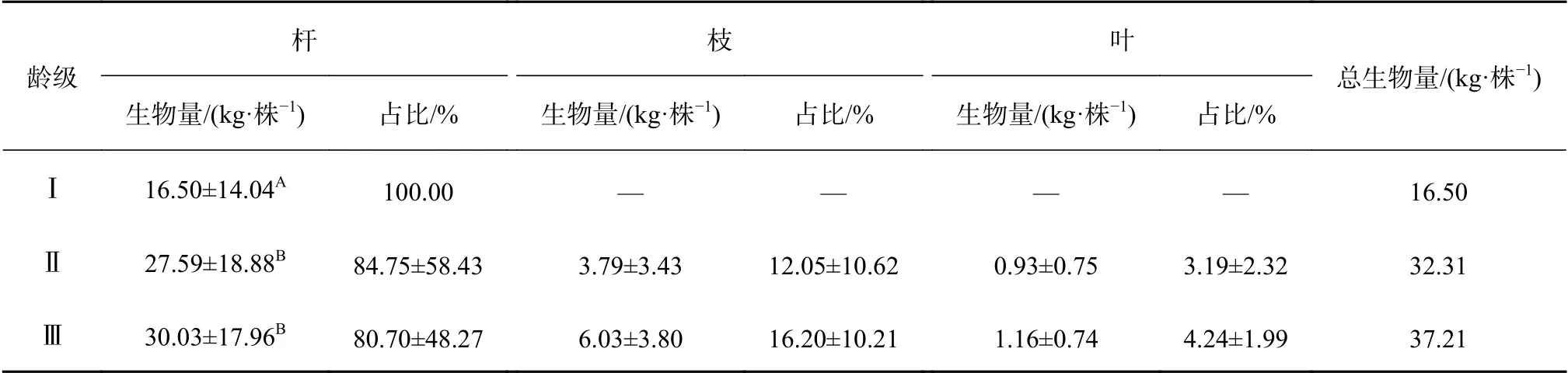
表3 不同龄级龙竹地上部分各器官生物量分配Table 3 Biomass allocation of various organs in the ground of different age classes
根据不同龄级龙竹地上部分生物量分配结果可知,Ⅰ到Ⅲ龄级龙竹的单株地上生物量平均值依次为l6.50、31.36、37.64 kg/株;Ⅰ龄级龙竹枝和叶未完全生长开,生物量集中在竹杆且生物量累积较少,因此生物量较小。从不同龄级生物量的变化可知,Ⅰ到Ⅲ龄级,随着龙竹的生长,龙竹杆、枝、叶的生物量和总生物量均随龄级的增加而增加;但从各器官生物量的占比来看,随着龙竹的生长,枝和叶逐渐繁茂,枝和叶生物量的配比也逐渐增加,而杆生物量的占比随着龄级的增加而降低。
3.3 单株龙竹地上生物量回归模型的构建
根据国内外已有生物量模型构建研究结果可知,乔木地上生物量与其胸径存在显著相关性,对于野外实际测量,乔木高度的测量结果通常存在较大的误差,所以对于生物量的估测来说,高度并不是较优的参数[21-22]。为避免因竹杆高度的引入带来的龙竹地上生物量模型构建误差以及提高在野外测量工作中的效率和可行性,不同品种竹类单株地上生物量模型的研究证明,采用以胸径为单变量的回归模型能够较好地反映竹类生物量随胸径的变化趋势,生物量模型均是基于胸径的单变量回归模型,且结果较好[22-25]。因此,本研究以龙竹的胸径(DBH)为单一变量,对龙竹各器官生物量进行相关性分析后,建立单株地上生物量回归模型。
3.3.1 相关性分析
为分析龙竹枝、叶生物量对构建地上生物量模型的影响,除了对不同龄级各器官进行生物量模型构建,还建立1年生(竹龄≤1 a)龙竹地上生物量模型和多年生(不分龄级)龙竹地上生物量模型。根据不同龄级龙竹地上部分各器官生物量计算结果,对龙竹的胸径(DBH)与各器官生物量进行相关性分析,结果见表4。
由相关性分析结果可知,Ⅰ龄级到Ⅲ龄级以及多年生的龙竹中,胸径(DBH)与各器官生物均具有良好的相关性,各龄级龙竹胸径与杆生物量、枝生物量、叶生物量和地上生物量的相关系数均在0.75~0.93,且达到极显著水平(P<0.01)。其中,胸径与杆生物量和总地上生物量的相关系数均在0.92以上,而胸径与枝和叶生物量的相关性较低,相关系数为0.73~0.82,但也达到了极显著水平(P<0.01)。1年生龙竹胸径与地上生物量相关系数为0.930,多年生龙竹胸径与地上生物量相系数为0.933,均具有良好的相关性。
3.3.2 地上生物量模型的构建
以121株龙竹样本胸径为自变量,各龄级杆生物量、枝生物量、叶生物量和总生物量数据为因变量,构建龙竹地上生物量模型,分别绘制龙竹的胸径-杆生物量、胸径-枝生物量、胸径-叶生物量、胸径-地上生物量散点图,见图2。结合二倍标准差法、三倍标准差法进行异常数据的判断和剔除。结果表明,本次调查采集的龙竹样本无异常数据。
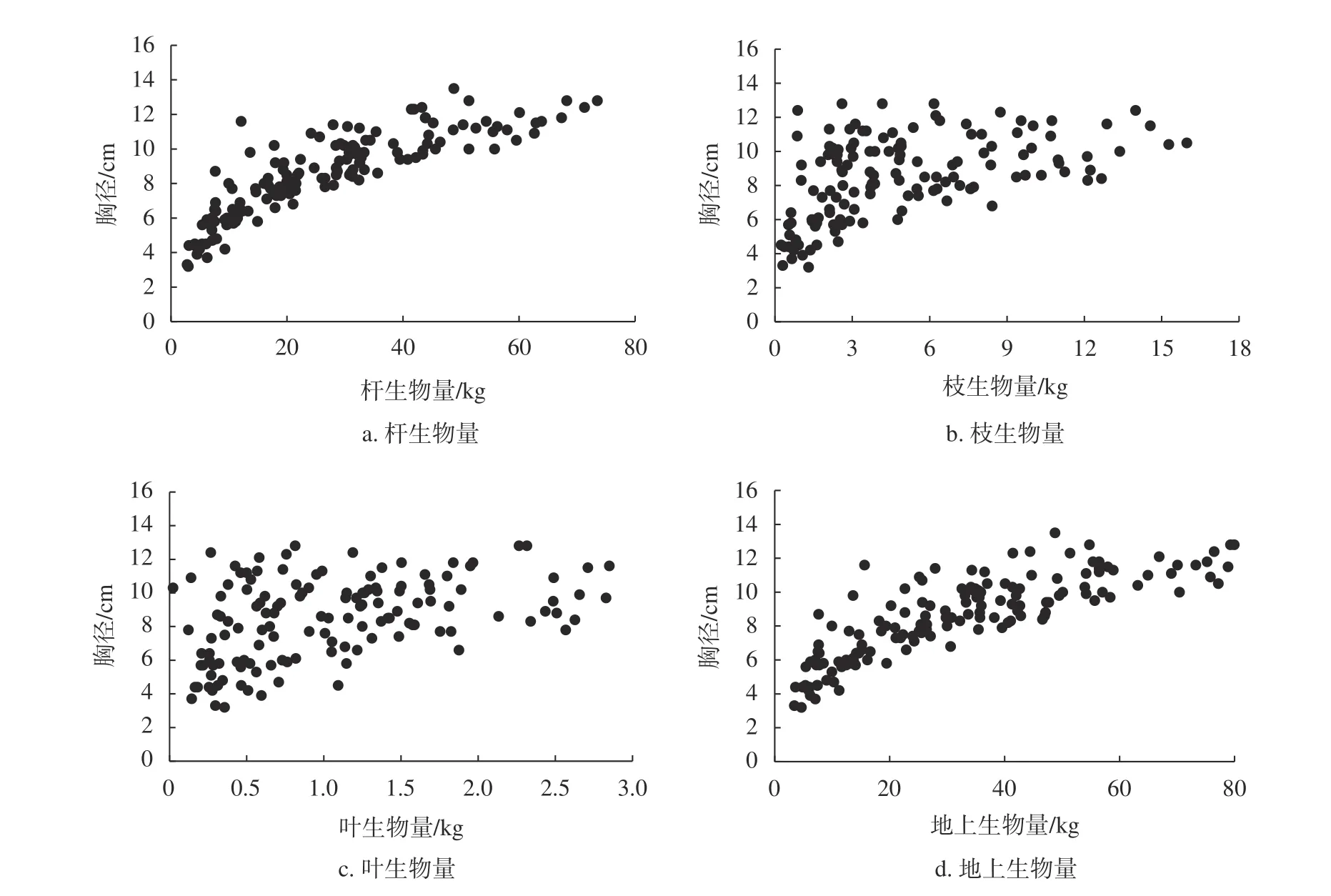
图2 胸径与各器官生物量建模样本散点图Fig. 2 Scatter plot of the modeling sample of DBH and biomass of organs
根据不同龄级胸径(DBH)与各器官生物量相关性分析结果,以胸径(DBH)为自变量,分别以龙竹的杆生物量、枝生物量、叶生物量和地上生物量为因变量,参考相关研究[22-23],建立不同龄级龙竹生物量估测模型,其中以幂函数拟合效果最佳,模型拟合结果见表5。
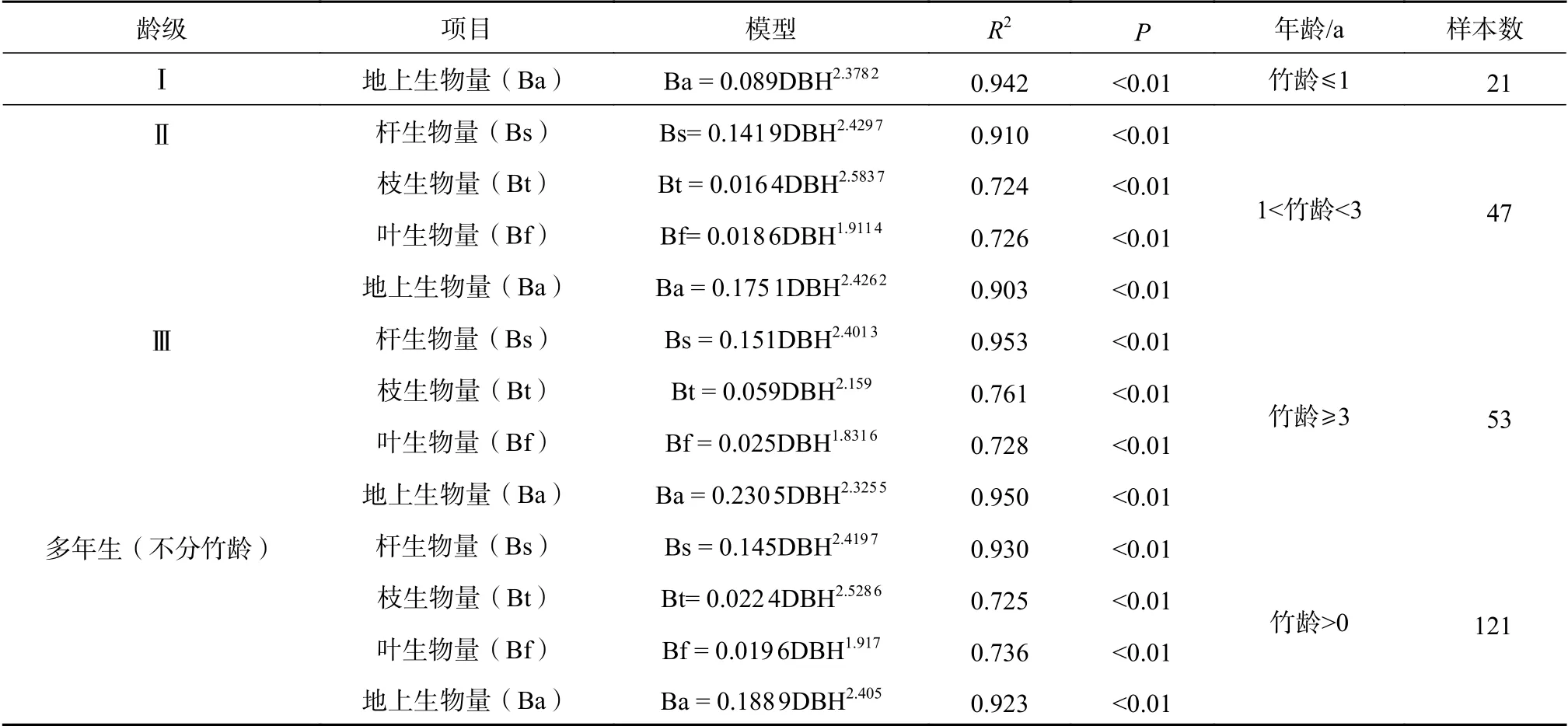
表5 不同龄级各器官及总生物量模型Table 5 Different age organs and total biomass models
从生物量模型的拟合结果来看,Ⅰ龄级到Ⅲ龄级以及多年生的地上龙竹生物量模型决定系数均在0.90以上。其中,杆生物量模型和地上生物量模型决定系数均在0.92以上,而枝和叶生物量模型决定系数较低,决定系数为0.72~0.76,但模型显著均小于0.01,此结果与相关性分析结果一致。1年生龙竹地上生物量模型R2为0.942,多年生龙竹地上生物量模型R2为0.923,相比1年生龙竹地上生物量模型决定系数较低,可知多年生龙竹地上生物量模型精度受Ⅱ、Ⅲ龄级龙竹枝、叶生物量影响有所降低。杆生物量模型和总生物量模型精度较高,一方面由于杆的测量误差比枝和叶的误差小,另一方面结合生物量配比分析结果可知,多年生龙竹枝和叶的占比均在15%~20%,因此枝和叶的误差对单株龙竹地上生物量的影响较小,所以龙竹总生物量模型精度较高。
4 结论与讨论
本研究以大型丛生人工龙竹单株地上生物量为研究对象,采伐不同龄级、不同径阶、生长良好且无病虫害的151株龙竹样本,对龙竹地上部分各器官的含水率、生物量配比及生物量模型进行研究。得到以下结论:
1)Ⅰ龄级到Ⅲ龄级随着龄级的增加各器官生物量的逐渐积累,含水率逐渐下降。Ⅰ龄级龙竹杆含水率最高,与Ⅱ和Ⅲ龄级杆含水率有极显著差异;Ⅱ龄级和Ⅲ龄级中含水率呈现出杆>枝>叶的变化规律。
2)随着龄级的增加各器官生物量的逐渐积累,Ⅰ龄级龙竹枝和叶未完全长开,生物量集中在竹杆;随着龙竹的生长,龙竹杆、枝、叶的生物量和地上生物量均随着龄级的增加而增加,枝和叶生物量的配比也逐渐增加,而杆生物量的占比随龄级的增加而降低。
3)各龄级龙竹胸径与杆生物量、枝生物量、叶生物量和总生物量的有着良好的相关性且达到极显著水平,以胸径为自变量建立的幂函数单株生物量模型决定系数均在0.9以上。Ⅱ龄级和Ⅲ龄级枝和叶生物量模型决定系数相较偏低,均在0.7~0.8,但模型显著性(P)均小于0.01,表明生物量模型具有较好的可信度,同时也有着较强的适用性,可用于相似生长条件下的龙竹生物量估测。
单株龙竹地上部分生物学特征分析和生物量模型的构建是进行区域尺度龙竹生物量估算和龙竹经营管理的基础,从生物量模型的拟合结果来看,总生物量模型的决定系数均在0.92以上,各龄级的龙竹样本株数均大于30株,龙竹样本总株数大于150株,生物量模型有着较好的普适性。研究结果与唐建维等[14]和付小勇等[15]所研究得到的结果差异较小,但枝和叶生物量模型对比唐建维等[14]的研究结果可知,本次所建立的枝和叶生物量模型精度偏低,其原因可能为本研究所获取样本数量较少,且龙竹枝和叶的样本收集均在样竹伐倒之后进行采集,此过程中造成了枝和叶的部分损失。在今后的研究中,龙竹数据采集和生物量估测应用于其他地区的植被,研究方法的普适性需做进一步探索,确保生物量估算达到预期效果。
猜你喜欢
湖南生态科学学报(2022年4期)2022-11-11 03:44:34
散文诗世界(2022年5期)2022-05-23 07:12:21
电脑与电信(2021年10期)2021-02-10 06:53:44
陕西煤炭(2020年4期)2020-08-05 14:34:50
南方农业学报(2020年4期)2020-06-04 15:51:13
河南农业大学学报(2020年2期)2020-05-22 02:56:30
南方农业学报(2020年10期)2020-01-21 15:36:41
山西林业科技(2019年1期)2019-05-27 12:24:52
科学与财富(2018年12期)2018-06-11 01:49:24
阅读与作文(小学高年级版)(2016年2期)2016-03-04 22:21:18
