
人参愈伤诱导及其次生代谢产物的影响因素研究进展
2021-11-29 07:06强宝宝韦坤华梁莹韦桂丽赵立春缪剑华
湖北农业科学 2021年21期
强宝宝,韦坤华,梁莹,韦桂丽,赵立春,缪剑华
(1.广西药用植物园,a.广西药用资源保护与遗传改良重点实验室;b.广西壮族自治区中药资源智慧创制工程研究中心,南宁 530023;2.广西中医药大学,南宁 530200)
人参(Panax ginsengC.A.Mey)是五加科(Araliaceae)人参属(Panax L.)植物,是中国传统名贵中药,被称为“百药之王”,主要分布于东三省、河北省、陕西省等地[1,2],具有大补元气、复脉固脱、补脾益肺、生津养血、安神益智等功效[3]。现代研究表明,人参所含的主要活性成分为人参皂苷,目前已知的人参皂苷有50多种,其中人参皂苷Rb1、Rb2、Rb3、Rc、Rd、Re、Rf、Rg1是主要人参皂苷,占总人参皂苷的90%以上[4,5]。此外,人参还含有多糖、氨基酸、生物碱、黄酮等多种化学成分[6],在抗肿瘤、抗衰老、促进生长发育、提高免疫力等方面具有很好的作用[7,8]。人参不仅作为药材使用,还在药膳、保健、化妆品等多个领域都有广泛的应用[9,10]。人参栽培年限长(一般4~6年),而且生长过程中易受到许多环境因素的限制,如土壤、气候和害虫等[11],对人参的品质具有较大影响。虽然野山参的质量及其人参皂苷含量都很高,但是资源缺乏[12]。
组织培养技术不受时间和地域的制约,在短时间可获得比长期栽培植物更有价值的培养物。在人参的组织培养中,可以通过改变培养基的成分、变化培养条件、添加植物生长调节剂、添加诱导子等有效手段来促进目的产物(主要为人参皂苷)的大量产生,使人参皂苷的生产可以达到预期的结果。Lee等[13]和Murthy等[12]比较了栽培人参根、山参根和不同人参培养物中人参皂苷的积累,发现不定根培养物中(系AR-4)人参皂苷含量积累最多(32.46 mg/g),约是栽培人参根的3倍(11.35 mg/g)。Murthy等[14,15]研究发现,除皂苷Re外,人参不定根中皂苷含量均高于野生参,这些结果均表明了人参组织培养的优越性。本研究综述了人参愈伤组织诱导研究进展,并总结了人参次生代谢产物积累的影响因素,以期为组织培养提高人参的质量和人参皂苷的产量提供参考方法和理论依据。
1 人参愈伤组织诱导研究
通过愈伤组织建立的组织培养技术不仅是植物快速繁殖的手段,也是植物遗传改良、品种选育和提高目的产物的理想途径。1964年,中国植物学家罗士韦教授采用人参根为外植体,成功培养获得愈伤组织[16]。之后,美国、苏联、韩国等国家的研究者也相继获得了人参愈伤组织[17]。
1.1 不同外植体
人参不同的器官和组织对离体培养的反应是不同的,其形态发生的能力也不同。可用于诱导人参愈伤组织的外植体以根和茎最为常见。金英善等[18]表明用东北刺人参的茎段和叶柄在AS和1/2 MS培 养基上,添加2.0 mg/L 6-BA和0.2~1.0 mg/L NAA可以获得数量较多、质量较好的愈伤组织。此后,王建华等[19,20]以人参芽孢、实生苗的根、茎、叶为外植体进行愈伤组织诱导试验,结果表明各种不同外植体在相同诱导条件下均能诱导出愈伤组织,并确定了人参愈伤组织诱导和生长最优培养基分别为MS+2,4-D 3 mg/L+BA 0.5 mg/L+NAA 0.5 mg/L+KT 0.5 mg/L和MS+2,4-D 3 mg/L+KT 1 mg/L。邸雪峰等[21]研究表明,人参茎外植体诱导愈伤组织的最佳培养基为MS+2,4-D 2.57 mg/L,诱导率可达64.75%。黄景嘉等[22]以人参根切片作为外植体,在改良MS培养基中成功诱导出愈伤组织。
此外,有研究者用人参叶片诱导愈伤组织,最佳培养基为MS+7 mg/L 2,4-D和0.9 mg/L KT,诱导率可达80%以上[23]。齐海军等[24]研究也表明,人参叶片能诱导获得愈伤组织,最佳的诱导培养基为MS+2,4-D 2 mg/L+NAA 0.66 mg/L+KT 1 mg/L,在此培养基中接种的叶片外植体,30 d后愈伤组织诱导率为93%。而用人参成熟根诱导得到愈伤组织产生许多胚状体,在补充有6-BA和赤霉素的培养基(1/2 MS或B5)中继代培养得到大量的再生植株[25]。此外,Du等[26]以人参花药进行培养诱导获得愈伤组织,发现培养基中添加5 mg/L 2,4-D和1 mg/L KT诱导愈伤效果显著,转移到分化培养基中40 d后,分化成芽、根和最终的幼苗。
1.2 不同培养条件
组织培养的外在条件对人参愈伤组织的影响以温度和光照最为显著,常在25℃、黑暗环境中诱导人参愈伤组织。在植物生长调节剂配比方面,以2,4-D为主,并添加6-BA、KT、NAA等作为辅助调节剂。有研究者发现,人参根在24℃培养箱中暗培养,经20~30 d可形成愈伤组织,到45 d时,可见明显的淡黄色愈伤组织[22]。蔡茁等[27]发现,广谱植物培养基中N、P、K的配比不能完全适应人参愈伤组织的诱导。基于此,杨晶等[28]研究了人参与胡萝卜共培养体系,发现所获得的愈伤组织在23℃、1 000 lx的条件下生长良好,在培养基MS+2,4-D 3 mg/L+IBA 2 mg/L+KT 0.5 mg/L上培养25 d时皂苷含量达到最高,共培养物的皂苷含量比人参愈伤组织皂苷含量高6.25%。王建华等[29]研究表明,人参发根愈伤组织诱导的最佳培养基为MS+2,4-D 1 mg/L+6-BA 0.1 mg/L;人参发根愈伤组织生长的最佳培养基为MS+2,4-D 1.5 mg/L+KT 1.5 mg/L。Zhang等[30]以野生人参不定根为外植体,在MS+2,4-D 0.5 mg/L+KT 0.3 mg/L的培养基上诱导得到愈伤组织(诱导率53.3%),愈伤组织在含有0.5 mg/L 2,4-D的MS培养基上可诱导得到胚性愈伤组织和体细胞胚,体细胞胚在含有5 mg/L赤霉素的MS培养基上,85%的体细胞胚萌发;萌发的芽在1/3 SH+0.25 mg/L NAA的培养基上,可发育成具有良好主根的植物。
另外,张丹[31]研究了人参发状根悬浮培养,得出以1/2 MS液体培养基(pH 5.8,3%蔗糖,NO3-/NH4+为2∶1,磷酸盐浓度85 mg/L)为最佳培养基;摇床条件最适组合为接种量2.14 g,装液量256.88 mL,温度22.36℃,转速134.68 r/min。在人参细胞培养中,添加0.01~1.00 mmol/L的草酸可以促进人参细胞的生长[32]。
2 组培条件对人参次生代谢产物的影响
2.1 培养基
培养基是供植物组织生长和维持的惟一养料,蔗糖是植物培养所需的主要碳源和能源,碳消耗与生物量积累以及细胞和器官的代谢状态直接相关,培养基中所含成分对人参次生代谢产物有着直接的影响(表1)。为了改善人参细胞生长和提高皂苷的产量,Furuya等[33]研究改良MS培养基(除去NH4NO3)对5年生人参根愈伤组织悬浮培养的影响作用,在培养基提供0.5%葡萄糖和2%蔗糖时,最有利于愈伤组织的生长。在此培养条件下,与对照组相比,皂苷含量达到最大值(7.33 mg/g),并在2周后添加2%的蔗糖,能够改善人参细胞的生长,愈伤组织干重达到最大值(17 g/L)。黄滔等[34]研究表明,随着蔗糖浓度的升高,人参不定根干重增殖倍数呈先升高后降低的趋势,当蔗糖浓度在40 g/L时,单位体积培养基中的多糖产量(230.45 mg/L)和皂苷产量(91.41 mg/L)均达到最大值,表明40 g/L蔗糖浓度最有利于人参不定根中的多糖和皂苷积累。
无机盐在培养基中可调节渗透压、pH和酶活性等,是植物生长和分化所必需的。Paek等[35]研究了MS培养基中铵与硝酸盐的比率对人参培养物中不定根生长和人参皂苷积累的影响,表明低铵浓度和高硝酸盐浓度有利于根系生长,在铵与硝酸盐比率为7.19∶18.5时,不定根的生物量最大;以0 mmol/L铵与18.5 mmol/L硝酸盐的比例实现最大人参皂苷产率(83.6 mg/L),当铵用作惟一的氮源时,根生物量和人参皂苷含量减少了1/3,所以较低浓度的铵与硝酸盐的比例有利于人参培养。
同样,磷元素是细胞内能量代谢的重要因子,田鹏玮[36]以人参悬浮培养细胞为材料,通过优化培养条件对人参多糖和皂苷的积累进行了研究,结果表明适当提高培养基中的PO43-浓度可以促进多糖和皂苷的积累,最高可分别达(30.69±2.42)mg/g和(28.14±4.34)mg/g;Ca2+也是细胞生长所必需的营养元素和次级信号物质,适当提高培养基中钙离子的浓度可以使每克干细胞中总皂苷含量增加7.92 mg。Hg2+对人参细胞中代谢产物的生长也有很大影响,当Hg2+浓度为1 μmol/L时,多糖含量达到最大值(23.52 mg/g),当Hg2+浓度为100 μmol/L时,皂苷含量达到最大值(14.82 mg/g)。
此外,Jeong等[37]以1.5 MS培养基,辅以10 mg/L IAA和75 g/L蔗糖来培养人参不定根,以期提高人参皂苷的积累,在培养10 d后,培养基补充0.75强度的MS培养基,以满足不定根的营养需求,当培养20 d后再补充1.0强度MS培养基,使生物量增加27.45%(28.66 g/L),人参皂苷含量增加8.25%(4.92 mg/g),表明此方式显著促进了不定根的生长和人参皂苷的合成。并且发现培养开始后20~30 d内NH4+和HPO42-离子完全耗尽,其他营养物质(K+,Cl-和SO42-)和糖的消耗量更大。此试验结果表明,随着培养周期的增加,初始培养基不能再满足人参不定根生长的需求,通过培养基补充策略很好地改善了这一问题,对人参不定根中生物量的积累和人参皂苷的生物合成均具有重要促进作用。
2.2 植物生长调节剂
植物生长调节剂是影响人参生物量增长以及次生代谢产物合成的关键因素之一(表1)。Kim等[38]测试了4种生长调节剂2,4-D、IBA、IAA和NAA对人参的调节作用,对比发现,在含有IBA(24.6 μmol/L)的培养基中获得了更高含量的总人参皂苷(8.09±0.30)mg/g。有研究表明,1~5 mg/L浓度范围内的2,4-D对细胞生长至关重要,而0.1 mg/L 2,4-D有利于人参皂苷的合成[39]。在西洋参细胞悬浮培养中发现,除了单一生长调节剂的作用,不同生长调节剂的组合对细胞生长和人参皂苷积累也有很好的效果[40],如MS+6-BA 1 mg/L+2,4-D 0.5 mg/L+NAA 1 mg/L时细胞中人参多糖含量最高(20.82±0.56)mg/g);MS+6-BA 1.5 mg/L+2,4-D 0.5 mg/L+NAA 2 mg/L时细胞中人参总皂苷的含量最高(17.92±0.71)mg/g)[36]。在大规模培养过程中,NAA和6-BA作为生长调节剂[41],而2,4-D仅用于愈伤组织诱导。

表1 培养基成分对人参次生代谢产物的影响
2.3 不同诱导剂
不同的激发子通过信号分子和靶基因诱导不同类型的人参皂苷,诱导方式和诱导持续时间也影响人参次生代谢产物的含量(表2)。Kim等[42-44]发现茉莉酸甲酯(MJ)的过表达增强了转基因人参根中原人参二醇(PPD)的产生,MJ通过催化上调来自Rh2的酶,促进人参皂苷Rg3在毛状根中积累。用MJ处理人参不定根,随着MJ的加入,人参皂苷的产量以剂量依赖性的方式增加,150 μmol/L的MJ诱导后皂苷高达60 mg/g。于丽莉[45]对人参发状根的皂苷合成进行研究,在培养第22 d添加MJ,当作用到培养周期的第27 d,人参发状根皂苷含量达到最大(5.269 mg/g),MJ的最佳添加浓度为6×10-4μmol/L,同时发现,添加MJ后,刺激了发状根代谢活动的增强,有利于次生代谢产物的合成。此外,Hu等[46]发现与MJ相比,茉莉酸-2-羟乙酯(HEJ)可以刺激三七中的人参皂苷Rb1和Rd葡萄糖基转移酶(UGRdGT)活性升高,并表明HEJ对三七中的人参皂苷生物合成的下游调控不同,HEJ可以比MJ更有效地产生人参皂苷Rb1。
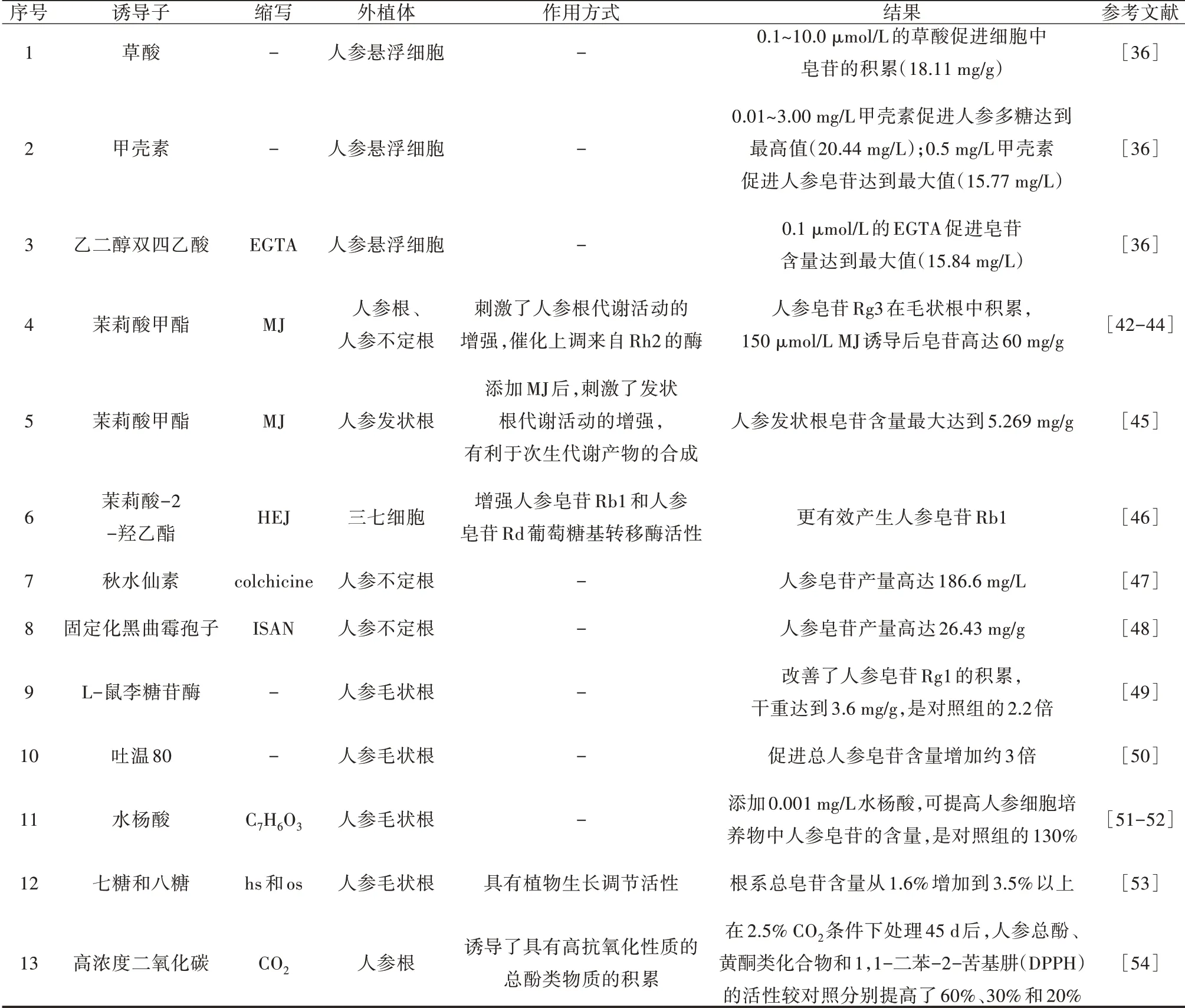
表2 诱导剂对人参次生代谢产物的影响
培养基中添加草酸可以使总皂苷含量提高5 mg/g,加甲壳素使多糖含量增加2.92 mg/g,皂苷含量增加2.32 mg/g;添加Hg2+有助于稀有皂苷C-K的积累,而添加乙二醇双四乙酸(EGTA)增加了皂苷Rh2的积累[36]。Le等[47]用秋水仙素诱导人参不定根产生突变体,在生物反应器中,使用添加了IBA的MS液体培养基培养6周,人参皂苷产量可达186.6 mg/L,比对照组高4.85倍。
Wang等[48]将人参不定根与固定化黑曲霉孢子(ISAN)共培养24 h后,人参皂苷含量最高为(26.43±0.49)mg/g,比未处理对照(8.55±0.78)mg/g高2.09倍。因此,人参不定根与黑曲霉孢子(ISAN)共培养是提高人参皂苷产量的一种很有前景的方法,为促进植物次生代谢产物的积累提供了新的途径。
此外,Zhang等[49]将短双歧杆菌的α-L-鼠李糖苷酶基因过表达到毛根培养体系中,培养30 d后,人参皂苷Rg含量是对照的2.2倍,表明α-L-鼠李糖苷酶基因根提高了人参皂苷Rg1的积累,使干重最高可达3.6 mg/g。与此相同的是,Liang等[50]用吐温80渗透到人参毛状根中处理25 d后,发现总人参皂苷的含量增加了约3倍,表明吐温80不仅可以作为有效的透化剂来促进毛状根分泌人参皂苷,而且还可以作为促进人参皂苷生物合成的诱导剂。
张悦等[51,52]在人参愈伤悬浮培养生长对数期添加0.001 mg/L水杨酸,发现在培养后28 d添加水杨酸可以明显促进人参愈伤组织中人参皂苷的合成,是对照组的130%。Zhou等[53]利用从重楼中分离出的两种低聚糖—七糖(hs)和八糖(os),刺激人参毛状根的生长和人参皂苷的积累,在接种10 d后分别加入30 mg/L的七糖和八糖,根系总皂苷含量从1.6%增加到3.5%以上,结果表明这两种低聚糖在植物组织培养中可能具有植物生长调节活性。另外,Ali等[54]研究发现2.5%的CO2处理人参根45 d后,总酚、类黄酮和1,1-二苯基-2-苦基肼(DPPH)活性分别增加了60%、30%和20%,表明酚类化合物对植物免受二氧化碳的氧化起重要作用。以上结果均表明,向人参根悬浮培养物中递送的高浓度CO2,诱导了具有高抗氧化性质的总酚类物质的积累。因此,人参根是具有高抗氧化能力的酚类化合物的良好来源。
2.4 人参内生菌
内生菌广泛存在于健康植物组织内部,能够产生或促进同原植物相同或相似的活性物质,是植物微生态系统的重要组成部分,人参内生菌对人参皂苷的产生也具有调节作用(表3)。陈雪英[55]从西洋参的根部分离出3株内生细菌,证明了其中1株中含有人参皂苷Rb1,且该菌株可将皂苷Rb1部分转化为人参皂苷Rd。之后,张薇[56]从人参根部得到2株人参内生菌——莫勒接霉(Zygorhynchus moelleri)和灰绿犁头霉(Absidiaglauca),发现能够将人参皂苷Rb1转化为人参皂苷Rd。吕国忠等[57]进一步证实灰绿犁头霉菌具有转化底物人参皂苷Rb1制备Rd的能力。另外,侯耀达等[58]从7年生林下参根部分离得到1株霉菌GS1-33,能有效地将人参根总皂苷转化为人参稀有皂苷C-K及Rh1。此外,陈泠[59]考察3R-2菌丝对人参毛状根生长和次生代谢成分生物合成的影响,研究表明在3R-2菌丝与人参毛状根共培养的14~21 d内有显著的促进作用;培养14d时,人参皂苷Rc的含量达到最高。

表3 人参内生菌对次生代谢产物的影响作用
2.5 其他
目前,除了以上所述的影响条件,在促进人参产生更多次生代谢产物的影响因素方面,还发现几种比较新的方法和诱导条件(表4)。Kim等[60]研究人参-西洋参杂交体(PGQ),发现其生长旺盛且它的根比亲本的大,对PGQ及其亲本的体细胞胚胎发生(SE)进行了比较研究,PGQ外植体的胚胎诱导率较高(2 mg/L 2,4-D作用下胚胎诱导率约56%),其发芽胚在土壤转移后都产生新的芽,在亲本中发现了种间杂交人参,F1杂交体(PGQ)含有2个种特异性人参皂苷(人参皂苷Rf和西洋参假人参皂苷F11),其他人参皂苷含量均高于亲本。

表4 其他作用方式对人参次生代谢产物的影响
张磊等[61]研究NO供体硝普钠(SNP)在光照胁迫下对人参愈伤组织细胞生长及次生代谢产物积累的影响,发现在光照胁迫下,SNP处理后人参愈伤组织中抗氧化酶活性得到增强,清除了强光胁迫下产生的活性氧,缓解了强光对人参愈伤组织的胁迫作用,并发现NO可促进萜类物质的合成,并可促进细胞合成纤维素、木质素、多糖和皂苷的积累。
Le等[62]用伽马射线照射不定根培养物,产生了4个独特的突变株,发现突变系的人参皂苷含量比对照组的高4.2倍。伽马射线处理的不定根突变株代表了人参皂苷商业化生产的良好方式。Lee等[63]用根霉属真菌Rhizopus oligosporus发酵野生人参14 d后,总皂苷含量从3 274 mg/kg增加到5 573 mg/kg。
3 小结与展望
目前,关于人参组织培养技术的研究已经日趋完善,在中国、日本和韩国已经实现了人参组织培养的工业化生产。在人参愈伤组织诱导方面,以人参根、茎、叶诱导最为常见,其中,茎和叶诱导效果比较明显,诱导率比较高,因此,使用人参茎和叶诱导愈伤组织已被广泛采用。人参愈伤诱导大多使用MS培养基,添加2,4-D作为愈伤组织主要诱导调节剂,并添加6-BA、KT、NAA等作为辅助调节剂。
人参皂苷是人参合成的重要代谢产物。在组织培养中,人参的次生代谢产物受培养基、生长素、诱导剂以及人参内生菌等多种因素的影响,研究者们常利用此类因素的影响作用以达到促进人参皂苷积累的目的。因此,深入地研究组培条件对人参次生代谢产物的影响作用,才能最大化地提高人参组培物的质量和产量。随着科技的进步和相关研究的发展,提高人参皂苷的研究必将成为热点,未来将产生巨大的科学价值和经济效益。
猜你喜欢
作物研究(2022年1期)2022-11-27
作物学报(2022年10期)2022-07-21
高师理科学刊(2020年1期)2020-11-26
天然产物研究与开发(2018年4期)2018-05-07
现代园艺(2018年3期)2018-02-10
现代园艺(2017年13期)2018-01-19
山东林业科技(2017年5期)2017-04-05
中国继续医学教育(2015年5期)2016-01-07
中国合理用药探索(2011年3期)2011-03-20
茶业通报(2010年3期)2010-03-21
