
绿盲蝽G蛋白偶联受体激酶2基因(AlGRK2)的表达分析及在绿盲蝽生长发育中的功能
2021-12-07 05:43谭永安姜义平赵静肖留斌
中国农业科学 2021年22期
谭永安,姜义平,赵静,肖留斌
绿盲蝽G蛋白偶联受体激酶2基因()的表达分析及在绿盲蝽生长发育中的功能
谭永安,姜义平,赵静,肖留斌
江苏省农业科学院植物保护研究所,南京 210014
【目的】克隆绿盲蝽()G蛋白偶联受体激酶2基因()cDNA序列,明确其时空表达谱,阐明外源蜕皮激素(20E)对表达的影响,分析在绿盲蝽生长发育中的作用,为进一步研究其在蜕皮激素信号转导通路中的功能打下基础。【方法】RACE法克隆获得全长,实时荧光定量PCR(qRT-PCR)分析不同日龄绿盲蝽及雌成虫不同组织中的表达谱,分析外源20E诱导及RNAi处理后,mRNA表达的应答反应及对绿盲蝽生长发育主要参数(发育历期、若虫体重及成虫羽化率)的影响。【结果】cDNA序列全长2 715 bp,开放阅读框2 106 bp,编码701个氨基酸,ExPASy预测其蛋白分子量为80.2 kD,理论等电点为6.56;蛋白结构分析显示AlGRK2包含4个结构域,即G蛋白信号调节区(RGS,54—175 aa)、丝氨酸/苏氨酸激酶结构域(S-TKc,191—454 aa)、丝氨酸/苏氨酸型蛋白激酶的伸展部分(S-TK-X,455—534 aa)和PH结构域(PH,558—655 aa),其中PH结构域是GRK2蛋白的典型结构域;系统发育分析结果表明,绿盲蝽GRK2与茶翅蝽GRK2亲缘关系最近;在绿盲蝽1—16日龄虫体内均有表达,mRNA表达量呈现出波动式下降的模式,在绿盲蝽初始龄期的表达量较高,而在末龄期的表达量显著下降;在绿盲蝽雌成虫卵巢和脂肪体中高表达,在胸与足中的表达量较低;外源20E处理后,在绿盲蝽1日龄和3日龄表达量显著下调,在雌成虫各组织中均表达上调,在卵巢及脂肪体中上调幅度最大,相反的是,20E信号通路中PLC抑制剂U73122处理的表达量下调;绿盲蝽若虫发育历期、末龄若虫体重和成虫羽化率均显著下降,相反的是,U73122处理组若虫期的发育历期显著延长;此外,与注射ds处理组相比,注射ds处理后绿盲蝽的表达水平显著下降,若虫死亡率及发育历期显著增加,而成虫羽化率和5龄若虫体重均显著下降。【结论】在绿盲蝽体内的表达谱显示出发育阶段特异性和组织特异性;外源20E抑制剂及RNAi处理后,均可抑制的表达,同时还可对绿盲蝽生长发育产生不利影响,表现为延缓绿盲蝽的发育进度、降低5龄若虫体重及成虫羽化率。
绿盲蝽;G蛋白偶联受体激酶2;蜕皮激素;基因克隆;表达谱;RNA干扰
0 引言
【研究意义】绿盲蝽()属半翅目盲蝽科,是果树、蔬菜及棉花等多种经济作物上的重要刺吸类害虫[1]。近年来,随着我国农业产业结构的调整及化学农药使用等因素,绿盲蝽在多种作物上危害趋势加重[2-3],造成巨大的经济和生态效益的损失。因此,有效防控绿盲蝽亟待开拓新型无公害防控手段。蜕皮激素(20-hydroxyecdysone,20E)可调控昆虫多种重要的生理活动,挖掘蜕皮激素信号转导中的关键基因,将有利于开发新型绿色昆虫生长调节剂及有效控制绿盲蝽危害。【前人研究进展】20E是一种类固醇激素,主导调控昆虫发育、变态及繁殖等重要生理活动。蜕皮激素信号转导途径一直是昆虫学研究的重点和热点。研究表明,昆虫蜕皮激素转导分为基因组途径及非基因组途径。目前已经明确了基因组转导的途径:20E依靠自身脂溶性穿过细胞膜进入细胞核,激活核受体基因蜕皮激素受体基因(ecdysone receptor,EcR)和超气门蛋白(ultraspiracle protein,USP)形成异源二聚体,启动下游基因的转录,进而通过核受体途径调控昆虫生长发育及繁殖[4-6]。20E转导还存在着多种非基因组途径。其中最重要的一种是20E通过细胞膜上的G蛋白偶联受体(G protein-coupled receptor,GPCR),调控细胞内磷脂酶C(phospholipase C,PLC)、Ca2+、蛋白激酶C(protein kinase C,PKC)等因子的表达,进而触发基因组途径的启动,启动20E下游靶标基因的转录[7-8]。G蛋白偶联受体激酶(G protein-coupled receptor kinase,GRK)是一类重要的膜蛋白,属于丝氨酸/苏氨酸蛋白激酶家族成员,可磷酸化GPCR,磷酸化后的GPCR与-抑制蛋白(-arrestin)结合后可发生脱敏反应,导致G蛋白不能与GPCR结合,从而参与20E信号通路中一系列的生理活动[9-10]。GRK通常具有4个典型结构域:G蛋白信号调节域(regulator of G-protein signaling,RGS)、AGC激酶家族的Ser/Thr蛋白激酶结构域(Ser/ Thr protein kinase domain)、氨基端N域(N-terminal domain)和羧基端C域(C-terminal domain)[11-12]。此外,根据蛋白结构差异将GRK分为GRK1、GRK2和GRK4这3个亚家族,其中GRK1亚家族包含GRK1和GRK7,其蛋白C端可以发生法尼基化,可促进其与质膜结合[13];GRK2亚家族包含GRK2和GRK3,其蛋白序列C端存在一个PH结构域,可与酸性磷脂和G亚基相互作用,因可参与肾上腺素传导,又被称为-肾上腺素受体激酶家族[14-16];GRK4亚家族包含GRK4、GRK5和GRK6,其N-和C-末端存在两个多碱基区域,其中GRK4和GRK6蛋白C端可发生棕榈酰化[17-18],GRK5的C端含有碱性氨基酸富集区[9]。研究表明,昆虫的GRK2基因可参与激素信号通路的转导,进而调控昆虫的多种生理活动。在棉铃虫()中,20E通过调控GRK2由细胞质向细胞膜移动,随后磷酸化ErGRCR-2(ecdysone-responsible GPCR-2),并与之结合,诱导ErGRCR-2后发生内吞,最后可终止20E信号的转导[19];此外,在黑腹果蝇()中,表达量的减少会导致果蝇胚胎分化异常及成虫无法飞行等[20]。上述研究结果表明,GRK2可参与昆虫20E信号的转导,并在多种生理活动中发挥着重要功能。此外,在家蚕()中的研究也发现,当对幼虫注射20E后,发育受到阻遏,呈现出发育历期的缩短及死亡率增加[21];同样在荨麻蛱蝶()和孔雀蛱蝶()中也发现,20E也可显著降低幼虫体重及存活率[22]。【本研究切入点】目前,对的研究多集中在哺乳动物及某些模式昆虫上,对绿盲蝽()的相关研究尚未见报道。【拟解决的关键问题】获得全长,明确其时空表达动态,分析外源20E对其表达的影响,阐明在绿盲蝽生长发育中的功能。
1 材料与方法
试验于2019年5月至2021年3月在江苏省农业科学院植物保护研究所完成。
1.1 供试虫源
初始绿盲蝽采自2018年早春的江苏大丰及东台蚕豆()田,于室内用四季豆()豆荚继代饲养,饲养条件为温度(25±1)℃,相对湿度70%±5%,光周期12L﹕12D。
1.2 主要试剂和仪器
Trizol(Invitrogen,Rockville,USA);M-MLV反转录试剂盒(Promega,Madison,USA);PrimeScriptTMRT Master Mix、SMARTer®RACE 5′/3′ Kit、TB Green®TaqTm、pMD®19-T Vector(TaKaRa,Shiga,Japan);2×EXP Taq(Accurate Biotechnology,中国湖南);AxyPrepTMDNA Gel Extraction Kit、AxyPrepTMPlasmid Miniprep Kit(Axygen,Union City,CA,USA);T7 RiboMAXTMExpress RNAi System(Promega,Madison,Wisconsin,USA);20E、U73122(Sigma,Missouri,USA)。Allegra 21R台式高速冷冻离心机(Beckman,California,USA);水平电泳系统(Bio-Rad,California,USA);LightCycler 480 II(Roche,Basel,Switzerland);Eppendorf FemtoJet 4i-TransferMan 4r微量自动注射仪(Eppendorf,Hamburg,Germany)。
1.3 AlGRK2克隆
取绿盲蝽3龄若虫20头,Trizol法提取绿盲蝽总RNA,M-MLV反转录合成cDNA。基于NCBI报道的已知昆虫,设计简并引物(-F和-R)(表1),获得保守序列,进一步根据获得的序列设计特异性引物(表1),进行5′和3′端序列的克隆。5′端序列克隆反应体系:10×PCR缓冲液5.0 μL,25 mmol·L-1MgCl23.0 μL,10 mmol·L-1dNTP mix 1.0 μL,10 μmol·L-15′--F 1.0 μL,10 μmol·L-15′--R 1.0 μL,cDNA 2.5 μL,Taq DNA Polymerase 0.5 μL,超纯水补足至20.0 μL;PCR反应条件:94℃ 2 min,94℃ 30 s,55℃ 30 s,72℃ 1 min,循环35次,94℃延伸7 min。3′端序列克隆反应体系:PCR-Grade Water 34.5 μL,10×Advantage PCR Buffer 5.0 μL,10 mmol·L-1dNTP Mix 1.0 μL,50×Advantage Polymerase Mix 1.0 μL,cDNA 2.5 μL,10 µmol·L-13′--F 1.0 μL,10 µmol·L-13′--R 1.0 μL,超纯水补足至50.0 μL;PCR反应条件:94℃ 2 min,94℃ 30 s,68℃ 30 s,72℃ 1 min,循环30次,72℃延伸7 min。将获得的5′和3′端及保守区域进行测序和拼接(上海生工生物工程有限公司),最终获得全长,最后对全长基因进行克隆验证。
1.4 AlGRK2序列分析与进化树构建
采用Expasy在线分析程序(http://web.expasy.org/ compute_pi/)预测核苷酸序列的基本物理性质;SMART(http://smart.embl-heidelberg.de/)在线预测蛋白结构域;利用MEGA6.0软件构建系统进化树(neighbor-joining,NJ)。

表1 本研究中所使用的引物
下划线处为T7-RNA聚合酶启动子序列 T7-RNA polymerase promoter is underlined
1.5 AlGRK2时空表达谱
取自然条件下1—16日龄绿盲蝽虫体及羽化8 d后绿盲蝽雌成虫的7个不同组织(头、胸、翅、足、中肠、卵巢和脂肪体),作为时空表达谱分析的样本。各日龄样本每个处理15头绿盲蝽,各组织样本每个处理10头绿盲蝽,4次生物学重复。液氮处理样品后-80℃保存备用。Trizol法提取总RNA,Prime ScriptTMRT Master Mix反转录合成cDNA,-20℃保存。qRT-PCR试验步骤按照SYBR Premix Ex Taq Kit说明书进行。所用的-QF及-QR引物参见表1,并以绿盲蝽持家基因为内标基因(GenBank登录号:JN616391)。扩增体系:2×TB Green Premix Ex Taq II 10 μL,上下游引物各0.4 μL,cDNA 2.0 μL,ddH2O 7.2 μL,总体积20.0 μL。反应条件:95℃ 30 s;95℃ 5 s,60℃ 20 s,72℃ 10 s,40个循环。
1.6 不同药剂处理后AlGRK2表达量分析
分别用20E(2 μmol·L-1)、U73122(20E信号通路中PLC抑制剂[23],50 μmol·L-1)、20E+U73122处理、乙醇(CK)浸泡切除两端的四季豆1 min,晾干后饲喂初孵1龄若虫。取不同处理的1—6日龄绿盲蝽若虫及上述1.5中不同雌成虫组织样本,作为表达特性分析的样本。各日龄样本每个处理15头绿盲蝽,各组织样本每个处理10头绿盲蝽,4次生物学重复。表达量测定方法同1.5。
1.7 绿盲蝽若虫发育历期、5龄若虫体重及成虫羽化率
将上述不同处理后的初孵绿盲蝽若虫置于一次性圆形餐盒(15 cm×6.5 cm)内,用一次性纱布封顶。逐日记录若虫发育进度及成虫羽化率,每处理50头绿盲蝽若虫,4次重复。计重绿盲蝽5龄末期若虫(出现翅芽)体重,每处理10头若虫,4次重复。
1.8 dsAlGRK2的合成与注射
以测序验证正确的质粒作为DNA模板,利用带T7启动子的引物(-T7F、-T7R,表1)分别进行PCR产物正义链与反义链的扩增。用T7 RiboMAXTMExpress RNAi System合成试剂盒合成双链RNA(ds)和GFP双链RNA(ds),最后用无RNA酶水配制成1 μg·μL-1终浓度的溶液备用。于绿盲蝽初孵4龄若虫的后足与胸的连接处进行注射,每头若虫dsRNA注射量为1 μg。设未注射及注射ds为对照组。
1.9 RNAi处理后AlGRK2的表达及相关生物学参数分析
分别于注射后6、12、24和48 h收集上述不同处理的初孵4龄绿盲蝽若虫,每个处理60头,4次重复。参照1.5中的方法分析表达量。此外,将不同处理后的初孵4龄绿盲蝽若虫置于含有新鲜四季豆的一次性圆形餐盒(15 cm×6.5 cm)中,逐日统计死亡率、羽化率及5龄若虫发育历期,并计重5龄末期若虫(出现翅芽)体重,每个处理10头若虫,4次重复。
1.10 数据分析
相对表达量变化均采用2-ΔΔCt计算[24]。生物学相关参数的差异显著性统计分析均采用统计软件SAS 8.0 完成。
2 结果
2.1 AlGRK2 cDNA序列克隆
利用RACE法获得了全长cDNA序列(GenBank登录号:MN514868)。序列分析结果表明,cDNA序列全长2 715 bp,开放阅读框2 106 bp,编码701个氨基酸。ExPASy预测其蛋白分子量为80.2 kD,理论等电点为6.56。SMART在线预测AlGRK2蛋白包含4个结构域:G蛋白信号调节区(RGS,54—175 aa)、丝氨酸/苏氨酸激酶结构域(S-TKc,191—454 aa)、丝氨酸/苏氨酸型蛋白激酶的伸展部分(S-TK-X,455—534 aa)和PH结构域(PH,558—655 aa),其中PH结构域具有与肌醇磷酸酯和蛋白质结合的能力(图1),此为GRK2蛋白的典型特征,表明获得的基因是亚型基因。

绿色、红色、黄色和蓝色阴影标注的氨基酸分别代表RGS结构域、S-TKc结构域、S-TK-X结构域和PH结构域
2.2 AlGRK2系统发育
对核苷酸推导出的氨基酸序列进行Blast比对。结果表明,AlGRK2氨基酸序列与半翅目蝽科茶翅蝽()GRK 2 isoform X2基因序列同源性最高,为92.3%,表明克隆获得的基因为2。利用MEGA6.0软件中的NJ法对包括绿盲蝽在内的18种不同种类昆虫的GRK2氨基酸序列构建系统进化树(图2),结果显示,绿盲蝽GRK2与茶翅蝽GRK2亲缘关系最近。
2.3 AlGRK2时空表达特性
在1—16日龄绿盲蝽中均有表达,mRNA表达量呈现出波动式下降的模式,以1日龄绿盲蝽表达量最高(图3-A,<0.05)。综合来看,在绿盲蝽初始龄期的表达量较高,而在末龄期的表达量显著下降。在绿盲蝽8日龄雌成虫卵巢和脂肪体中高表达,而在胸和足中微弱表达(图3-B,<0.05),显示具有典型的龄期和组织表达特异性。
2.4 不同药剂处理对绿盲蝽若虫发育历期的影响
与对照相比,20E处理后绿盲蝽若虫期的发育历期显著缩短(14.94 d17.04 d,<0.05);相反,20E信号通路中PLC抑制剂U73122处理组若虫期的发育历期显著延长(18.09 d17.04 d,<0.05)。绿盲蝽若虫各龄期发育历期与整个若虫期发育趋势一致,基本为U73122>20E+U73122>乙醇(CK)>20E。显示出20E可加快绿盲蝽若虫的发育(表2)。
2.5 不同药剂处理对绿盲蝽若虫体重及羽化率的影响
20E处理组5龄若虫体重显著低于对照(<0.05),降低了9.81%(4-A);成虫羽化率结果表明(图4-B),与对照相比,20E处理组成虫羽化率最低,其次是U73122处理组,分别下降了15.36%和12.19%。此外,20E与其抑制剂U73122处理后的成虫羽化率差异不显著。
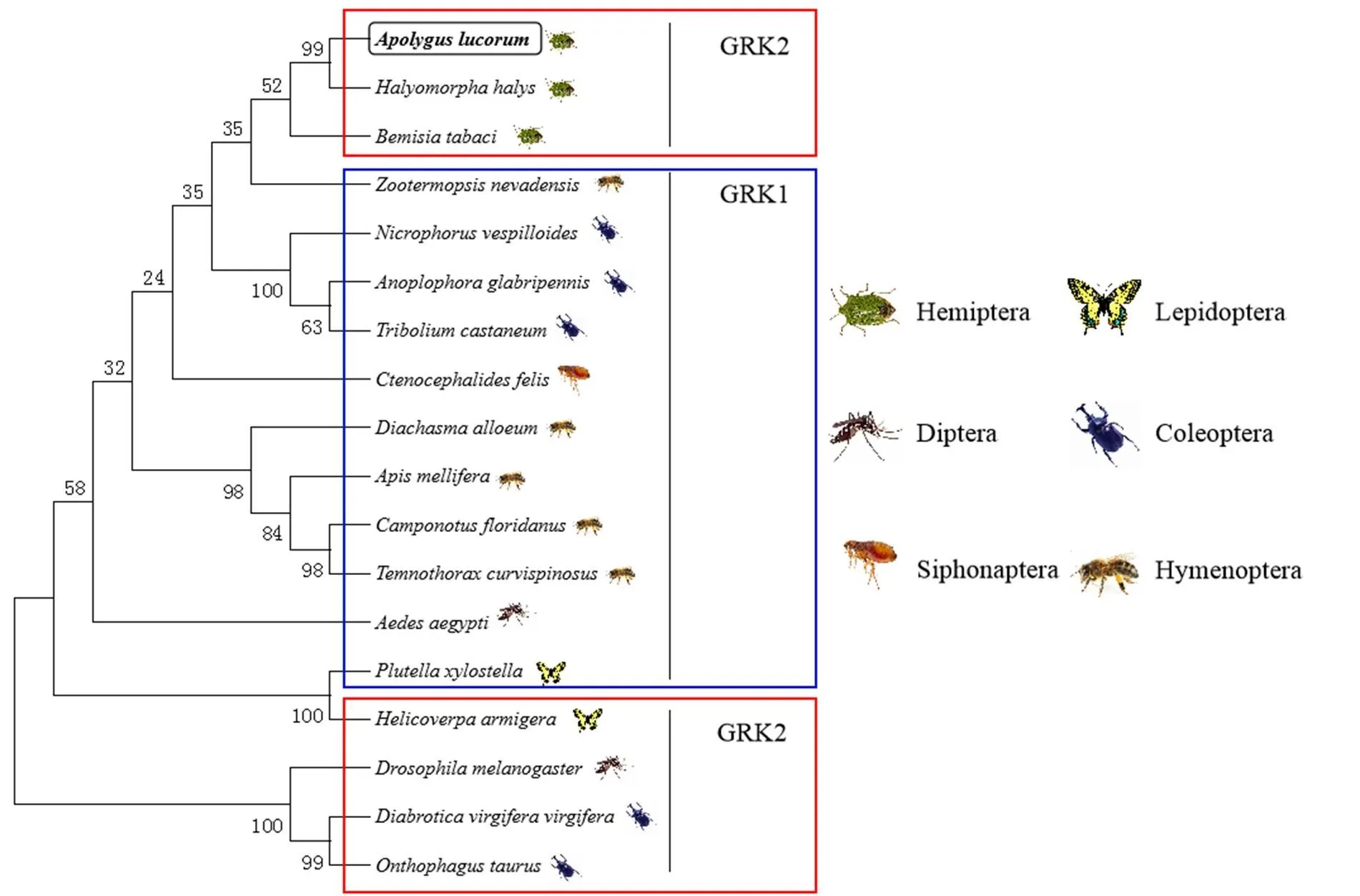
绿盲蝽Apolygus lucorum GRK2(MN514868)、茶翅蝽Halyomorpha halys GRK2(XP_014284300)、烟粉虱Bemisia tabaci GRK2(XP_018907528)、湿木白蚁Zootermopsis nevadensis GRK1(XP_021942271)、大红葬甲Nicrophorus vespilloides GRK1(XP_017785563)、光肩星天牛Anoplophora glabripennis GRK1(XP_023313123)、赤拟谷盗Tribolium castaneum GRK1(XP_015838145)、猫蚤Ctenocephalides felis GRK1(XP_026471851)、胡锋Diachasma alloeum GRK1(XP_015116743)、西方蜜蜂Apis mellifera GRK1(XP_026297912)、佛罗里达弓背蚁Camponotus floridanus GRK1(XP_011267492)、切胸蚁Temnothorax curvispinosus GRK1(XP_024874981)、埃及伊蚊Aedes aegypti GRK1(XP_021706708)、小菜蛾Plutella xylostella GRK1(XP_011558851)、棉铃虫Helicoverpa armigera GRK2(ANZ22924)、黑腹果蝇Drosophila melanogaster GRK2(NP_476867)、玉米根萤叶甲Diabrotica virgifera virgifera GRK2(XP_028135185)、食粪金龟Onthophagus taurus GRK2(XP_022917771)
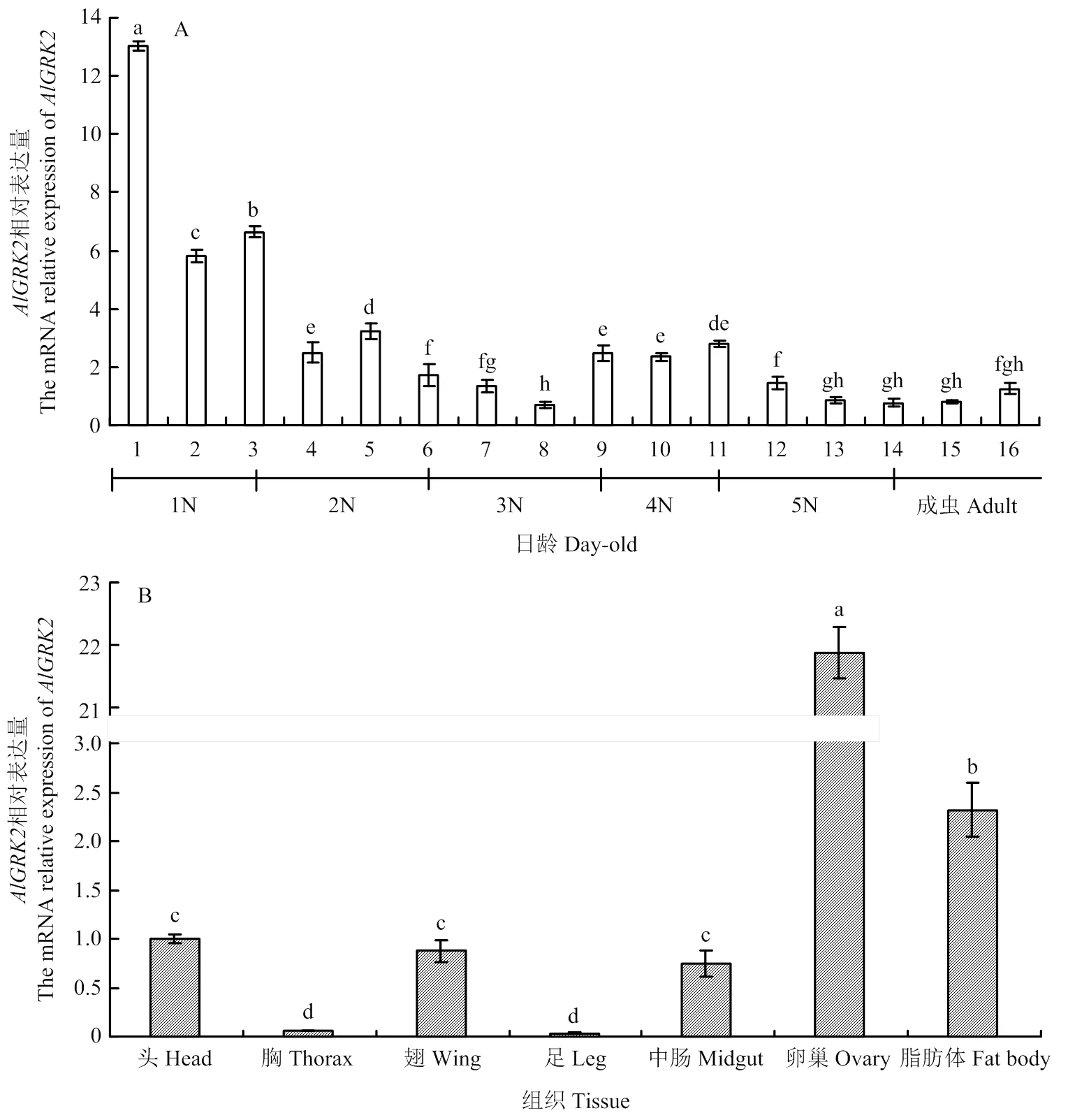
图中数据为平均数±标准误,柱上不同字母表示差异达显著水平(P<0.05)。下同
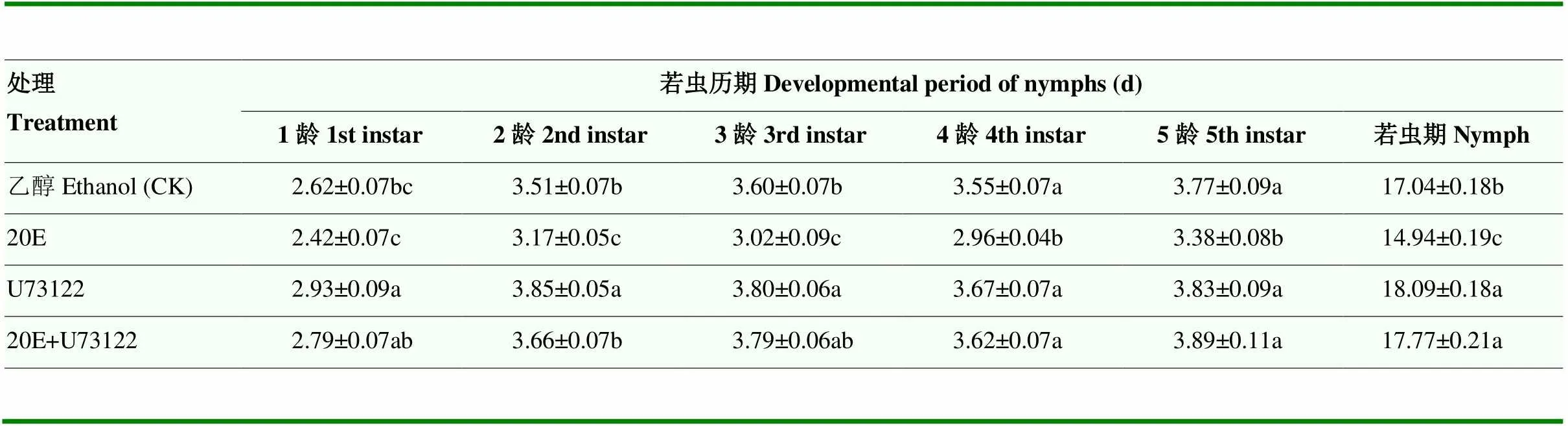
表2 不同药剂处理下绿盲蝽若虫发育历期
表中数据为平均数±标准误,不同小写字母表示差异达显著水平(<0.05)
Data in the table are means±standard error, different lowercases indicate significant difference (<0.05)
2.6 不同药剂处理对AlGRK2表达量的影响
与对照相比,20E处理后的表达显著下降(1、3日龄)(<0.05);而20E抑制剂U73122处理后,供试的1—6日龄绿盲蝽若虫的表达均下调,其中1、3和5日龄下调表达显著(<0.05);总体来看,不同处理的表达量大致趋势为乙醇(CK)>20E>U73122>20E+U73122(图5-A),显示20E有抑制绿盲蝽若虫表达的效应。此外,与对照相比,20E处理后绿盲蝽雌成虫各组织中表达均上调,其中以卵巢和脂肪体中表达量最高(48.57和199.64倍,<0.05);相反,U73122处理后绿盲蝽雌成虫各组织中的表达均下调,其中以卵巢和脂肪体中表达量下调最为显著(<0.05)(图5-B),表明20E有提高绿盲蝽雌成虫组织中表达的效应。
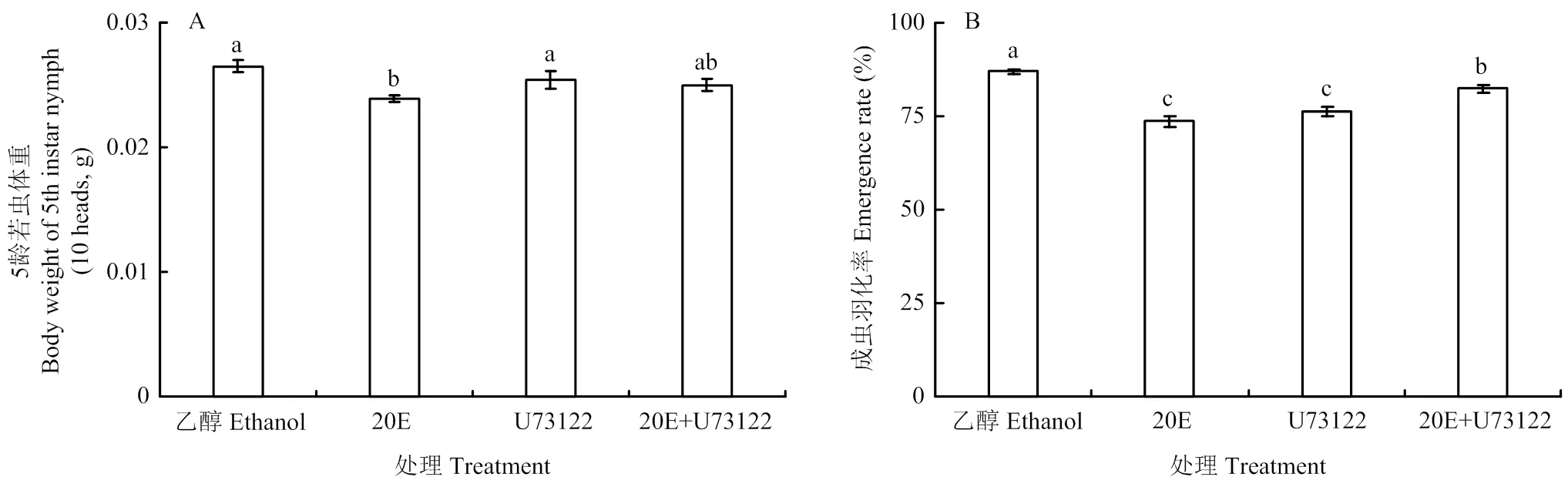
图4 不同处理对绿盲蝽5龄若虫体重(A)和成虫羽化率(B)的影响
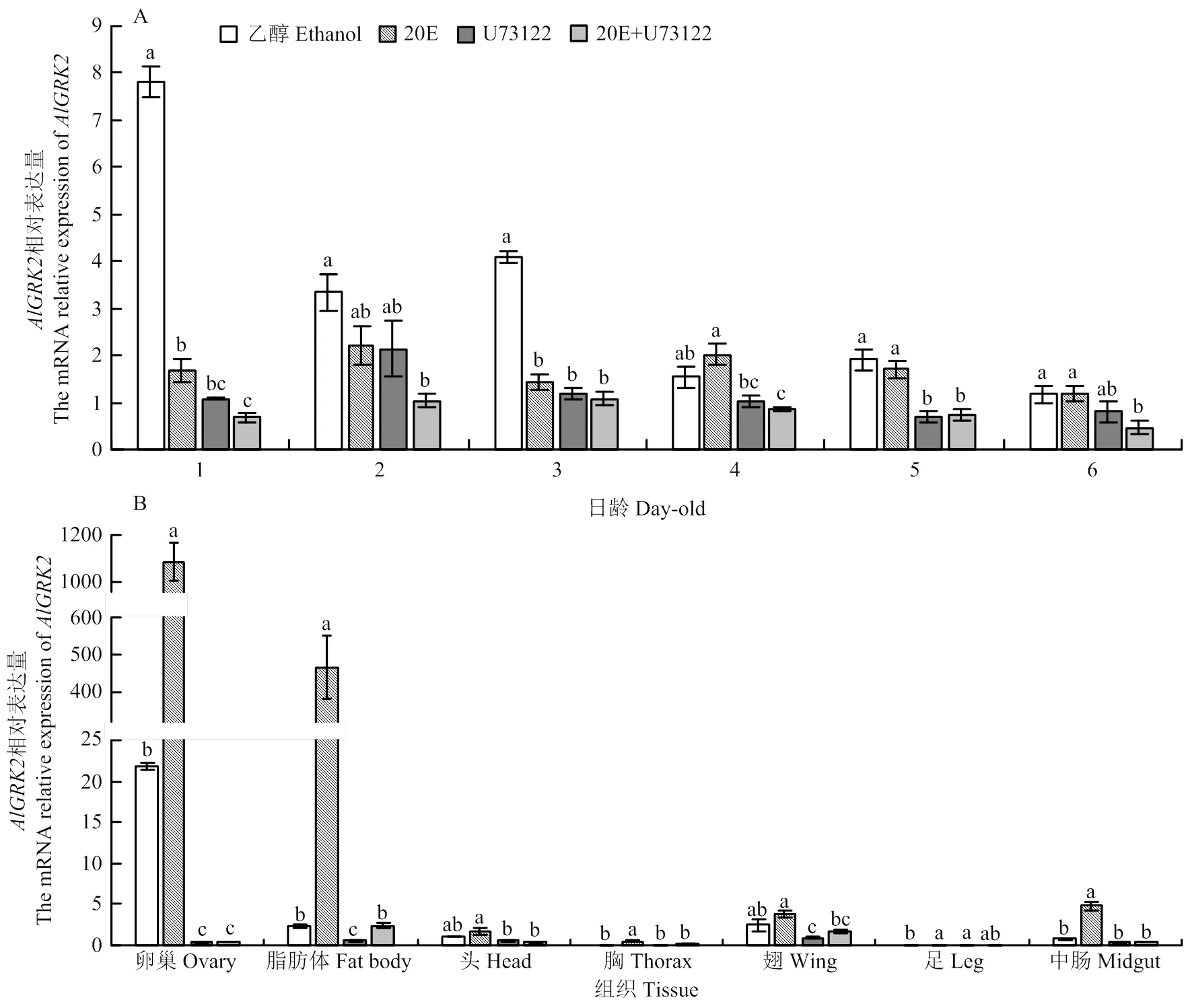
图5 不同处理下绿盲蝽不同日龄(A)和组织(B)中AlGRK2相对表达量
2.7 dsAlGRK2注射对AlGRK2表达量的影响
qRT-PCR检测结果显示,与CK相比,注射ds后的表达水平均显著下降(<0.05,图6)。其中以注射后6 h的下降率最高,与对照组相比分别下降了66.77%(dsCK)和65.62%(dsds);与注射ds对照组相比,注射后12、24和48 h检测发现,绿盲蝽若虫中表达量分别下降了50.31%、38.27%和23.37%。此外,表达量在未注射处理和注射ds处理两个对照组之间无显著差异(>0.05)。表明注射的dsRNA对该基因具有明显的沉默效应。
2.8 dsAlGRK2注射对绿盲蝽生长发育的影响
注射dsRNA后3 d内死亡率统计结果显示(图7-A),与对照组相比,注射ds后,绿盲蝽若虫死亡率显著增加(<0.05),分别增加了73.77%(dsCK)和53.63%(dsds)。若虫死亡率在未注射(CK)和注射ds两个对照组之间无显著差异(>0.05)。注射dsRNA后绿盲蝽成虫羽化率结果表明,注射ds处理组绿盲蝽羽化率最低(图7-B),与注射ds组相比,下降了33.33%。未注射(CK)和注射ds两个对照组之间羽化率无显著差异(>0.05)。
注射ds对5龄若虫体重和发育历期均存在显著影响(<0.05)(图7-C、7-D),其中注射ds后末龄若虫体重显著减少了25.52%(dsds),而发育历期显著延长,推迟了11.42%(dsds)。未注射(CK)和注射ds两个对照组之间均无显著差异(>0.05)。
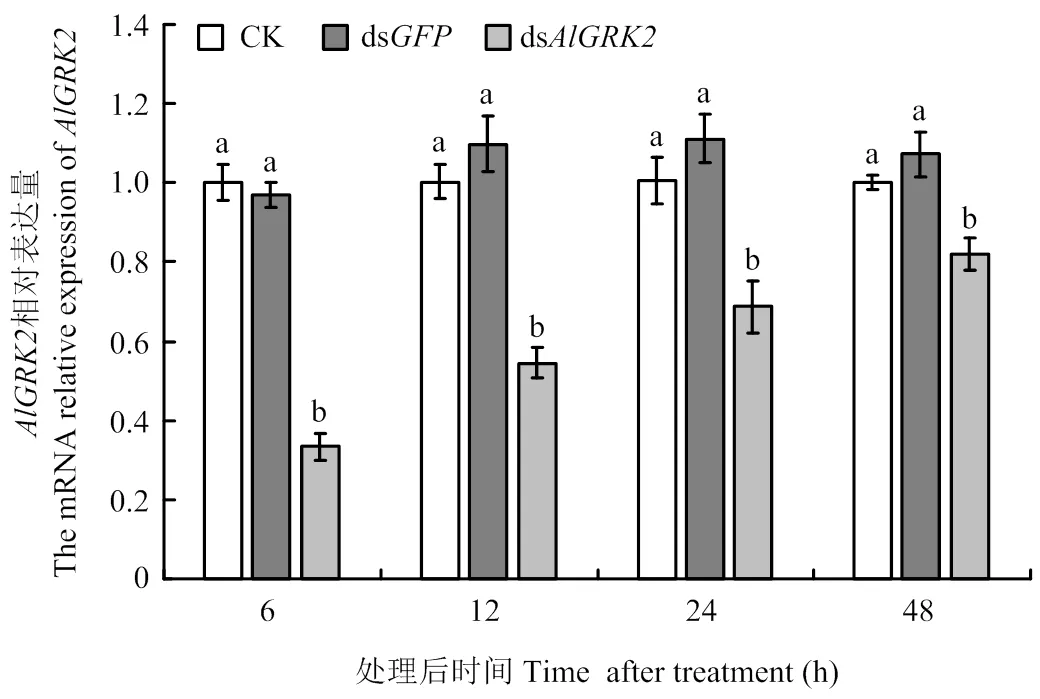
图6 dsRNA处理后绿盲蝽若虫AlGRK2的相对表达量
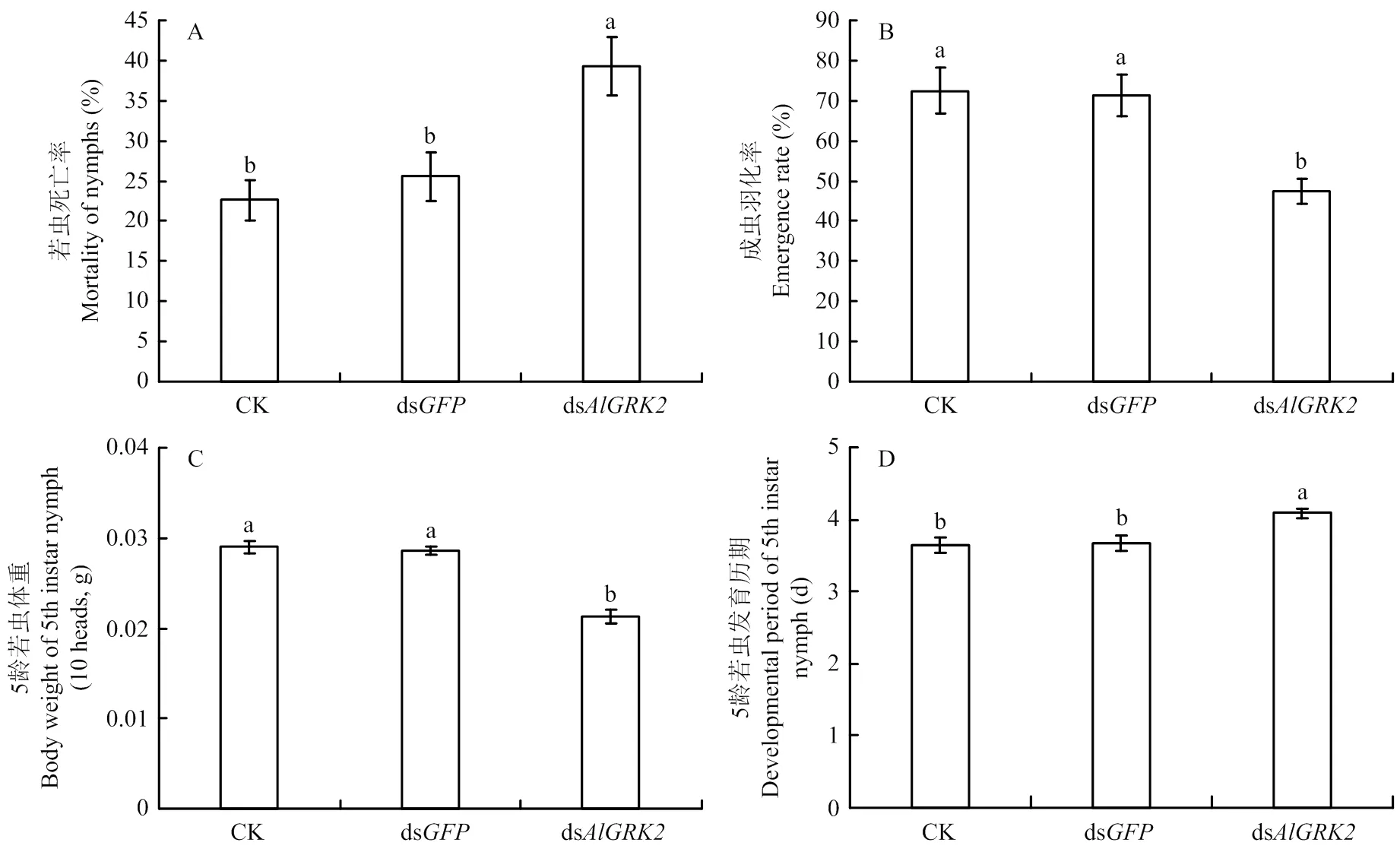
图7 dsRNA处理对绿盲蝽若虫死亡率(A)、成虫羽化率(B)、5龄若虫体重(C)和发育历期(D)的影响
3 讨论
蜕皮激素又称蜕皮酮,在昆虫幼虫或若虫阶段主要由前胸腺分泌,并在P450酶催化下转化为有活性的20-羟基蜕皮酮,进而与保幼激素协同合作以调控昆虫的生长发育与变态。此外,成虫阶段的性腺器官也是主要分泌器官之一,进一步调控昆虫的生殖和胚胎发育[25-28]。蜕皮激素作为昆虫生长发育的关键激素,其滴度含量对昆虫生长发育具有核心的调控作用,一旦昆虫体内激素含量出现异常,可能会对昆虫产生不可逆的损伤。Malausa等[29]对桃蚜()的研究发现,高浓度20E显著降低其若虫数量;Sun等[30]对小菜蛾()的研究发现,外源20E显著降低其幼虫体重、发育历期、繁殖和存活率。本研究结果显示,外源20E显著缩短了绿盲蝽若虫发育历期,且末龄若虫体重和羽化率均显著降低。另外,20E抑制剂U73122处理下可显著延长绿盲蝽若虫发育历期,并有降低羽化率的效应。表明20E可促进绿盲蝽的生长发育。
迄今为止,科学家研究证实蜕皮激素的转导途径存在基因组和非基因组两种途径。其中,20E通过细胞膜上的GPCR调控PLC、Ca2+、PKC等效应因子的表达,进而诱导基因组途径的启动,是目前已明确的非基因组途径[7-8]。GPCR是目前发现的最大跨膜受体家族,能传递多种细胞外信号,而GRK作为GPCR负调控因子,是GPCR信号转导过程中的重要调控开关[31-32]。根据蛋白结构差异,GRK一般包含3个亚族的7个成员。目前该激酶已在哺乳动物体内广泛研究,并发现多与疾病相关[33]。但是,昆虫GRK的功能目前仅在一些模式昆虫中有相关报道。果蝇中报道有3类GRK蛋白,而在一些鳞翅目、半翅目和鞘翅目昆虫中发现1—2类亚型[19-20],且其具体的功能尚未清楚。因此,本文以绿盲蝽为研究对象,首先通过RACE方法,克隆获得了一种绿盲蝽,进一步通过昆虫同源性分析及蛋白结构推导,发现该基因具备亚型典型的PH结构域特征,因此将该基因命名为。
在本研究中,在绿盲蝽前3日龄若虫中表达量显著高于其他日龄,1—16日龄整体表达量呈现波动下降趋势;在绿盲蝽雌成虫卵巢和脂肪体中高表达,而在胸和足中表达量较低。这些结果暗示在绿盲蝽组织和龄期中具有表达特异性,可能存在着不同的功能。具体来说,在绿盲蝽若虫蜕皮前后高表达,笔者推测GRK2是20E信号通路中的重要分子,参与了20E调控绿盲蝽的蜕皮进程;在卵巢和脂肪体中高表达,表明参与了绿盲蝽的卵巢发育及卵子形成。此外,外源20E处理后,1日龄和3日龄的相对表达量显著下调,说明20E对绿盲蝽初孵若虫的表达存在抑制作用。但Zhao等[19]通过对棉铃虫6龄幼虫进行注射处理6 h后,发现20E上调表皮中的表达水平。在本研究中,20E促进了绿盲蝽若虫的发育,降低了的表达、5龄若虫的体重及成虫羽化率;而ds处理后,的表达量、5龄若虫的体重及成虫羽化率均呈现出下降趋势,但绿盲蝽若虫的发育却受到了延缓。实际上,过快发育或发育历期的显著增加,可能是不利于昆虫正常生长发育的两个方面。此外,出现上述结果的原因,还可能与处理方式及处理龄期的差异有关。APPLE等研究发现,一定浓度蜕皮激素处理6 h后,果蝇表皮蛋白相关基因显著上调表达,而当蜕皮激素持续处理果蝇后,表皮蛋白相关基因表达量反而下调[34-35]。上述结果表明,昆虫发育和变态过程中体内蜕皮激素含量是动态的,含量过高会抑制下游基因的表达。此外,在哺乳动物中,抑制蛋白及Mdm2(一种泛素连接酶E3)等介导的磷酸化作用对的降解具有调控作用[36-38]。抑制蛋白和GRK共同作用可导致GPCR发生脱敏反应,Salcedo等[36]研究发现,抑制蛋白可促进泛素连接酶E3与GRK2结合,促进GRK2蛋白的降解。
GRK2是哺乳动物胚胎发育过程中必不可少的蛋白,表达量下调会导致胚胎发育迟缓,而该基因的缺失甚至会引发胚胎死亡[39-40]。Wang等[41]研究发现,GRK2功能丧失不利于秀丽隐杆线虫()产卵。本研究发现绿盲蝽雌虫经过20E处理后,各组织中表达量均有所上升,其中卵巢和脂肪体中显著上调表达,而20E抑制剂U73122处理雌虫后,卵巢、脂肪体和翅中表达量均显著下调,说明20E对绿盲蝽组织中的表达存在诱导作用。昆虫脂肪体和卵巢分别是卵黄及卵子发生场所,以往的研究表明,昆虫卵产生过程中,蜕皮激素能够调控脂肪体中卵黄蛋白的合成、卵壳的发生及卵子的成熟等过程[26,42],而GRK2作为细胞周期调节器在早期胚胎发育过程中担任重要角色[43]。因此笔者推测,在蜕皮激素转导途径中参与调控雌虫生育和繁殖过程,这也是笔者团队后续正在进行的工作。
RNA干扰(RNA interference,RNAi)是由保守的双链RNA(double-stranded RNA,dsRNA)特异性与靶标基因mRNA序列配对,从而高效、特异性降解靶标基因mRNA的表达[44],最终导致靶标基因沉默。该技术作为一种重要的研究手段,在昆虫基因功能的研究中已被广泛应用[45-46]。本研究通过全长序列设计引物合成dsRNA,ds注射后6、12、24和48 h检测表达量均显著低于对照,其中注射后6 h绿盲蝽体内表达量最低。在家蚕中,参与脂动激素受体内吞作用,进而调控家蚕体内脂类和糖类代谢[47]。此外,还可参与棉铃虫20E的内化过程,并且表达减少会导致果蝇躯体分化异常和无法飞行[19-20]。本研究发现,注射ds后绿盲蝽表达显著下降,且若虫死亡率显著增加,而成虫羽化率和末龄若虫体重均显著降低。这与本研究中20E有抑制表达的效应,并对成虫羽化率和末龄若虫体重存在副效应的结果吻合。说明AlGRK2是20E信号调控绿盲蝽生长发育过程中的重要分子,下一步的研究重点为明确介导20E信号转导的内在分子机制。
4 结论
在绿盲蝽体内的表达谱显示出发育阶段和组织特异性;外源20E抑制剂及RNAi处理后,均可抑制的表达,同时还可对绿盲蝽生长发育产生不利影响,表明AlGRK2参与了20E对绿盲蝽生长发育的调控。
[1] 陆宴辉, 吴孔明. 棉花盲椿象及其防治. 北京: 金盾出版社, 2008.
LU Y H, WU K M. The cotton mirids and its control. Beijing: Jindun Publishing House, 2008. (in Chinese)
[2] LU Y H, WU K M, JIANG Y Y, XIAO B, LI P, FENG H Q, WYCKHUYS K A G, GUO Y Y. Mirid bug outbreaks in multiple crops correlated with wide-scale adoption of Bt cotton in China. Science, 2010, 328(5982): 1151-1154.
[3] PAN H S, LIU B, LU Y H, WYCKHUYS K A G. Seasonal alterations in host range and fidelity in the polyphagous mirid bug,(Heteroptera: Miridae). Plos One, 2015, 10(2): e0117153.
[4] COSTANTINO B F B, BRICHE D K, ALEXANDRE K, SHEN K, MERRIAM J R, ANTONIEWSKI C, CALLENDER J L, HENRICH V C, PRESENTE A, ANDRES A J. A novel ecdysone receptor mediates steroid-regulated developmental events during the mid-third instar of. Plos Genetics, 2008, 4(6): e1000102.
[5] CHEN C H, PAN J, DI Y Q, LIU W, HOU L, WANG J X, ZHAO X F. Protein kinase c delta phosphorylates ecdysone receptor B1 to promote gene expression and apoptosis under 20-hydroxyecdysone regulation. Proceedings of the National Academy of Sciences of the United States of America, 2017, 114(34): E7121-E7130.
[6] 谭永安, 肖留斌, 郝德君, 赵静, 孙洋, 柏立新. 绿盲蝽AlEcR-A的单克隆抗体制备及在外源20E诱导下的应答. 中国农业科学, 2017, 50(1): 86-93.
TAN Y A, XIAO L B, HAO D J, ZHAO J, SUN Y, BAI L X. Preparation of monoclonal antibody against AlEcR-A protein and its response induced by exogenous 20-hydroxyecdysone in.Scientia Agricultura Sinica, 2017, 50(1): 86-93. (in Chinese)
[7] LIU W, CAI M J, WANG J X, ZHAO X F. In a nongenomic action, steroid hormone 20-hydroxyecdysone induces phosphorylation of cyclin-dependent kinase 10 to promote gene transcription. Endocrinology, 2014, 155(5): 1738-1750.
[8] 谭永安. 磷脂酶C、E75在绿盲蝽蜕皮激素信号传导中的功能分析[D]. 南京: 南京林业大学, 2019.
TAN Y A. The functional of phospholipase C and E75 in 20E pathway of[D]. Nanjing: Nanjing Forestry University, 2019. (in Chinese)
[9] PENELA P, RIBAS C, MAYOR F. Mechanisms of regulation of the expression and function of G protein-coupled receptor kinases. Cellular Signallin, 2003, 15(11): 973-981.
[10] ZHANG X, KIM K M. Multifactorial regulation of G protein-coupled receptor endocytosis. Biomolecules and Therapeutics, 2017, 25(1): 26-43.
[11] GUREVICH E V, TEAMER J J G, MUSHEGIAN A, GUREVICH V V. G protein-coupled receptor kinases: more than just kinases and not only for GPCRs. Pharmacology and Therapeutics, 2012, 133(1): 40-69.
[12] KOMOLOV K E, BENOVIC J L. G protein-coupled receptor kinases: past, present and future. Cellular Signalling, 2018, 41: 17-24.
[13] INGLESE J, KOCH WJ, CARON MG, LEFKOWITZ R J. Isoprenylation in regulation of signal transduction by G-protein-coupled receptor kinases. Nature, 1992, 359(6391):147-150.
[14] BENOVIC J L, MAYOR F, SOMERS R L, CARON M G, LEFKOWITZ R J. Light-dependent phosphorylation of rhodopsin by-adrenergic receptor kinase. Nature, 1986, 321(6073): 869-872.
[15] BOEKHOFF I, INGLESE J, SCHLEICHER S, KOCH W J, LEFKOWITZ R J, BREER H. Olfactory desensitization requires membrane targeting of receptor kinase mediated by-subunits of heterotrimeric G proteins. The Journal of Biological Chemistry, 1994, 269(1): 37-40.
[16] OPPERMANN M, DIVERSE-PIERLUISSI M, DRAZNER M H, DYER S L, FREEDMAN N J, PEPPEL K C, Lefkowitz R J. Monoclonal antibodies reveal receptor specificity among G-protein- coupled receptor kinases. Proceedings of the National Academy of Sciences of the United States of America, 1996, 93(15): 7649-7654.
[17] STOFFEL R H, RANDALL R R, PREMONT R T, LEFKOWITZ R J, INGLESE J. Palmitoylation of G protein-coupled receptor kinase, GRK6. lipid modification diversity in the GRK family. The Journal of Biological Chemistry, 1994, 269(45): 27791-27794.
[18] PREMONT R T, MACRAE A D, STOFFEL R H, CHUNG N J, PITCHER J A, AMBROSE C, INGLESE J, MACDONALD M E, LEFKOWITZ R J. Characterization of the G protein-coupled receptor kinase GRK4. Identification of four splice variants. The Journal of Biological Chemistry, 1996, 271(11): 6403-6410.
[19] ZHAO W L, WANG D, LIU C Y, ZHAO X F. G-protein-coupled receptor kinase 2 terminates G-protein-coupled receptor function in steroid hormone 20-hydroxyecdysone signaling. Scientific Reports, 2016, 6: 29205.
[20] Garcia-Guerra L, Vila-Bedmar R, Carrasco-Rando M, Cruces-Sande M, Martín M, Ruiz-Gómez A, Ruiz-Gómez M, Lorenzo M, Fernández-Veledo S, Mayor F, Murga C, Nieto-Vázquez I. Skeletal muscle myogenesis is regulated by G protein-coupled receptor kinase 2.Journal of molecular cell biology, 2014, 6(4): 299-311.
[21] KUBO I, KLOCKE J A, ASANO S. Insect ecdysis inhibitors from the East African medicinal plant(Labiatae). Agricultural and biological chemistry, 1981, 45(8): 1925-1927.
[22] BLACKFORD M J P, DINAN L. The effects of ingested 20- hydroxyecdysone on the larvae of,,(Lepidoptera: Nymphalidae) and(Lepidoptera: Arctiidae). Journal of Insect Physiology, 1997, 43(4): 315-327.
[23] KLEIN R R, BOURDON D M, COSTALES C L, WAGNER C D, WHITE W L, WILLIAMS J D, HICKS S N, SONDEK J, THAKKER D R. Direct activation of human phospholipase C by its well known inhibitor U73122. The Journal of Biological Chemistry, 2011, 286(14): 12407-12416.
[24] LIVAK K J, SCHMITTGEN T D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCTmethod. Methods, 2001, 25: 402-408.
[25] KARLSON P. On the hormonal control of insect metamorphosis. A historical review. The International Journal of Developmental Biology, 1996, 40(1): 93-96.
[26] LANOT R, THIEBOLD J, COSTET-CORIO M F, BENVENISTE P, HOFFMANN J A. Further experimental evidence for the involvement of ecdysone in the control of meiotic reinitiation in oocytes of(Insecta, Orthoptera). Developmental Biology, 1988, 126(1): 212-214.
[27] LOEB M J, DE LOOF A, GELMAN D B, HAKIM R S, JAFFE H, KOCHANSKY J P, MEOLA S M, SCHOOFS L, STEEL C, VAFOPOULOU X, WAGNER R M, WOODS C W. Testis ecdysiotropin, an insect gonadotropin that induces synthesis of ecdysteroid. Archives of Insect Biochemistry and Physiology, 2010, 47(4): 181-188.
[28] DAIMON T, UCHIBORI M, NAKAO H, SEZUTSU H, SHINODA T. Knockout silkworms reveal a dispensable role for juvenile hormones in holometabolous life cycle. Proceedings of the National Academy of Sciences of the United States of America, 2015, 112(31): E4226-E4235.
[29] MALAUSA T, SALLES M, MARQUET V, GUILLEMAUD T, ALLA S, MARION-POLL F, LAPCHIN L. Within-species variability of the response to 20-hydroxyecdysone in peach-potato aphid (Sulzer). Journal of Insect Physiology, 2006, 52(5): 480-486.
[30] SUN L J, LIU Y J, SHEN C P. The effects of exogenous 20-hydroxyecdysone on the feeding, development, and reproduction of(Lepidoptera: Plutellidae). Florida Entomologist, 2015, 98(2): 606-612.
[31] MARINISSEN M J, GUTKIND J S. G-protein-coupled receptors and signaling networks: emerging paradigms. Trends in Pharmacological Sciences, 2001, 22(7): 368-376.
[32] BELMONTE S L, BLAXALL B C. G protein coupled receptor kinases as therapeutic targets in cardiovascular disease. Circulation Research, 2011, 109(3): 309-319.
[33] KANG J H, TOITA R, KAWANO T, MURATA M, ASAI D. Design of substrates and inhibitors of G protein-coupled receptor kinase 2 (GRK2) based on its phosphorylation reaction. Amino Acids, 2020, 52(6): 863-870.
[34] APPLE R T, FRISTROM J W. 20-Hydroxyecdysone is required for, and negatively regulates, transcription ofpupal cuticle protein genes. Developmental Biology, 1991, 146(2): 569-582.
[35] DOCTOR J, FRISTRIM D, FRISTRIM J W. The pupal cuticle of: biphasic synthesis of pupal cuticle proteinsandin response to 20-hydroxyecdysone. The Journal of Cell Biology, 1985, 101(1): 189-200.
[36] SALCEDO A, MAYOR F, PENELA P. Mdm2 is involved in the ubiquitination and degradation of G-protein-coupled receptor kinase 2. The EMBO Journal, 2006, 25(20): 4752-4762.
[37] HUANG J A, NALLI A D, MSHAVADI S, KUMAR D P, MURTHY K S. Inhibition of Giactivity by Gis mediated by PI 3-kinase-- and cSrc- dependent tyrosine phosphorylation of Giand recruitment of RGS12. American Journal of Physiology Gastrointestinal and Liver Physiology, 2014, 306(9): G802-G810.
[38] WALDSCHMIDT H V, HOMAN K T, CRUZ-RODRIGUEZ O, CATO M C, WANINGER-SARONI J, LARIMORE K M, CANNAVO A, SONG J, CHEUNG J Y, KIRCHHOFF P D, KOCH W J, TESMER J J G, LARSEN S D. Structure-based design, synthesis and biological evaluation of highly selective and potent G protein-coupled receptor kinase 2 inhibitors. Journal of Medicinal Chemistry, 2016, 59(8): 3793-3807.
[39] JABER M, KOCH W J, ROCKMAN H, SMITH B, BOND R A, SULIK K K, ROSS J, LEFKOWITZ R J, CARON M G, GIROS B. Essential role of-adrenergic receptor kinase 1 in cardiac development and function. Proceedings of the National Academy of Sciences of the United States of America, 1996, 93(23): 12974-12979.
[40] PHILIPP M, FRALISH G B, MEIONI A R, CHEN W, MACINNES A W, BARAK L S, CARON M G. Smoothened signaling in vertebrates is facilitated by a G protein-coupled receptor kinase. Molecular Biology of the Cell, 2008, 19(12): 5478-5489.
[41] WANG J J, LUO J S, ARYAL D K, WETSEL W C, NASS R, BENOVIC J L. G protein-coupled receptor kinase-2 (GRK-2) regulates serotonin metabolism through the monoamine oxidase AMX-2 in. The Journal of Biological Chemistry, 2017, 292(14): 5943-5956.
[42] ISAAC R E, REES H H. Isolation and identification of ecdysteroid phosphates and acetylecdysteroid phosphates from developing eggs of the locust,. The Biochemical Journal, 1984, 221(2): 459-464.
[43] JIANG X, YANG P, MA L. Kinase activity-independent regulation of cyclin pathway by GRK2 is essential for zebrafish early development. Proceedings of the National Academy of sciences of the United States of America, 2009, 106(25): 10183-10188.
[44] 杨中侠, 文礼章, 吴青君, 王少丽, 徐宝云, 张友军. RNAi技术在昆虫功能基因研究中的应用进展. 昆虫学报, 2008, 51(10): 1077-1082.
YANG Z X, WEN L Z, WU Q J, WANG S L, XU B Y, ZHANG Y J. Application of RNA interference in studying gene functions in insects. Acta Entomologica Sinica, 2008, 51(10): 1077-1082. (in Chinese)
[45] MCFARLANE M, LAURETI M, LEVEE T, TERRY S, KOHL A, PONDEVILLE E. Improved transient silencing of gene expression in the mosquito female. Insect Molecular Biology, 2021, 30(3): 355-365.
[46] ZHU J H, LIU X Q, ZHU K M, ZHOU H Y, LI L, LI Z X, QIN W W, HE Y P. Knockdown of TRPV genes affects the locomotion and feeding behavior of(Hemiptera: Delphacidae). Journal of Insect Science, 2020, 20(1): 9.
[47] 黄海山. 家蚕脂动激素受体信号转导机制研究[D]. 杭州: 浙江大学, 2010.
HUANG H S. The mechanism of signal transduction of adipokinetic hormone receptor in[D]. Hangzhou: Zhejiang University, 2010. (in Chinese)
Expression profile of G protein-coupled receptor kinase 2 gene () and its function in the development of
TAN YongAn, Jiang YiPing, ZHAO Jing, XIAO LiuBin
Institute of Plant Protection, Jiangsu Academy of Agricultural Sciences, Nanjing 210014
【Objective】The objective of this study is to clone the full-length cDNA of the G protein-coupled receptor kinase 2 () in, clarify its expression profiles and the effect of exogenous ecdysterone hormone 20E on the expressionof, analyze the role of, and to provide a preliminary research basis for further study of its function in the ecdysone signaling transduction pathway. 【Method】Thewas cloned and obtained by RACE method. Using the qRT-PCR method, the relative expression levels ofwere determined. Finally, the response of themRNA expression inafter exogenous 20E induction and RNAi treatment and their effects on the main parameters (development progress, nymphs weight and adult emergence rate) in the growth and development of thewere analyzed.【Result】The full length ofcDNA is 2 715 bp, and ORF is 2 106 bp, which encodes 701 amino acids. ExPASy predicts that the protein molecular weight is 80.2 kD and the theoretical isoelectric point is 6.56. Protein structure analysis shows that AlGRK2 contains 4 domains: G protein signalling domain (RGS, 54-175 aa), serine/threonine protein kinases domain (S-TKc, 191-454 aa), extension to Ser/Thr-type protein kinases (S-TK-X, 455-534 aa) and pleckstrin homology domain (PH, 558-655 aa), and the PH domain is a typical protein feature in the GRK2 subtype. Phylogenetic analysis showed that GRK2 inhad the closest genetic relationship withGRK2. qRT-PCR results showed thatwas expressed in 1-day-old to 16-day-old nymphs of, and the mRNA expression showed a fluctuating downward pattern. The expression level ofwas higher in the initial instar,but decreased significantly in the last instar of. Thewas highly expressed in the ovary and fat body of female adults and less expressed in the thorax and leg. After exogenous 20E treatment, the expression ofwas significantly down-regulated in nymphs at the 1-day-old and 3-day-old. The relative expression oftreated with U73122 (PLC inhibitor) was down-regulated. Compared with ethanol control, the developmental duration of nymph, the body weight of the last instar nymph and the emergence rate of adult inwere significantly decreased after 20E treatment. On the contrary, the development duration of nymph stage in U73122 treatment group was significantly prolonged. In addition, compared with the dsgroup,expression level ofwas significantly decreased after injection of ds, and nymphs mortality and developmental stages were significantly increased, while adult emergence rate and weight of the 5th instar nymph were significantly decreased. 【Conclusion】The expression profile ofshowed the specificity of the developmental stage and the tissue. Exogenous 20E inhibitor and RNAi treatment can inhibit the expression of, and have adverse effects on the growth and development of, such as delaying the development progress of, reducing the weight of 5th instar nymph and adult emergence rate.
; G protein-coupled receptor kinase 2 (GRK2); ecdysterone hormone; gene cloning; expression profile; RNAi
2021-04-16;
2021-06-10
国家重点研发计划(2017YFD0201900)、国家现代农业产业技术体系建设专项资金(CARS-15-20)、国家自然科学基金(31301668)、转基因棉花环境安全性评价技术(2016ZX08011)、江苏省农业科学院院基金(611613)
谭永安,E-mail:kellytan001@163.com。通信作者肖留斌,E-mail:xlb@jaas.ac.cn
(责任编辑 岳梅)
猜你喜欢
资源导刊(信息化测绘)(2022年4期)2022-11-24
农业工程学报(2022年10期)2022-08-22
湖北农业科学(2022年11期)2022-07-18
天津农业科学(2022年5期)2022-05-31
安徽林业科技(2021年2期)2021-06-15
南方农业学报(2021年11期)2021-03-05
实用肿瘤学杂志(2020年4期)2020-12-08
江苏农业科学(2017年12期)2017-09-16
小溪流(画刊)(2017年6期)2017-07-10
医学综述(2011年12期)2011-12-09
