
月季基因组SPL转录因子鉴定及表达特征分析
2021-11-29 07:07张宇鑫李惠张梦宋翔余游余马陈华
湖北农业科学 2021年21期
张宇鑫,李惠,张梦,宋翔,余游,余马,陈华
(1.西南科技大学生命科学与工程学院,四川 绵阳 621010;2.绵阳市农畜产品质量安全检验检测中心,四川 绵阳 621000)
月季(Rosa chinensis)是世界上最受欢迎的观赏花之一,具有重要观赏价值和经济价值。花瓣的形状及颜色决定了月季的五彩斑斓,因而花瓣的发育对月季的美观起着重要作用。Squamosa promoter binding protein like(SPL)基因家族作为植物特有的转录因子,其家族成员能够与SQUAMOSA启动子结合,被称为SQUA启动子结合蛋白[1]。SPL基因编码一个高度保守的SBP结构域。该结构域约含76个氨基酸残基,包括Cys-Cys-His-Cys和Cys-Cys-Cys-His两个串联锌指,并在C端具有核定位信号[2]。研究表明,SPL调控植物从营养生长到生殖生长的转化,调节叶片形态发生,参与环境信号的响应对植物生长发育发挥着重要作用[3]。此外,SPL基因参与调节花器官的形成及其后期发育[4]。
SPL基因最初从金鱼草的cDNA文库中分离出来[5],随着植物基因组数据的不断发表,许多物种相继进行了SPL基因的全基因组鉴定与分析。拟南芥的基因组共鉴定出16个SPL基因[6],随后SPL家族在番茄、苦荞、白桦树、绿藻、苔藓、玉米、葡萄、苹果、甜瓜、丹参、蓖麻、棉花、梅花、矮牵牛花、柑橘、小麦、菊花、辣椒等[7-24]中相继被分离鉴定出来,但目前月季的SPL基因尚未被鉴定。
本研究通过生物信息学的方法对月季RcSPL基因家族进行了全面分析,包括基因结构、染色体位置和共线性分析等,旨在探究RcSPL基因在月季花瓣发育中所行使的功能,为调控月季花瓣发育提供分子基础。
1 材料与方法
1.1 材料
从已公布的转录组数据中挖掘月季转录组数据,月季转录组材料为花蕾期、开花期、凋谢期的近轴与远轴端月季花瓣。
1.2 方法
1.2.1 月季SPL基因序列的鉴定 月季的基因组序列和注释文件下载于月季Rosa chinensis‘Old Blush’基因组网站(https://lipm-browsers.toulouse.inra.fr//pub/RchiOBHm-V2/)[25]。拟 南 芥 的16条SPL基 因下载于拟南芥数据库(https://www.arabidopsis.org)。SPL基因家族的SBP-box结构域(PF03110)下载于Pfam(http://pfam.xfam.org)数据库。16条拟南芥的SPL基因的蛋白序列被作为query序列进行比对,利用BLASTP(E<10-5)在月季蛋白质数据库进行比对搜索,并除去重复序列。采用HMMER 3.0软件的隐马尔可夫模型(HMM)对SBP结构域[26]再次进行比对搜索,将2次搜索得到的序列取交集,最终确认为月季的RcSPL家族基因数目。
1.2.2 月季SPL基因家族分类及蛋白结构域、顺式作用元件分析 利用ClustalX 1.83将鉴定到的月季RcSPL家族蛋白序列和拟南芥SPL家族蛋白序列进行多重序列比对[27],随后利用MEGA5采用邻接法(neighbor-joining,NJ)构建系统进化树[28],Bootstrap method设置为1 000。利用在线软件GSDS 2.0(http://gsds.cbi.pku.edu.cn)绘制基因结构图[29]。利用 在 线 软 件MEME 5.0.2(http://meme-suite.org)[30]对SPL蛋白家族的保守基序进行预测,预测数目设置为10,其他参数为默认值。利用plantcare(http://bioinformatics.psb.ugent.be)网站预测月季RcSPL基因上游2 000 bp的顺势作用元件。
1.2.3 染色体定位以及共线性分析 使用MapChart 2.2软件绘制月季RcSPL家族在染色体每条染色体的位置[31]。利用MCscanX软件分析月季RcSPL基因家族在基因组中的共线性以及复制事件[32]。
1.2.4 月季SPL基因的表达分析 根据已发表NCBI的SRA数据库中月季转录组数据(https://www.ncbi.nlm.nih.gov/bioproject)提取出RcSPL基因的相关表达量,利用pheatmap软件包绘制相关热图[33]。
2 结果与分析
2.1 月季SPL基因家族成员鉴定、特征分析、进化树分析
通过blast和hmmer在月季的基因组中共检索出17个RcSPL基因,分别命名为RcSPL1至RcSPL17(表1)。系统发育树将拟南芥与月季的RcSPL基因分成8个亚组(图1),RcSPL6为Ⅰ组,RcSPL7、RcSPL8、RcSPL17为Ⅱ组,RcSPL1为Ⅲ组,RcSPL11、RcSPL14、RcSPL15为Ⅳ组,RcSPL2、RcSPL10为Ⅴ组,RcSPL3、RcSPL12、RcSPL13为Ⅵ组,RcSPL4、RcSPL9、RcSPL16为Ⅶ组,RcSPL5为Ⅷ组,Ⅳ组与Ⅶ组处于同一进化分支,Ⅲ组、Ⅷ组、Ⅴ组处于同一分支且亲缘关系更近。RcSPL基因蛋白序列长度为156~1 075 aa,分子量为17.7~118.62 kDa,等电点除RcSPL7均超过7,显弱碱性。
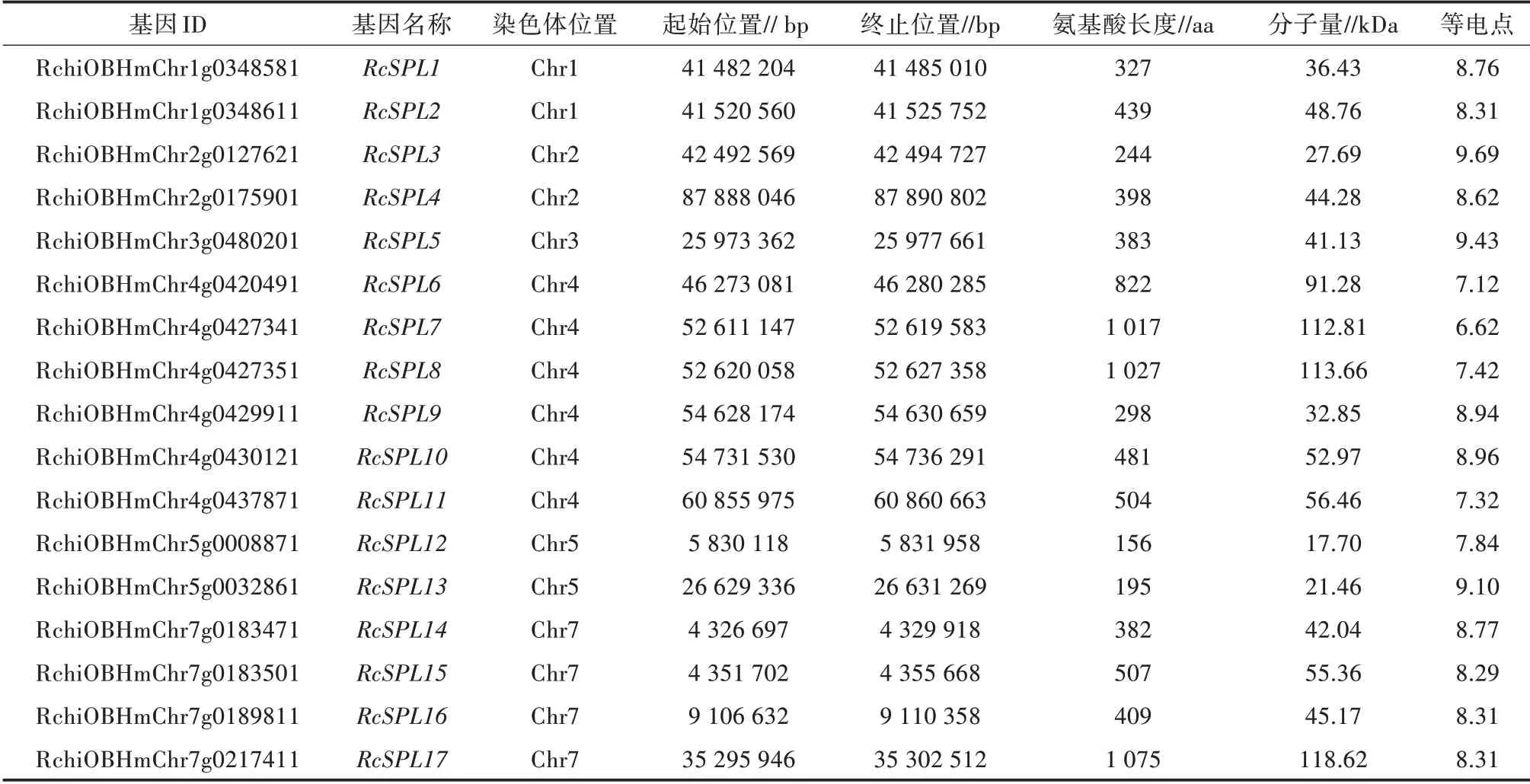
表1 月季SPL基因的起始位置、终止位置、相对分子质量、蛋白序列长度和等电点
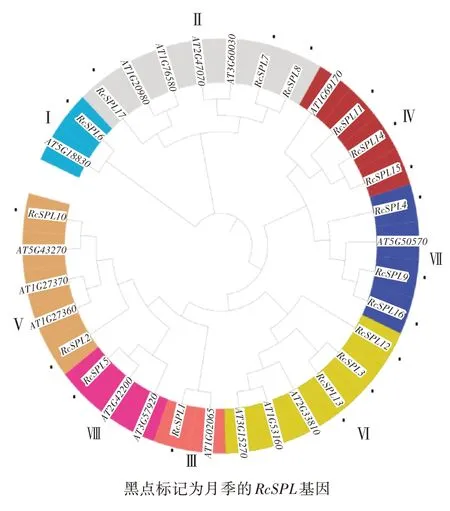
图1 拟南芥和月季的SPL基因的系统发育树
2.2 月季SPL基因保守结构域、motif、基因结构、顺势作用元件分析
多序列比对分析结果表明,RcSPL基因中SBP结构域非常保守(图2),其包含两个锌指结构,Zn-1、Zn-2和一个高度保守的核定位信号。比较RcSPL基因外显子和内含子数量和位置(图3),RcSPL基因含有2~10个外显子,各亚组间成员所含外显子差异较大,Ⅱ亚组中含有最多的外显子,其中RcSPL7包含10个外显子,Ⅵ亚组成员所含外显子最少,均为2~3个;各亚组内成员间外显子数目相似。
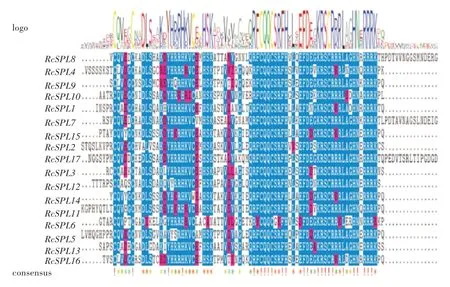
图2 月季SPL蛋白序列关键部位的多序列比对结果

图3 月季SPL基因的基因结构
保守基序分析预测到10个保守基序(图4),将这10个保守基序按顺序命名为Motif 1~Motif 10,分析发现所有蛋白序列均含有Motif 1。Ⅱ亚组成员具有最多的保守基序,并且所含种类也最多,Ⅲ、Ⅴ、Ⅵ、Ⅶ亚组成员均只含有Motif 1。

图4 月季SPL基因的进化关系和motif的预测
预测月季SPL基因上游2 000 bp的顺势作用元件(图5),定向分析了MYB与MYC的作用位点,发现其上游2 000 bp中含有2~13个MYB与MYC作用位点,除RcSPL2、RcSPL5均含有5个及以上的作用位点。
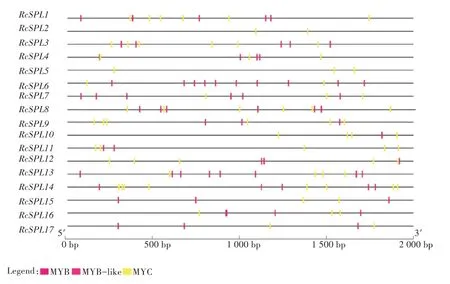
图5 月季SPL基因部分顺式作用元件分布情况
2.3 月季SPL基因的共线性分析、复制分析、染色体定位分析
染色体定位分析表明,RcSPL基因在除6号染色体外的其余6条染色体上均有分布(图6)。RcSPL的复制事件分析结果表明,该家族在月季中有两对串联复制基因,分别为1号染色体的RcSPL1和Rc-SPL2,2号染 色体的RcSPL7和RcSPL8。RcSPL1和RcSPL2分别为Ⅲ亚组和Ⅴ亚组,RcSPL7和RcSPL8均为Ⅱ亚组。共线性分析发现,月季中RcSPL基因有四个共线性块(图7)。

图6 月季SPL基因在染色体上的分布情况

图7 月季SPL基因的共线性分析
2.4 月季SPL基因家族表达分析
通过NCBI转录组数据分析发现月季花蕾期(FB)、开花期(OF)、凋花期(SF)花瓣的近轴(AD)与远轴(AB)RcSPL基因表达相关,大多数月季SPL基因的表达模式具有相似性,RcSPL3与RcSPL5显著正相关为0.9,且与其他基因具有相反的表达模式(图8)。基因表达分析表明月季的RcSPL基因在开花后期表达相对较高,RcSPL3、RcSPL5、RcSPL16在花蕾期表达相对较高,RcSPL1、RcSPL13、RcSPL9、RcSPL12在花凋谢时期表达相对较高,RcSPL2、Rc-SPL14、RcSPL4、RcSPL15在开花期和花蕾期表达相对较 高,RcSPL10、RcSPL11、RcSPL17、RcSPL6、Rc-SPL7、RcSPL8在花蕾期表达相对较高(图9)。
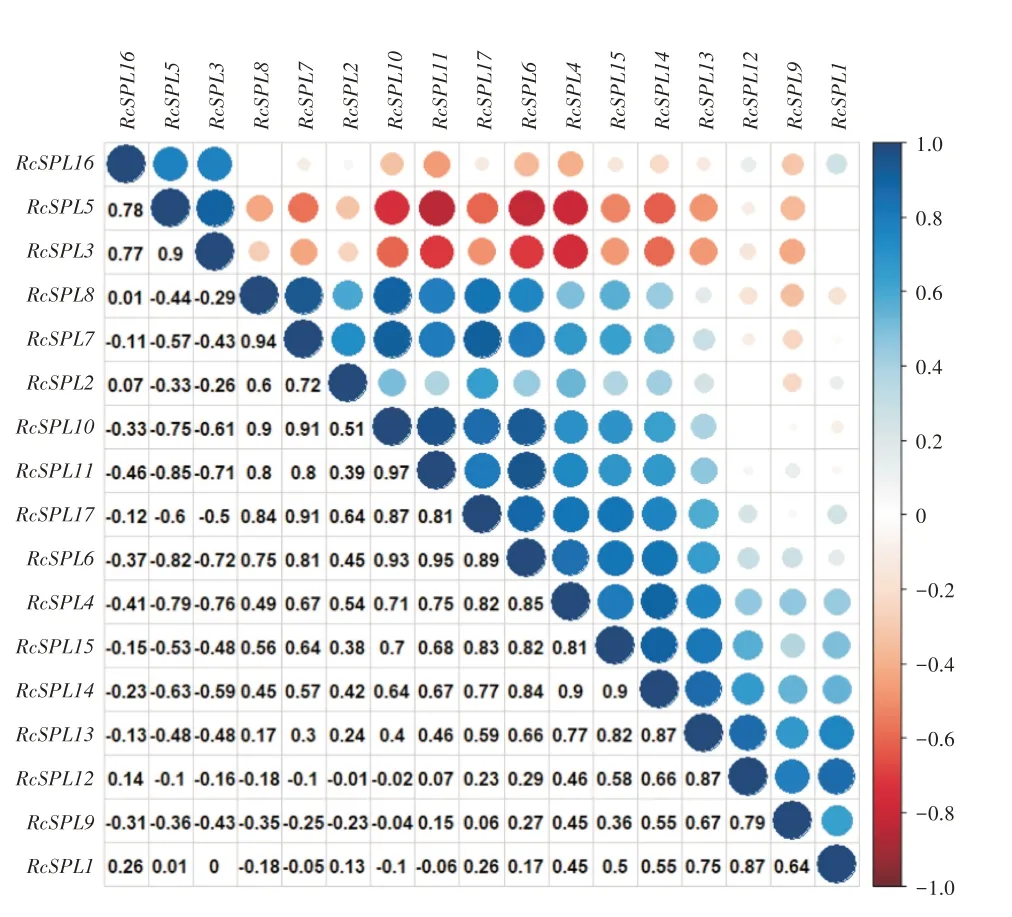
图8 月季SPL基因表达模式的相关性

图9 月季花瓣近轴与远轴RcSPL基因花蕾期、开花期、凋谢期的表达量
3 讨论与结论
SPL基因在植株的生长发育过程行使着重要的功能,研究在月季的基因组中共发掘出17个RcSPL成员,系统发育树分析将其分成8个亚组,每个亚组具有较为相似的基因结构且外显子数目基本相同,这与前人研究结果相符[34,35]。通过RcSPL蛋白序列的保守基序分析发现,每条RcSPL均具有Motif 1。此外,Ⅱ亚组具有最多的保守基序,最多的外显子,具有较高水平的表达,推测其在月季花开过程行使重要的功能。
RcSPL基因上游2 000 bp的顺式作用元件分析发现,该段区域具有2~13个MYB与MYC转录因子的作用位点,MYB是一种调控花色的转录因子,MYB与MYC在花青素的合成中起着重要的作用。有研究表明SPL基因与花青素的含量和花香有关[36],因此推测SPL基因很可能与MYB和MYC相互作用调控月季花青素的含量,共同调控花的生长发育。
分析基因表达模式的相关性发现,大多数月季RcSPL基因的表达模式具有相似性,RcSPL3、RcSPL5的表达模式与其他基因相反,推测这两个基因可能发挥与其他RcSPL基因相反的功能。转录因子通常在控制组织特异性基因的表达中起着关键性作用[37]。根据RcSPL家族表达量的分析,发现RcSPL成员间表达差异明显。研究表明,拟南芥SPL8能够影响花粉的发育[38],GbSBP6基因在银杏胚珠中显著表达且与拟南芥SPL8位于同一亚组,可能参与了银杏的花发育[39],RcSPL1也位于该亚组,推测RcSPL1可能参与月季花粉形成的调控。前人研究表明拟南芥SPL3、SPL4、SPL5编码的转录因子能够调控开花时间和成花转变,该亚组的多数SPL成员与植物生殖器官的生长发育有关,野生葡萄的VvSBP11参与了 成 花 转 变 过 程[40]。RcSPL3与 拟 南 芥 的SPL3、SPL4、SPL5处于同一亚组且在花蕾期有相对的高表达,因此推测RcSPL3可能参与月季开花时间的调控。
猜你喜欢
实用心脑肺血管病杂志(2023年12期)2023-12-14
电子科技大学学报(2022年5期)2022-10-29
园林科技(2021年3期)2022-01-19
中国生殖健康(2020年4期)2021-01-18
心电与循环(2020年1期)2020-02-27
中国生殖健康(2018年4期)2018-11-06
生命科学研究(2018年1期)2018-05-29
上海农业学报(2017年3期)2017-04-10
山东农业工程学院学报(2016年6期)2016-12-01
中国全科医学(2015年26期)2015-02-23
