
静脉滴注心房利钠肽对阿勒泰羊血液脂代谢激素的影响及尾脂转录组分析
2021-11-24 11:27:24李翔宇刘越越
畜牧兽医学报 2021年11期
马 昕,杨 光,李翔宇,刘越越,陈 勇*
(1.新疆农业大学动物科学学院,乌鲁木齐 830052;2.新疆肉乳用草食动物营养实验室,乌鲁木齐 830052)
阿勒泰羊作为典型的地方脂尾型绵羊,尾脂脂肪过度沉积不仅影响畜产品品质,还明显降低饲料转化率[1],减少尾部脂肪沉积正成为养羊业重要的研究领域之一[2]。
心房利钠肽(atrial natriuretic peptide, ANP)主要由心房心肌作为前体激素产生和分泌,已证实ANP可调节水盐平衡和血压稳定,且表现出强效的脂解作用[3]。ANP促进脂肪细胞褐变、脂肪分解、脂质氧化和脂肪因子分泌,已成为能量消耗和代谢的重要调节因子。尽管ANP曾被认为仅能促进灵长类动物脂肪细胞的脂解[4],且ANP在人和啮齿动物表达中具有同源性[5],但也有研究证实,绵羊也产生ANP[6],本实验室也已发现ANP会影响绵羊脂肪代谢,但其作用机制尚不清楚[7]。本研究利用Illumina高通量测序技术,根据静脉滴注ANP后阿勒泰羊尾脂基因表达和转录组数据,确定ANP对参与绵羊脂解主要通路和关键基因表达规律的影响,为阐明绵羊尾部脂肪沉积的分子调控机制提供基础数据。
1 材料与方法
1.1 试验材料
ANP由苏州强耀生物科技有限公司合成,分子量为3 960.48,纯度为98.83%。ANP、脂联素(adiponectin,ADPN)、胰岛素(insulin,INS)、瘦素(leptin,LEP)和环鸟苷酸(cyclic guanosine monophosphate,cGMP)ELISA检测试剂盒购自上海笃玛生物科技有限公司。
1.2 试验设计及饲养管理
在新疆畜牧科学院绵羊繁育试验基地选取平均体重为(45.6±6.5)kg、健康状况良好的1.5岁阿勒泰母羊8只。采用自身对照试验设计,对照期在晨饲前颈静脉滴注生理盐水,试验期在晨饲前颈静脉滴注ANP溶液。滴注时,绵羊侧卧在人工授精保定台上。对照期时,在(45±2)min内滴注完毕100 mL生理盐水,连续滴注4 d;试验期时,将浓度为562.5 μg·mL-1的ANP母液以1.125 μg·kg-1BW的剂量溶解在100 mL生理盐水中滴注[8],滴注方式与时间同对照期。滴注结束后自由采食、饮水,饲粮组成及营养水平见表1。

表1 饲粮组成及营养水平(以干物质计)
1.3 样品采集与处理
每期的第4天分别在滴注前(0 min)和滴注开始后15、30、45、60、90、120 min采集空腹血液,分离血浆并于-20 ℃保存。各期最后一次血样采集结束后,以盐酸普鲁卡因对尾部局部麻醉后手术采集尾脂0.2 g左右,剪碎后储存于RNAstore保存液中,于4 ℃过夜后-80 ℃冷冻保存备用。
1.4 激素的测定
ANP、脂联素、胰岛素、瘦素和环鸟苷酸ELISA测定按照产品说明进行。
1.5 RNA提取、cDNA文库构建及高通量测序
TRIzol法提取脂肪组织总RNA,并检测纯度及完整性。从对照期和试验期各随机选取5个合格样本送至北京诺禾致源科技股份有限公司,利用Illumina的NEBNext®UltraTM RNA Library Prep Kit试剂盒构建样品的cDNA文库,经检测合格的文库再进行准确定量。随后使用Illumina HiseqTM 2500平台进行测序。
1.6 生物信息学分析
测序下机原始数据经质控后得到有效数据,计算有效数据的容错率以及Q20、Q30、GC的含量,再由HISAT2比对软件比对序列到参考基因组,获得比对数及比对率。基因比对后,通过DESeq2软件对两组测序文库的有效数据进行差异表达分析,以校正后的P(Padj)<0.05且差异表达倍数值|log2fold change|>1为标准,筛选差异表达基因。并对差异表达基因进行GO功能富集和KEGG通路富集分析。
1.7 qRT-PCR验证
根据转录组分析结果,选取7个与脂肪代谢相关的基因,采用荧光定量PCR进行相对定量(2-ΔΔCt),验证高通量测序结果的准确性。在GenBank中搜索绵羊的水通道蛋白7(AQP7)、脂肪酸结合蛋白4(FABP4)、脂滴包被蛋白基因5(PLIN5)、脂联素受体2(ADIPOR2)、单酸甘油酯脂肪酶(MGLL)、胰岛素降解酶(IDE)及长链脂酰辅酶A合成酶1(ACSL1)基因mRNA序列信息,以磷酸甘油醛脱氢酶(GAPDH)为内参基因,使用Primer5.0软件设计引物,引物由生工生物工程(上海)有限公司合成,PCR引物序列见表2。

表2 验证基因及其引物信息
1.8 数据处理
采用SPSS 19.0软件进行配对样本t检验分析,GraphPad Prism 8.0进行做图,所有数据均用“平均值±标准差(mean±SD)”表示,P<0.01表示差异极显著,P<0.05表示差异显著。
2 结 果
2.1 静脉滴注ANP对血浆激素含量的影响
由表3可知,滴注ANP后,血浆中ANP在30、90 min时,试验期显著高于对照期(P<0.05);脂联素在30 min时,试验期显著高于对照期(P<0.05),90 min时,试验期血浆中脂联素含量极显著高于对照期(P<0.01);在30、90 min时,试验期环鸟苷酸含量极显著高于对照期(P<0.01),在45、60 min 时,试验期cGMP含量显著高于对照期(P<0.05);胰岛素在开始滴注后0、30、120 min时,试验期极显著高于对照期(P<0.01),15、45、60、90 min时,试验期胰岛素含量显著高于对照期(P<0.05);瘦素在0、15 min时,试验期显著高于对照期(P<0.05),30 min时,试验期瘦素含量极显著高于对照期(P<0.01)。

表3 滴注ANP对阿勒泰羊血浆脂代谢相关激素含量的影响
2.2 转录组分析
2.2.1 测序数据质检 测序获得的原始数据经质控后平均每个样本的有效数据为5 407万条,数据量为8.11G,样本容错率均在0.03%以下,Q20、Q30分别超过97%和94%,表明获得了较高质量测序数据;质控后的有效数据与参考基因组比对后,总比对率>88%,唯一比对率>79%,表明所选参考基因组可以满足信息分析的需求。
2.2.2 差异表达基因的筛选 以P<0.05且|log2fold change|>1为标准,筛选滴注ANP前后的差异表达基因,发现共有差异基因3 686个,其中1 482个基因上调表达、2 204个基因下调表达,总体基因的表达情况可见火山图(图1)。由差异基因热图(图2)可见,组内两两样品的基因表达量基本趋于一致,而组间样品则存在显著的基因表达变化,证明样品间相关性良好,聚类效果较好。表4所示为对照期与试验期之间脂代谢相关差异基因,PLIN5、PLIN1、MGLL、FABP4、ADIPOR2、AQP1、IDE等与脂肪代谢相关基因的表达量差异极显著(P<0.01),表明滴注ANP对阿勒泰羊尾脂中与脂肪代谢相关基因的表达量产生了一定影响。

表4 滴注ANP对阿勒泰羊尾脂脂肪代谢相关基因表达量(FPKM)的影响

图1 差异基因火山图

图2 差异基因热图
2.3 转录组功能注释
2.3.1 差异基因的GO功能富集分析 对差异基因进行GO分析共富集到118个生物学功能,其中77.1%为生物过程(biology process, BP)、17.0%为细胞组分(cellular component, CC)、5.9%为分子功能(molecular function, MF)。各功能中富集最显著的前10条通路见图3。其中表达上调基因在BP中主要显著富集在电子传递链、核苷酸代谢过程、线粒体复合体I的组装和生物发生及氧化磷酸化相关;在CC中主要显著富集在核糖体组成部分、线粒体、线粒体膜及呼吸链;在MF中主要显著富集在核糖体的构成部分、NADH脱氢酶活性、rRNA结合位点、谷胱甘肽转移酶活性及氧化还原酶活性(图4A)。表达下调基因在BP中主要显著富集在白细胞活化、淋巴细胞活化、免疫反应激活及免疫应答调控;在CC中主要显著富集在浓缩染色体及中心体;在MF中主要显著富集在酶激活剂活性、核苷三磷酸酶调节剂活性、GTP酶激活剂及GTP酶调节剂活性、糖蛋白结合(图4B)。

图3 差异表达基因的GO功能富集

A.表达上调基因;B.表达下调基因
2.3.2 差异基因的KEGG通路富集分析 通过KEGG通路富集分析发现,与对照期相比,表达上调基因共富集到262条信号通路中,其中15条通路显著富集;表达下调基因共富集到284条信号通路中,其中31条通路显著富集。(除去疾病)其中表达上调基因显著富集到核糖体、氧化磷酸化、产热、逆行内源性大麻素信号、细胞色素P450代谢、调节脂肪细胞中的脂肪分解等通路(图5A);表达下调基因显著富集到溶酶体、Toll样受体信号、B细胞受体信号、NOD样受体信号、吞噬体、趋化因子信号等通路(图5B)。其中,与脂代谢和能量相关的主要通路见表5,核糖体、产热、氧化磷酸化通路极显著富集(P<0.01);cAMP信号通路、调节脂肪细胞中的脂肪分解、逆行内源性大麻素信号通路显著富集(P<0.05),表明滴注ANP可能通过调控上述通路实现脂解作用。

表5 滴注ANP对阿勒泰羊尾脂脂肪代谢相关KEGG通路的影响

A.表达上调基因;B.表达下调基因
2.4 差异表达基因的验证
根据功能及代谢通路选取RNA-seq中5个表达上调基因(AQP7、FABP4、PLIN5、ADIPOR2、MGLL)和2个表达下调基因(IDE、ACSL1),经qRT-PCR验证。结果如图6显示,7个功能候选基因表达水平变化趋势与RNA-seq结果基本一致。通过qRT-PCR分析可以看出,IDE、ACSL1在滴注ANP后的表达分别显著(P<0.05)和极显著(P<0.01)下调;ADIPOR2在滴注ANP后的表达显著上调(P<0.05),AQP7、FABP4、MGLL和PLIN5则极显著上调(P<0.01)(图7)。

图6 差异基因qRT-PCR验证
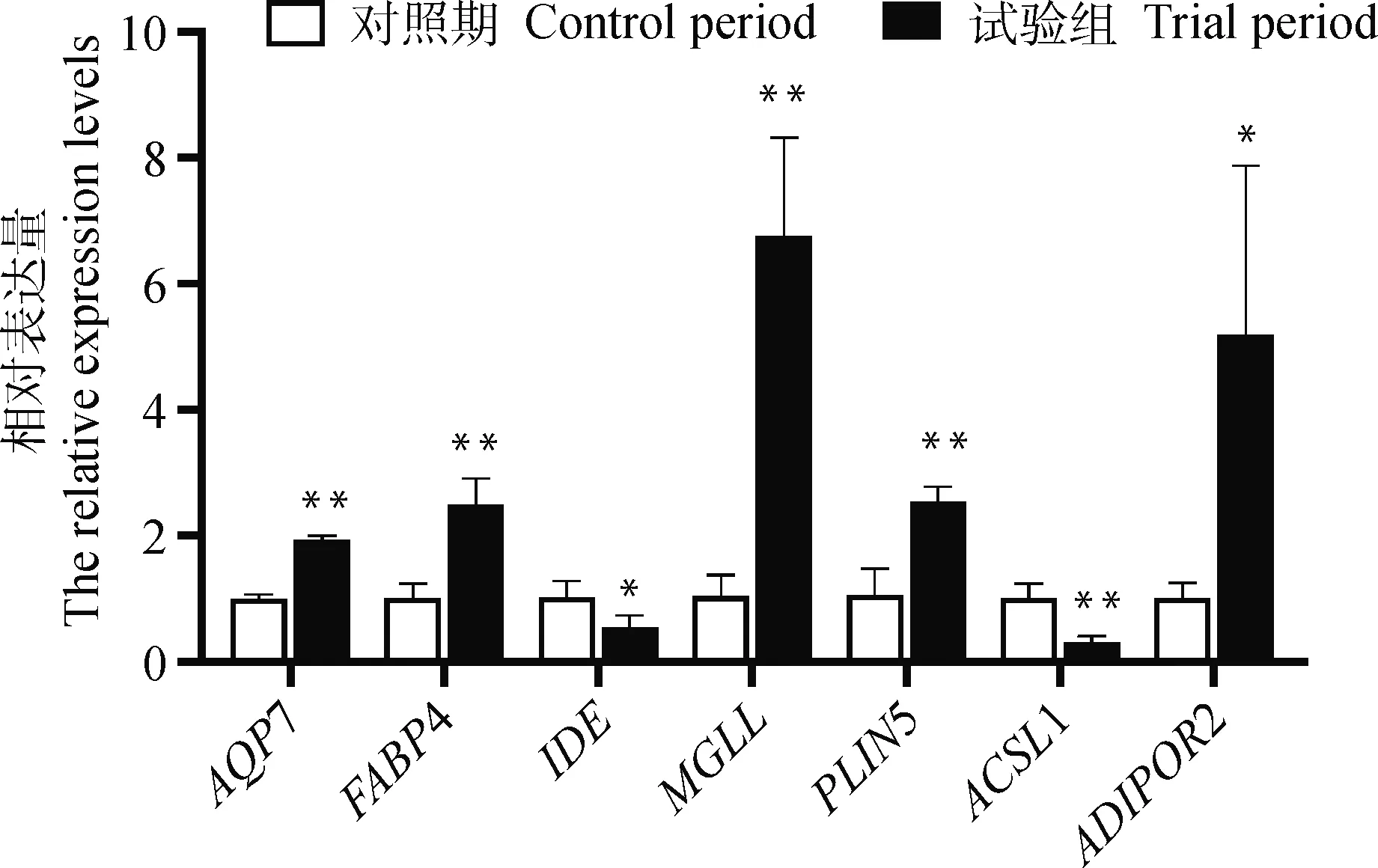
*.差异显著(P<0.05);**.差异极显著(P<0.01)
3 讨 论
在人体皮下组织中注射ANP可诱导脂肪分解并增加局部血流量,从而增强脂质代谢[9]。本试验中,试验期血浆ANP含量均高于对照期,在人与猪模型中输注适宜剂量ANP后均发现了类似效应[10-11],表明滴注外源ANP对血浆ANP含量产生了一定影响。胰腺α和β细胞上存在ANP受体NPRA(natriuretic peptide receptor A),滴注ANP后可能刺激生成更多受体[12],Birkenfeld等[8]的研究结果表明,受试者胰岛素浓度随着ANP注射的增加而增加,本研究中试验全期胰岛素含量均显著增加(P<0.05)。ANP和脑钠肽(brain natriuretic peptide,BNP)通过激活过氧化物酶体增殖物激活受体γ(peroxisome proliferator activated receptor gamma,PPARγ)的表达来调节体内脂肪分布,促进脂肪细胞脂联素的分泌[13]。研究表明,BNP可能通过释放脂肪因子发挥部分代谢作用,BNP与ADPN水平呈正相关[14],由此推测,BNP的代谢效应可能部分是由脂联素介导的。本试验中,试验全期血浆中ADPN平均水平显著高于对照期(P<0.05),Wang等[15]研究也表明,ANP显著增加了总脂联素含量,ANP与BNP有类似生物活性,表明ANP也可能是脂联素分泌的主要刺激因素。NPRA和NPRB与鸟苷酸环化酶受体偶联,可促进GTP转化为二级信使cGMP,从而提高尿液和血浆中cGMP的水平,升高的细胞内cGMP进而激活下游效应因子蛋白激酶G(PKG)促进激素敏感脂肪酶和脂滴包被蛋白的磷酸化[16],本试验中,就全期平均水平来看,试验期血浆中cGMP含量极显著高于对照期(P<0.01),其他学者研究也有类似结果[17],表明滴注ANP后,cGMP含量升高刺激脂肪组织脂解。研究表明,瘦素的分泌与脂肪组织中储存的能量成正比,可通过增强脂肪细胞的生物活性,促进瘦素的释放[18]。本试验中,试验期血浆瘦素含量显著高于对照期(P<0.05),ANP刺激脂肪细胞释放瘦素,高瘦素水平增加脂肪酸氧化,同时减少葡萄糖摄取,导致心脏胰岛素抵抗和从葡萄糖向游离脂肪酸代谢的转变[19]。但瘦素与血浆BNP之间的联系还受其他神经激素共同调控[20],同理推及ANP与瘦素之间互作效用还有待探讨。较高的胰岛素水平和较高的葡萄糖浓度可能通过诱导人脂肪细胞中清除受体NPRC(natriuretic peptide receptor C)的表达,导致ANP介导的脂解减少,从而导致胰岛素抵抗[21],但增加脂联素的分泌可间接改善ANP的胰岛素增敏效果[22]。由此可见,血浆ANP、脂联素、cGMP、胰岛素和瘦素的共同调控可能是联系ANP和糖脂代谢活动之间的关键效用。
研究人员基于RNA-seq技术比较分析同种基因在尾型不同绵羊脂肪组织的相对表达量,以期鉴定遗传决定因素[23]。为揭示差异表达基因的潜在生理功能,本研究对筛选到的差异表达基因进行GO功能与KEGG通路富集分析。GO功能主要富集在核糖体与线粒体功能,脂代谢受蛋白与能量共同调控,ANP可能间接调控关键脂肪细胞分化,实现转录因子(PPARγ、CCAAT增强子结合蛋白(C/EBPα)、固醇调节元件结合蛋白1(SREBP-1)等)功能,促使能量代谢相关过程的上调表达基因显著富集。表达下调的基因主要富集在免疫功能中,表明ANP可能通过某些下调基因参与免疫调节代谢过程。通过KEGG通路富集分析显示,氧化磷酸化、产热通路与核糖体及能量产生密切相关,与本研究GO功能富集结果相对应;Carper等[24]发现,ANP是哺乳动物非颤抖产热的生理性内分泌激活剂,生热脂肪细胞可通过部分线粒体解偶联蛋白1(uncoupling protein 1,UCP1)将化学能散失为热量,以p38MAPK依赖的方式增加棕色脂肪和线粒体标志物的表达。脂肪代谢相关KEGG通路中,cAMP调节胰岛素分泌,瘦素直接刺激醛固酮的分泌[25]。表达下调显著富集的KEGG通路中多数通路与免疫反应通路相关,与本研究GO功能富集分析下调表达结果相对应。
在前期研究滴注ANP后,阿勒泰羊血液中甘油、甘油三酯、游离脂肪酸含量均有不同程度的增加,促进了脂解[7]。Guo等[26]的研究表明,可通过PPAR信号通路依赖方式增强AQP7在脂肪组织中的表达,减轻体重,介导脂肪细胞甘油输出,改善糖脂代谢。FABP4的表达受PPARγ和C/EBPα在转录水平上的调控[27],FABP4通过下调PPARγ信号通路显著抑制了脂肪细胞内脂滴的积累和成脂细胞的形成[28]。本研究中,FABP4的上调表达可能与ANP介导脂肪组织(腹脂和内脏脂肪)向尾脂转运更多脂肪酸有关。PLIN5在脂肪酸氧化程度较高的组织中含量丰富,在cAMP/PKA介导的脂解刺激下,优先与单不饱和脂肪酸结合[29],Wang等[30]研究发现,骨骼肌中PLIN5过表达促进了PPARγ和PPARγ辅活化子-1α(PGC-1α)共同控制下脂肪酸分解代谢。有研究表明,ANP通过激活AMPKα2亚基活性刺激脂解作用[31],ADIPOR2参与AMPK信号通路,介导AMPKα2亚基活性和PPAR活性的升高,ADIPOR2还可介导脂联素激活,刺激PPARα配体表达,导致胰岛素敏感性增加,从而调节糖脂代谢[32]。刺激MGLL被抑制后,体内主要的内源性大麻素-2-花生四烯酰甘油酯分泌水平随之增加[33],与本研究KEGG中逆行内源性大麻素信号通路上调结果一致。IDE最初被认为具有降解胰岛素的能力,后来发现可与ANP相互作用,已经有研究表明,IDE可作为某些疾病的治疗靶点,通过抑制IDE来增强胰岛素的活性[34]。ACSL1的高表达通过PPARβ途径减少脂肪酸γ氧化,参与甘油三酯生物合成及脂肪酸代谢通路从而提高甘油三酯水平,ACSL1下调后甘油三酯水平下降[35],已证实静脉滴注ANP 45 min后血浆中甘油三酯水平有降低趋势[7]。可能由于本研究没有直接评估蛋白质表达水平,也没有直接评估脂肪酸通量,相对的基因表达量并不一定意味着蛋白质将被翻译或具有功能。因此,后续的研究应该同时测量蛋白质水平和脂代谢基因的活性。
4 结 论
4.1静脉滴注ANP后,阿勒泰羊血浆中ANP、脂联素、cGMP、胰岛素和瘦素含量均有不同程度的增加,ANP在一定程度上促进了阿勒泰羊的脂肪动员,增加了脂肪分解。
4.2静脉滴注ANP后尾脂组织中有1 482个基因上调,2 204个基因下调;GO富集分析显示,差异基因主要富集在氧化磷酸化相关的生物过程;KEGG富集分析显示,差异基因主要富集在核糖体、产热、氧化磷酸化和调节脂肪细胞中的脂肪分解等信号通路上。表明外源ANP通过增加氧化磷酸化和产热促进脂肪分解。
猜你喜欢
动物医学进展(2024年4期)2024-04-10 01:50:04
草食家畜(2021年3期)2021-06-08 03:18:16
心电与循环(2020年1期)2020-02-27 07:48:24
中国猪业(2019年7期)2019-11-16 07:15:28
胃肠病学(2016年7期)2016-03-13 23:11:56
安徽医科大学学报(2015年9期)2015-12-16 11:09:42
明清小说研究(2015年3期)2015-11-30 01:26:03
郑州大学学报(医学版)(2015年2期)2015-02-27 14:50:45
郑州大学学报(医学版)(2015年2期)2015-02-27 14:50:45
湖北农业科学(2014年3期)2014-07-21 10:25:00
