
基于表型性状和SSR标记的9份万寿菊种质遗传多样性分析
2021-11-24 03:14宋江琴唐道城丁圆圆马洪强
种子 2021年10期
宋江琴, 唐 楠, 唐道城, 丁圆圆, 马洪强
(青海大学高原花卉研究中心/青海省园林植物与观赏园艺重点实验室, 西宁 810016)
万寿菊(TageteserectaL.)为菊科万寿菊属的一年生植物,原产于墨西哥,因花期长、花型丰富、花色艳丽、易于栽培以及抗病虫害能力较强等优点,在园林景观布置和色素提取上应用广泛[1];另外,因其净化空气和美化环境的功能,市场上对万寿菊的需求也逐渐增大。万寿菊生产上多采用两系配套法制种,即强优势恢复系与雄性不育两用系中的不育系进行杂交获得F1代种子[2-3]。因此,筛选出具有育种潜力的万寿菊雄性不育两用系,对利用雄性不育两用系进行种质创新具有重要意义。
目前国内对万寿菊种质资源的遗传多样性研究报道较少,宋江琴等[4]利用SSR分子标记分析了24份万寿菊雄性不育材料的遗传多样性,结果表明,供试材料具有一定的多样性,并发现部分标记与育性和株高有一定的联系。杨洁等[5]利用SSR分子标记建立了部分万寿菊品种(系)的指纹图谱。杨帆[6]对万寿菊SSR-PCR体系进行了优化。利用形态学标记对万寿菊进行遗传多样性评价较少,其中伍亚平[7]从形态学标记、生化标记、分子标记三个方面对9个万寿菊两用系进行遗传差异性分析,研究其遗传关系。
本研究结合形态学标记和SSR分子标记从万寿菊表型和基因型两方面分析材料的遗传关系,计算材料之间的变异系数和多样性指数,进行性状间主成分分析和材料间聚类分析,并构建了供试材料的数字指纹图谱,为万寿菊亲本选育、种质资源鉴定等相关研究提供理论依据。
1 材料与方法
1.1 试验材料
本研究以青海大学高原花卉中心选育的9个万寿菊雄性不育两用系(C×40-1(2)、34-2(2)、5-2、C×40-1(1)、35-1、34-1、C×40-1(3)、2-2、34-2(1))可育株为材料(表1)。
1.2 9个万寿菊表型性状的测量
根据《中华人民共和国万寿菊DUS测试指南》中品种性状记载标准,对8个数量性状(株高、冠幅、花朵数、一级分枝数、花径、开花前期、单花花期、千粒重、花色)采用测量法统计,对1个质量性状(花色)采用目测法记录(表1),将花色分成三个色号,即1#(偏浅黄色),2#(偏橙色),3#(偏橘红色)。每个品系至少测量30株。
1.3 SSR分子标记
于2018年7月从试验田采集材料盛花期嫩叶为样本,每株取3~5片叶,液氮冷冻,-80 ℃保存。采用改良CTAB法[8]进行DNA提取,用琼脂糖凝胶电泳(2%)和微量核酸仪检测DNA质量和浓度,DNA提取液于-20 ℃保存备用。SSR引物序列来源于青海大学高原花卉中心万寿菊2-2转录组信息(表2)[4]。从78对合成的引物中筛选出11对多态性良好的引物进行扩增分析,琼脂糖凝胶电泳(2%)检测是否具有目标条带,利用8%聚丙烯凝胶电泳电压220 V分离约1.2 h,使用硝酸银染色法进行染色,显影后记录基因型数据。
1.4 统计分析
计算表型性状的均值、标准差、变异系数和 Shannon-Weaver 多样性指数。Shannon-Weaver多样性指数的计算参考孙东雷等[9]的方法。利用SPSS 19.0软件进行性状间的主成分分析和显著性分析[10],材料间的聚类分析,采用Ward 法和欧氏距离进行系统聚类。
统计目标位置条带的有无,赋以“1”或“0”,缺失数据记为“-9”,建立“0,1”分子数据矩阵。通过Cervus 3.0软件[11]计算引物的PIC(多态性信息)值。利用Popgen(Version 1.32)软件[12]计算各引物的观察等位基因数(Na),有效等位基因数(Ne),Nei’s多样性指数和Shannon’s信息指数(I)等遗传参数;利用 NTSYS(version 2.10 t)软件计算9个两用系品系间的遗传相似系数并进行非加权组平均法(UPGMA)聚类分析。
2 结果与分析
2.1 形态指标的观测与分析
对9个万寿菊雄性不育两用系的9个表型性状进行记录与分析(表1)。结果表明,9个两用系株高范围66.31~77.8 cm,变异系数7.2%;冠幅范围42.63~69.53 cm,变异系数12.6%;花朵数范围17.33~36.8朵,变异系数最大(19.7%);一级分支数范围12.27~15.63,变异系数10%;花径范围6.33~7.71 cm,变异系数2.9%;开花前期范围61~66 d,变异系数最小(2.8%);单花花期范围24~27 d,变异系数3.7%;千粒重范围3.15~4.53 g,变异系数16.8%。结合变异系数,Shannon-Wiener信息指数和方差分析结果可以看出,除单花花期外,株高、冠幅、花朵数、一级分支数、花径、千粒重和开花前期在品系间差异均达到极显著水平(p<0.01)。一级分枝数的遗传多样性指数最大,表明在9个两用系中一级分枝数性状变异丰富。其次是千粒重、冠幅、开花前期、花径、株高、花朵数。单花花期的遗传多样性指数最小,表明9个两用系的单花花期时间较为一致。

表1 9个万寿菊雄性不育两用系性状统计分析
2.2 基于形态指标对不同品系的主成分分析
以8个数量性状为基础,一般特征值大于 1 作为提取成分的标准[19],提取的3个主成分的特征值均大于1(表2),可以反映原始变量的主要信息。3个主成分的方差贡献率分别为42.588%,23.028%,17.731%,累计贡献率达到83.348%。以选取主成分荷载值绝对值大于0.5的性状为标准[14-15],第一主成分主要包括株高、冠幅、花朵数、一级分枝数、花径等性状,主要反映植株的长势、花朵等信息。第二主成分主要包括千 粒重等性状,主要反映了植株种子的情况。第三主成分主要包括花径、单花花期等性状,主要反映单花开放时间及植株营养生长时间等信息。株高、冠幅、一级分枝数、花朵数基本上可以作为反映这9个万寿菊两用系的表型性状,简化了描述指标,可视为今后这9个万寿菊两用系种质资源评价和亲本选择的主要考量指标。

表2 不同品系形态指标的主成分分析
2.3 基于形态指标对不同品系的聚类分析
以8个数量性状为基础,对9个万寿菊雄性不育两用系进行聚类(图1),结果表明,在阈值为5时,9个两用系被聚成三个大组,第一大组又包括2个小组,其中第一小组包括34-2(2),5-2和34-2(1),花朵数量集中在35~37朵之间,株高均在77 cm左右;第二小组包括C×40-1(2),C×40-1(1)和C×40-1(3),为同一系列不同花色的3个品系;第二大组包括35-1和34-1,花色均为浅黄色。材料2-2被单独聚成一类,其冠幅和花朵数均极显著低于其他品系,表明2-2与其他组的材料之间有较大的形态差异。
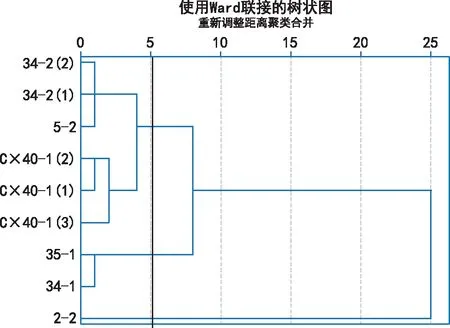
图1 基于8个数量性状的聚类分析
2.4 基于SSR标记的引物多态性分析
用筛选出的11对多态性良好,条带清晰,易于读带的SSR引物在9个两用系中的扩增信息如表3,对9个两用系品系的扩增结果如图2。11对引物共检测到29个条带,其中27个具有多态性,多态性比率为93.1%,平均每个位点等位基因数为2.45个。观测等位基因数(Na)为2;有效等位基因数(Ne)为1.78;Nei’s多样性指数(H)为0.429 1;Shannon信息指数(I)为0.607 8。本研究筛选的11对SSR引物的PIC值为0.353,具有中度多态性,说明11对引物可用于9个两用系材料的遗传多样性分析[16]。

注:材料编号1~9分别代表的材料名称为C×40-1(2)、34-2(2)、5-2、C×40-1(1)、35-1、34-1(2)、C×40-1(3)、2-2、34-2(1)。A→K:分别代表引物U 0860,U 1829,U 1456,U 0878,U 1795,U 1757,U 2041,U 2261,U 106-1,U 203,U 358对9个材料的PCR扩增结果。
2.5 基于SSR标记的9个万寿菊雄性不育两用系遗传多样性分析
遗传距离(表4)表明,材料34-2(2)和35-1材料的遗传距离最大(0.968 3),表明其亲缘关系最远;C×40-1(2)和2-2的遗传距离最小,为0.072,表明这两者亲缘关系最近。基于SSR标记的聚类结果(图3),当阈值为0.60时,9个雄性不育两用系被分成三大组,第一大组包括C×40-1(2),5-2,C×40-1(3),2-2和34-2(2);第二大组包括C×40-1(1);第三大组包括35-1,34-1和34-2(1)。综合形态聚类结果可以发现,最终35-1,34-1在表型聚类和分子聚类的同一类中,被聚为一类;C×40-1(3),C×40-1(2),5-2和34-2(2)在表型聚类和分子聚类的同一类中,被聚成一类;34-2(1),2-2 和C×40-1(1)在表型和分子的聚类结果不一致。
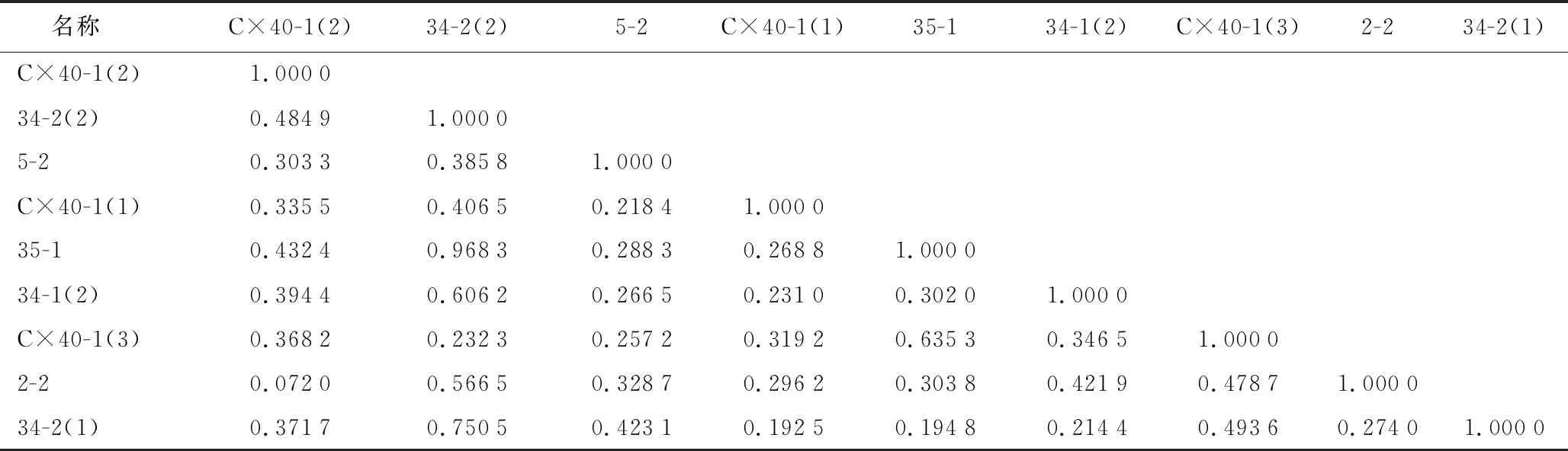
表4 9个万寿菊雄性不育两用系遗传距离
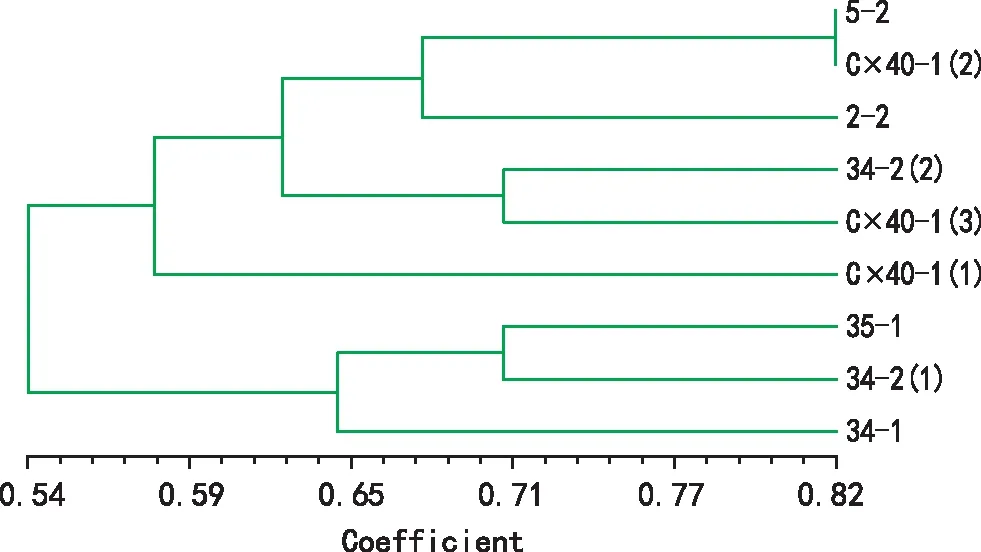
图3 9份万寿菊种质资源基于SSR标记的UPGMA聚类
2.6 指纹图谱构建
利用筛选出的11对SSR引物对9个万寿菊品系进行数字指纹图谱的构建(表5)。由SSR-PCR扩增图谱发现,引物U 1757、U 2041和U 1829可以将供试的9个万寿菊品系全部区分开来。对筛选出来的3个引物的扩增结果条带分别进行“0”“1”读带,构建3个万寿菊的数字指纹图谱。使用2对引物U 1757和U 2041可以区分7个(78%)品系,再结合引物U 1829可以完全区分9个品系。
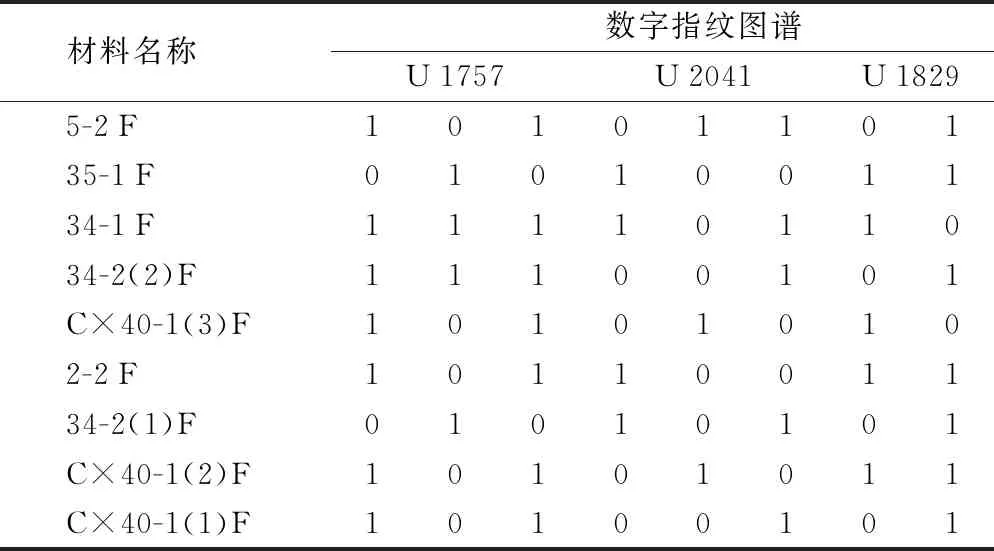
表5 9个万寿菊雄性不育两用系品系数字指纹图谱
3 结论与讨论
万寿菊种质资源创新通常是通过从国外引进新品种来实现,选育一些适合本土生长的万寿菊新品种具有市场性和创新性,故了解现有万寿菊种质资源的遗传背景及差异性有其必要性。本研究对9个万寿菊雄性不育两用系的9个形态学标记和11个SSR分子标记的多样性分析结果表明,供试材料具有丰富的遗传多样性。万寿菊的选育通常是根据形态的综合指标来进行,然而表型性状易受环境因子的影响,不能准确地对万寿菊品系进行鉴定。SSR分子标记具有较高的灵敏度,在一定程度能从DNA水平上揭示材料的遗传变异,故通过形态学差异结合分子标记技术来分析材料的遗传多样性,增加了结果的可靠性与科学性。
供试材料的表型遗传多样性指数为1.49,略低于多头菊种质资源的数量性状的遗传多样性指数(1.856),数量性状中花朵数的变异系数最大,为19.7%,但明显低于多头菊的花朵数[17],说明供试万寿菊的花朵数较多头菊来说该性状较为稳定。各性状除单花花期外,在品系间的差异均达到了极显著水平,说明这9个万寿菊种质资源之间的差异较大,作为母本选育的潜能也较高。主成分分析可以筛选出具有代表意义的性状,从8个表型性状中提取了3个主成分因子,累计贡献率达到83.348%,微低于非洲菊的7个主成分累计贡献率86.567%[21]。3个主成分因子中,荷载值较大的性状代表了主要的表型性状变异,其中影响这9个万寿菊种质资源表型差异较大的性状主要有株高、冠幅、一级分枝数和花朵数。
本研究利用11对SSR引物扩增出29个条带,其中27个条带具有多态性,远高于张华丽等[18]筛选出来的引物多态性比率(43.3%)。从引物扩增条带数来看,李浦[19]筛选出来单对引物扩增条带数(2.1)与本研究(2.45)基本一致。材料34-2(2)和35-1的遗传距离最大,亲缘关系最远;C×40-1(2)和2-2的遗传距离最小,亲缘关系最近。分子标记遗传多样性结果表明,9个两用系的Shannon信息指数为0.607 8,遗传相似系数的变化范围为0.54~0.82,平均值为0.679,说明这9个万寿菊雄性不育两用系具有较为丰富的遗传多样性,这一结果和表型遗传多样性分析结果基本一致[22]。虽然表型聚类和分子聚类结果没有达到完全吻合,但材料34-1和35-1;C×40-1(3)、C×40-1(2)、5-2和34-2(2)无论是在表型层面还是分子层面均划为一类。部分材料聚类结果没有达到一致,其原因可能与表型性状易受环境影响,测量时有一定的误差有关;也有可能与SSR位点的数目太少,不足以覆盖万寿菊整个基因组,以及供试样本的数量有关,这些因素都会影响两种聚类方法的结果,但两种聚类结果都有一定程度的参考作用[20],为万寿菊亲本的选育奠定一定的理论基础。
本研究利用3个SSR引物将9个万寿菊雄性不育两用系完全分开,构建了数字指纹图谱,为不同万寿菊品系的区分、杂交亲本的选择提供了依据。
猜你喜欢
水上消防(2022年1期)2022-11-20
湖南农业科学(2022年1期)2022-06-06
广西蚕业(2021年3期)2021-10-28
军民两用技术与产品(2021年7期)2021-10-13
云南农业(2021年9期)2021-09-24
军民两用技术与产品(2021年12期)2021-03-09
参花(下)(2019年8期)2019-09-11
云南农业科技(2019年2期)2019-04-04
食品与生活(2018年1期)2018-02-07
安徽农学通报(2017年24期)2018-01-12
